
Abstract
Background
Macrophages play significant roles in innate immune responses and are heterogeneous cells that can be polarized into M1 or M2 phenotypes. PRMT2 is one of the type I protein arginine methyltransferases involved in inflammation. However, the role of PRMT2 in M1/M2 macrophage polarization remains unclear. Our study revealed the effect and mechanism of PRMT2 in macrophage polarization.
Methods
Bone marrow-derived macrophages (BMDMs) were polarized to M1 or M2 state by LPS plus murine recombinant interferon-γ (IFN-γ) or interleukin-4 (IL-4). Quantitative polymerase chain reaction (qPCR), western blot and flow cytometry (FCM) assay were performed and analyzed markers and signaling pathways of macrophage polarization.
Results
We found that PRMT2 was obviously upregulated in LPS/IFN-γ-induced M1 macrophages, but it was little changed in IL-4-induced M2 macrophages. Furthermore, PRMT2 konckdown increased the expression of M1 macrophages markers through activation of STAT1 and decreased the expression of M2 macrophages markers through inhibition of STAT6.
Conclusions
PRMT2 silencing modulates macrophage polarization by activating STAT1 to promote M1 and inhibiting STAT6 to attenuate the M2 state.
Similar content being viewed by others


Background
Macrophages are essential in tissue homeostasis and inflammation, which perform vital tissue-specific functions and protect the organism from infection [1]. Additionally, macrophage populations vary among tissues, with osteoclasts and microglia as noted examples [2]. Notably, the diversity and plasticity of macrophage lineages are crucial aspects of their function. Murine bone marrow-derived macrophages (BMDMs) are usually used to research macrophage functions in vitro, which can be generated from bone marrow to mature macrophages by the addition of recombinant murine macrophage colony-stimulating factor (M-CSF) [3]. Macrophages can also be induced into different phenotypes when undergoing classical or alternative activation, called M1 (induced by LPS/IFN-γ) or M2 macrophages (induced by IL-4/IL-13) in vitro, respectively. Evidence has shown that M1 phenotype is tightly associated with pro-inflammatory responses, yet M2 state plays a critical role in anti-inflammatory responses [4, 5]. M1-type pro-inflammatory markers include tumor necrosis factor α (TNF-α), IL-1β, IL-6, inducible nitric oxide synthase (iNOS), and CD86, whereas IL-10, IL-13, found in inflammatory zone 1 (FIZZ1), Arginase 1 (Arg1), transforming growth factor β (TGF-β) and CD206 are M2-type anti-inflammatory markers [6, 7].
Post-translational modifications (PTMs) refer to chemical modifications during the post-translational formation of proteins. Specifically, during the process of protein synthesis, certain biochemically functional groups (phosphate groups, acetyl groups, lipids, carbohydrates, etc.) can be covalently bound to the protein backbone or side chains by the action of modifying enzymes that change their biophysical properties, thus affecting their localization, stability, interactions and lying at the heart of the fields of epigenetics and signal transduction [8]. Protein arginine methylation is a type of PTM catalyzed by protein arginine methyltransferases (PRMTs). Additionally, they can modify target proteins through methylation of the guanidinium nitrogen atom to the arginine residue [9]. Notably, PRMTs modulate basic cellular processes, including transcription, RNA processing, signal transduction cascades, and DNA damage responses [10,11,12]. PRMTs include PRMT1 to 9 and can produce MMA (mono-methyl-arginine), aDMA (asymmetric dimethyl-arginine), or sDMA (symmetric dimethyl-arginine) [13]. Based on the final methylarginine product generated, PRMTs are classified into three categories: type II PRMTs, including PRMT5 and PRMT9 catalyzing the generation of MMA and sDMA; type III PRMT, including PRMT7 catalyzing the generation of MMA, and the remaining are type I PRMTs catalyzing the generation of MMA and aDMA [14, 15].
PRMT2 is unique in the PRMT family for containing an Src Homology 3 (SH3) domain. It has been illustrated that it acts as a transcriptional coactivator for nuclear hormone receptors and is consequently associated with breast cancer [16]. A previous study indicated that PRMT2 was associated with the development of glioblastoma [17]. Higher PRMT2 expression is observed in intestinal specimens from patients with Crohn’s disease and ulcerative colitis, additionally, PRMT2 represses the SOCS3 promoter via histone H3R8 asymmetric dimethylation, thereby promoting the development of DSS-induced colitis [18]. PRMT2 has also been described to increase the accumulation of IκB-α (NF-κB blocker) in the nucleus, which inhibits NF-κB-dependent transcription [19]. Furthermore, in lung tissue and macrophages, reduced PRMT2 expression could increase their responsiveness to LPS and promotes the expression of inflammatory cytokines [20]. It has also been shown that PRMT2 is involved in the regression of diabetic atherosclerosis, and PRMT2 deficiency fosters the expression of genes associated with cytokine signaling and inflammatory pathways in atherosclerotic plaque CD68+ cells [21]. In addition, LPS induces arginine methylation of TLR4 and IRF3 in RAW264.7 via PRMT2, thereby promoting IFN-β production [22]. Thus, the expression of PRMT2 in macrophages is closely related to inflammation. However, few studies have investigated the relationship between PRMT2 and macrophage polarization and its mechanisms.
In this study, we screened and found that PRMT2 could regulate macrophage polarization. Briefly, PRMT2 silencing promotes M1 polarization by STAT1 activation and attenuates M2 polarization through STAT6 inhibition.
Results
PRMT2 is upregulated in LPS- or LPS/IFN-γ-treated BMDMs
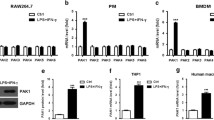
To investigate the potential roles of PRMTs in macrophage polarization, we treated mature BMDMs with LPS or LPS plus IFN-γ to induce the M1 phenotype. As depicted in Fig. 1A, the expression of Prmt2 mRNA was considerably increased by LPS in BMDMs. Consistent with this result, LPS treatment enhanced Prmt2 expression in BMDMs with different doses and times (Fig. 1B, C). We next detected the mRNA level of M1 markers and found that Tnf-α, IL-1β, and Nos2 expression were elevated obviously after LPS/IFN-γ stimulation (Fig. 1D-F). Similarly, surface marker CD86 was also significantly increased in LPS/IFN-γ-stimulated BMDMs by flow cytometry (Fig. 1G). Furthermore, we confirmed that the expression of Prmt2 was enhanced in LPS/IFN-γ-treated BMDMs (Fig. 1H). Therefore, these data suggest that PRMT2 is upregulated by LPS or LPS/IFN-γ in BMDMs, which may act as a regulator of macrophage polarization.

PRMT2 is upregulated in LPS- or LPS/IFN-γ-treated BMDMs. (A) Quantitative polymerase chain reaction (qPCR) analysis of Prmts expression in BMDMs treated with LPS (1 µg/ml) for 6 h. (B) qPCR analysis of Prmt2 expression in BMDMs treated with LPS (1 µg/ml and 5 µg/ml) for 6 h. (C) qPCR analysis of Prmt2 expression in BMDMs treated with LPS (1 µg/ml) for the indicated times. (D-G) qPCR analysis of Tnf-α (D), Il-1β (E), and Nos2 (F) expression and Flow cytometry (FCM) analysis of CD86 (G) in BMDMs treated with LPS (100 ng/ml) plus IFN-γ (20 ng/ml) for 24 h. FMO, fluorescence minus one. (H) qPCR analysis of Prmt2 expression in BMDMs treated with LPS (100 ng/ml) plus IFN-γ (20 ng/ml) for 24 h. 18s rRNA was used as an endogenous reference for qPCR. Data are representative of three independent experiments (mean ± SD). ***p < 0.001, ****p < 0.0001
PRMT2 silencing promotes M1 polarization
Since PRMT2 is elevated in M1 macrophages, we next silenced endogenous PRMT2 expression in BMDMs using Prmt2-specific siRNA and confirmed that Prmt2 mRNA and PRMT2 protein expression were notably reduced in PRMT2-knockdown cells (Fig. 2A, B). To explore the effect of PRMT2 during macrophage polarization, we exposed PRMT2-silenced BMDMs to LPS/IFN-γ induces M1 phenotype. Then, M1 macrophage markers, including TNF-α, IL-1β, iNOS, and CD86, were examined. As presented in Fig. 2C-E, Tnf-α, Il-1β, and Nos2 mRNA levels were significantly increased in PRMT2-knockdown M1 macrophages compared to controls. Similarly, CD86 expression was also increased in PRMT2-knockdown M1 macrophages by flow cytometry (Fig. 2F, G). LPS/IFN-γ treatment also enhanced iNOS protein levels in PRMT2-silenced macrophages (Fig. 2H). Thus, these results indicate that PRMT2 silencing promotes M1 polarization.

PRMT2 silencing promotes M1 polarization. (A, B) qPCR analysis of Prmt2 mRNA (A) expression and immunoblot analysis of PRMT2 protein (B) in BMDMs transfected with Prmt2 siRNA for 48 h. (C-H) qPCR analysis of Tnf-α (C), Il-1β (D), and Nos2 (E) mRNA expression and FCM analysis of CD86 (F) expression in BMDMs transfected with Prmt2 siRNA for 48 h and then treated with LPS (100 ng/ml) plus IFN-γ (20 ng/ml) for 24 h. (G) Quantification of mean fluorescence intensity of CD86 in (F). (H) Immunoblot analysis of iNOS in BMDMs transfected with Prmt2 siRNA for 48 h and then treated with LPS (100 ng/ml) plus IFN-γ (20 ng/ml) or IL-4 (20 ng/ml) for 24 h. 18s rRNA was used as an endogenous reference for qPCR. Data are representative of three independent experiments (mean ± SD). **p < 0.01, ***p < 0.001, ****p < 0.0001
PRMT2 silencing attenuates M2 polarization
It is clear that PRMT2 knockdown promotes M1 polarization. To determine whether PRMT2 silencing affects M2 polarization, we treated BMDMs with IL-4 to induce M2 polarization and demonstrated that the transcription of Il-10 and Arg1 was successfully induced in IL-4-treated macrophages (Fig. 3A, B). Consistent with these results, FCM showed that IL-4 treatment notably increased the expression of surface marker CD206 in BMDMs (Fig. 3C). However, unlike M1 macrophages, compared to M0 macrophages, the expression of Prmt2 was not changed in M2 macrophages (Fig. 3D). Regardless, the expression of Il-10 and Arg1 were reduced in PRMT2-knockdown M2 macrophages (Fig. 3E, F). Simultaneously, CD206 expression was reduced by the detection of flow cytometry (Fig. 3G, H). As predicted, IL-4 treatment decreased ARG1 protein level in PRMT2-knockdown macrophages (Fig. 3I). Therefore, silencing of PRMT2 attenuates M2 polarization.

PRMT2 silencing attenuates M2 polarization. (A-C) qPCR analysis of Il-10 (A) and Arg1 (B) expression and FCM analysis of CD206 (C) in BMDMs treated with IL-4 (20 ng/ml) for 24 h. (D) qPCR analysis of Prmt2 expression in BMDMs treated with IL-4 (20 ng/ml) for 24 h. (E-H) qPCR analysis of Il-10 (E) and Arg1(F) expression and FCM analysis of CD206 (G) in BMDMs transfected with Prmt2 siRNA for 48 h and then treated with IL-4 (20 ng/ml) for 24 h. (H) Quantification of mean fluorescence intensity of CD206 in (G). (I) Immunoblot analysis of ARG1 in BMDMs transfected with Prmt2 siRNA for 48 h and then treated with LPS (100 ng/ml) plus IFN-γ (20 ng/ml) or IL-4 (20 ng/ml) for 24 h. 18s rRNA was used as an endogenous reference for qPCR. Data are representative of three independent experiments (mean ± SD). **p < 0.01, ****p < 0.0001
PRMT2 silencing promotes M1 polarization through STAT1 activation
Next, we explored how PRMT2 modulated macrophage polarization. It is well known that signal transducer and activator of transcription 1 (STAT1), MAPK, and NF-κB pathways serve critical roles in inflammation and macrophage polarization [23]. To reveal whether PRMT2 knockdown affects these pathways in macrophages polarized toward the M1 state, we treated BMDMs with LPS/IFN-γ and found that phosphorylated ERK (p-ERK), p-JNK, p-P38, and p-P65 were only slightly influenced by PRMT2 silencing (Fig. 4A). However, STAT1 phosphorylation increased dramatically in PRMT2-knockdown BMDMs following LPS/IFN-γ treatment (Fig. 4A). To further validate the activation of STAT1 affecting M1 macrophage polarization in PRMT2-silenced BMDMs, the STAT1 inhibitor Fludarabine was administrated. Notably, Fludarabine blocked LPS/IFN-γ-induced elevation of Tnf-α, Il-1β, and Nos2 expression in PRMT2-silenced cells (Fig. 4B-D). Likewise, flow cytometry showed that CD86 increase was also blocked by Fludarabine in LPS/IFN-γ-treated PRMT2-knockdown macrophages (Fig. 4E, F). Collectively, PRMT2 silencing promotes M1 polarization through the activation of STAT1 signaling.

PRMT2 silencing promotes M1 polarization through STAT1 activation. (A) Immunoblot analysis of phosphorylated STAT1, ERK, JNK, P38, and P65 or total proteins in lysates of BMDMs transfected with Prmt2 siRNA for 48 h and then treated with LPS (100 ng/ml) plus IFN-γ (20 ng/ml) for the indicated times. (B-E) qPCR analysis of Tnf-α (B), Il-1β (C), and Nos2 (D) expression and FCM analysis of CD86 (E) in BMDMs transfected with Prmt2 siRNA for 48 h and then pretreated with STAT1 inhibitor Fludarabine (10 µM) for 1 h following by LPS (100 ng/ml) plus IFN-γ (20 ng/ml) stimulation for 24 h. (F) Quantification of mean fluorescence intensity of CD86 in (E). 18s rRNA was used as an endogenous reference for qPCR. Data are representative of three independent experiments (mean ± SD). *p < 0.05, ***p < 0.001, ****p < 0.0001
PRMT2 silencing attenuates M2 polarization through STAT6 inhibition
To further determine the mechanism of PRMT2-mediated M2 polarization, we detected phosphorylated STAT3 (p-STAT3) and p-STAT6, which are reported to regulate M2 polarization [24]. As shown in Fig. 5A, IL-4-induced STAT6 phosphorylation was noticeably reduced in PRMT2-knockdown BMDMs. Most importantly, the reduction of Il-10 and Arg1 expression caused by PRMT2 silencing could be counteracted by the STAT6 inhibitor, AS1517499. Specifically, these M2 markers were largely unaffected between controls and PRMT2 silencing with the addition of AS1517499 (Fig. 5B, C). Expectedly, the same conclusion was obtained from the flow cytometry detection of CD206 (Fig. 5D, E). Taken together, PRMT2 silencing attenuates M2 polarization by inhibition of STAT6 signaling.

PRMT2 silencing attenuates M2 polarization through STAT6 inhibition. (A) Immunoblot analysis of phosphorylated STAT3 and STAT6 or total proteins in lysates of BMDMs transfected with Prmt2 siRNA for 48 h and then treated with IL-4 (20 ng/ml) for the indicated times. (B-D) qPCR analysis of Il-10 (B) and Arg1 (C) expression and FCM analysis of CD206 (D) in BMDMs transfected with Prmt2 siRNA for 48 h and then pretreated with STAT6 inhibitor AS1517499 (1 µM) for 1 h following by IL-4 (20 ng/ml) stimulation for 24 h. (E) Quantification of mean fluorescence intensity of CD206 in (D). 18s rRNA was used as an endogenous reference for qPCR. Data are representative of three independent experiments (mean ± SD). **p < 0.01, ***p < 0.001, ****p < 0.0001. ns, not significant
Discussion
In the present study, we provide evidence to demonstrate that PRMT2 modulates macrophage polarization in vitro. PRMT2 belongs to PRMTs and can act as a coactivator of several nuclear hormone receptors [25]. A previous study found that PRMT2 promotes apoptosis by suppressing NF-κB-dependent transcription [19]. Moreover, PRMT2 plays a role in LPS-induced pulmonary inflammation and airway distress syndrome by regulating NF-κB [20]. In a study, utilizing lung tissue and macrophages, the expression of PRMT2 was downregulated with LPS, allowing NF-κB to bind to the promoter of its target gene. The absence of PRMT2 facilitates NF-κB accumulation in the nucleus after LPS treatment and then increases the production of TNF-α and IL-6, resulting in an inflammatory response. However, in our study, we demonstrate that PRMT2 is upregulated in LPS- or LPS/IFN-γ-treated BMDMs, and PRMT2 silencing has little influence on NF-κB activation. Our findings indicate that PRMT2 regulation of macrophage polarization might be an intrinsic mechanism of PRMT2-mediated inflammatory regulation. Our study is also consistent with Beyza Vurusaner and his/her colleagues’ research that loss of PRMT2 in BMDMs shows an increased expression of pro-inflammatory genes toward LPS and compatibility with a decrease in genes of inflammation resolving in response to IL-4 [21].
Signal transducer and activator of transcription (STAT) proteins consist of STAT1, 2, 3, 4, 5a, 5b, and 6, which are transcription factors localized in the cytoplasm [26]. STATs are widely distributed in the cytoplasm at rest and translocate into the nucleus in response to stimulation of extracellular ligands, cytokines, and growth factors, which participate in biological processes such as cell proliferation, differentiation, migration, and apoptosis [27]. Evidence suggests that IFN-γ activates JAK-STAT1 signaling and promotes STAT1 phosphorylation to promote the polarization of M1-type macrophage [28]. Meanwhile, tyrosine phosphorylation and activation of STAT6 mediates M2 polarization via transcriptional activation of M2 phenotype-specific genes [29]. In STAT6-overexpressing macrophages, M2 genes enhanced expression [30], whereas the ablation of STAT6 eliminated M2 gene expression [31]. Additionally, macrophage plasticity can be driven by different stimuli. With IFN-γ stimulation, STAT1 binds many regulatory elements linked to IL-4-inducible genes. Similarly, STAT6 binds a significant portion of the regulatory elements of IFN-γ activated genes in response to IL-4 [32]. Many STAT1- and STAT6-binding events generally occur on overlapping or adjacent genomic regions. The data imply a reciprocal inhibitory role of STAT1 and STAT6 in the genomic organization. Notably, PRMT2 silencing may affect these regions and thus modify STAT1 activation and STAT6 inhibition during macrophage polarization. In addition, the mutual inhibition between STAT1 and STAT6 may be due to the ability of STAT1 to transcribe some regulatory factors to regulate STAT6, and vice versa.
JAK-STAT signaling is regulated by arginine methylation. It includes the mechanism of PRMT1 binding and methylating STAT1 on Arg31 [33]. Arg31 is conserved in both STAT3 and STAT6, which can also undergo arginine methylation [34, 35]. Importantly, PRMT1 catalyzes STAT3 arginine methylation to facilitate the transcriptional activity of STAT3-targeted astrocyte-specific genes and then promotes astrocyte differentiation of neural stem/precursor cells [36]. In germinal center B cells, PRMT5 is a JAK-binding and JAK1- and JAK3-related arginine methyltransferase that regulates STAT6 activity in response to IL-4 stimulation [37]. In another study, authors identified that PRMT2 catalyzes H3R8me2a (H3R8 asymmetric methylation) on the promoter of BCL2 and then improves the accessibility toward STAT3, which has been shown to promote BCL2 transcription [38]. However, the relationship between PRMT2 and the arginine methylation of STAT1/STAT6 requires further investigation. Likewise, whether arginine methylation affects STAT1/STAT6 phosphorylation also needs further exploration.
Conclusions
To summarize, our results indicate that PRMT2 regulates macrophage polarization in vitro. Silencing of PRMT2 promotes M1 polarization mediated by activating STAT1 together with attenuating M2 polarization by inhibition of STAT6. Notably, our findings may provide a potential target for controlling macrophage polarization-associated diseases.
Methods
BMDMs isolation and culture
Mice on a C57BL/6 background were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. BMDMs were isolated from 8-12-week-old mice. Briefly, mice were anesthetized by intraperitoneal injection with avertin (20 µl per gram of body weight). The mice lost consciousness and were unresponsive by using forceps to clamp their toes after about 2 min. Then mice were euthanized by cervical dislocation, and bone marrow of femurs and tibias was taken and incubated in DMEM (BasalMedia) containing 20 ng/ml M-CSF (Peprotech), 10% FBS (fetal bovine serum, Gibco) and 1% penicillin/streptomycin (Gibco) for 7 days at 37 °C with 5% carbon dioxide then matured into a resting state of M0 cells for subsequent stimulation.
Macrophage polarization
BMDMs were seeded at 1 × 106 cells per well in 6-well plates. Then, cells were treated with LPS (100 ng/ml, sigma, O55: B5) and IFN-γ (20 ng/ml, Peprotech) for 24 h to generate M1 macrophages or exposed to IL-4 (20 ng/mL, Peprotech) for 24 h for the generation of M2 macrophages. M0 was untreated and served as the negative control.
Reagents and antibodies
DMEM was purchased from BasalMedia. Penicillin/streptomycin and FBS were acquired from Gibco. TRIzol reagent was obtained from Thermofisher. 5x HiScript II Q RT SuperMix and 2x AceQ Universal SYBR qPCR Master Mix were purchased from Vazyme. Recombinant murine M-CSF, IFN-γ, and IL-4 protein were purchased from Peprotech. The antibodies are listed as follows: p-STAT1 (Abclonal), STAT1 (Abclonal), p-STAT3 (Abclonal), STAT3 (Cell Signaling Technology, CST), p-STAT6 (Abcam), STAT6 (Abclonal), p-ERK (CST), ERK (CST), p-JNK (CST), JNK (CST), p-P65 (CST), P65 (CST), p-P38 (CST), P38 (Abclonal), iNOS (CST), Arg1 (CST), PRMT2 (Novus), β-actin (CST), PE anti-mouse F4/80 (Biolegend), FITC anti-mouse CD86 (Biolegend), APC anti-mouse CD206 (Biolegend).
RNA interference
BMDMs were seeded and adhered into 6-well plates at 1 × 106 cells per well, then transfected with 50 nM Prmt2 small interfering RNA (Prmt2 siRNA) duplexes (with the following siRNA sequences: 5’-CGGGUUCUGUUGUGUUACATT-3’) using jetPRIME® (Polyplus) for 48 h, according to the manufacturer’s instructions. BMDMs transfected with an equal amount of universal nontargeting siRNA (sequences: 5’-UUCUCCGAACGUGUCACGUTT-3’) were regarded as negative control (NC).
Western blot
The total protein of BMDMs was extracted using RIPA (Beyotime) lysis buffer consisting of 1x protease and phosphatase inhibitors cocktail (Beyotime). Protein lysate concentrations were decided using BCA Protein Assay Kit (EpiZyme). 10% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE, EpiZyme) electrophoresis was used to separate proteins, which were subsequently transferred onto polyvinylidene fluoride membranes (PVDF, Millipore) according to the standard procedures. After blocking the membranes with 5% milk (EpiZyme) diluted in TBST at room temperature for 1 h, use the primary antibodies to submerge the membranes and incubate them overnight at 4 °C. An HRP (horseradish peroxidase-conjugated) goat anti-rabbit IgG (H + L) (1:5000, Beyotime) and anti-mouse IgG (H + L) (1:2000, CST) were used as secondary antibodies. All signals were detected using Super ECL Detection Reagent (Yeasen) according to the manufacturer’s instructions by the Amersham™ Imager 600.
Quantitative polymerase chain reaction (qPCR)
The total RNA of BMDMs was extracted using TRIzol reagent (Thermofisher) and performed reverse transcription with 5x HiScript II Q RT SuperMix (Vazyme). Real-time qPCR amplification of reverse transcription products was performed using 2x AceQ Universal SYBR qPCR Master Mix (Vazyme). The primers of each cytokine and gene are displayed in Table 1.
Flow cytometry (FCM)
BMDMs were digested by trypsin into single-cell suspensions and then used for subsequent staining. After cells were incubated with Fc receptor blocker (1:500) in the dark under 4 °C for 30 min, samples were subjected to antibodies staining at 4 °C for 30 min, rinsed twice, and resuspended in FACS buffer. Subsequently, these samples were subjected to FCM (Beckman) using different antibodies to analyze samples. PE-labeled anti-mouse F4/80, FITC-labeled anti-mouse CD86, and APC-labeled anti-mouse CD206 were stained to perform the analysis.
Statistical analysis
Results were shown by mean ± SD. All data were analyzed by a Student’s t-test using Prism 9.0. Statistical values where p < 0.05 were defined as statistically significant.
Data Availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- PRMT2:
-
Protein Arginine Methyltransferase 2
- BMDMs:
-
Bone Marrow-derived Macrophages
- IFN-γ:
-
Interferon-γ
- qPCR:
-
Quantitative Polymerase Chain Reaction
- FCM:
-
Flow Cytometry
- STAT1:
-
Signal Transducer and Activator of Transcription 1
- TNF-α:
-
Tumor Necrosis Factor-α
- IL-4:
-
Interleukin-4
- iNOS:
-
Inducible Nitric Oxide Synthase
- CD86:
-
Cluster of Differentiation 86
- FIZZ1:
-
Found in Inflammatory Zone 1
- Arg1:
-
Arginase 1
- TGF-β:
-
Transforming Growth Factorβ
- PTMs:
-
Post-translational Modifications
- MMA:
-
Mono-methyl-arginine
- aDMA:
-
Asymmetric Dimethyl-arginine
- sDMA:
-
Symmetric Dimethyl-arginine
- NF-κB:
-
Nuclear Factor kappa B
- Nos2:
-
Nitric Oxide Synthase 2
- MAPK:
-
Mitogen-activated Protein Kinase
- ERK:
-
Extracellular-signal-regulated Kinase
- JNK:
-
C-Jun N-terminal Kinase
- JAK:
-
Janus Kinase
References
Orecchioni M, Ghosheh Y, Pramod AB, Ley K. Macrophage polarization: different gene signatures in M1(LPS+) vs. classically and M2(LPS-) vs. alternatively activated macrophages. Front Immunol. 2019;10:1084. https://doi.org/10.3389/fimmu.2019.01084.
Locati M, Curtale G, Mantovani A, Diversity. Mechanisms, and significance of macrophage plasticity. Annu Rev Pathol. 2020;15:123–47. https://doi.org/10.1146/annurev-pathmechdis-012418-012718.
Heap RE, Marin-Rubio JL, Peltier J, Heunis T, Dannoura A, Moore A, Trost M. Proteomics characterisation of the L929 cell supernatant and its role in BMDM differentiation. Life Sci Alliance. 2021;4(6). https://doi.org/10.26508/lsa.202000957.
Cheng P, Li S, Chen H. Macrophages in Lung Injury, Repair, and fibrosis. Cells. 2021;10(2). https://doi.org/10.3390/cells10020436.
Ploner C, Rauchenwald T, Connolly CE, Joehrer K, Rainer J, Seifarth C, Hermann M, Nagl M, Lobenwein S, Wilflingseder D, et al. Oxidant therapy improves adipogenic differentiation of adipose-derived stem cells in human wound healing. Stem Cell Res Ther. 2021;12(1):280. https://doi.org/10.1186/s13287-021-02336-3.
Cai L, Li D, Feng Z, Gu X, Xu Q, Li Q. YTHDF2 Regulates Macrophage Polarization through NF-kappaB and MAPK Signaling Pathway Inhibition or p53 Degradation. Dis Markers. 2022; 2022:3153362. https://doi.org/10.1155/2022/3153362.
Wang S, Xu S, Zhou J, Zhang L, Mao X, Yao X, Liu C. Luteolin transforms the polarity of bone marrow-derived macrophages to regulate the cytokine Storm. J Inflamm (Lond). 2021;18(1):21. https://doi.org/10.1186/s12950-021-00285-5.
Hitosugi T, Chen J. Post-translational modifications and the Warburg effect. Oncogene. 2014;33(34):4279–85. https://doi.org/10.1038/onc.2013.406.
Price OM, Thakur A, Ortolano A, Towne A, Velez C, Acevedo O, Hevel JM. Naturally occurring cancer-associated mutations disrupt oligomerization and activity of protein arginine methyltransferase 1 (PRMT1). J Biol Chem. 2021;297(5):101336. https://doi.org/10.1016/j.jbc.2021.101336.
Bryant JP, Heiss J, Banasavadi-Siddegowda YK. Arginine methylation in Brain tumors: Tumor Biology and therapeutic strategies. Cells. 2021;10(1). https://doi.org/10.3390/cells10010124.
Guccione E, Richard S. The regulation, functions and clinical relevance of arginine methylation. Nat Rev Mol Cell Biol. 2019;20(10):642–57. https://doi.org/10.1038/s41580-019-0155-x.
Blanc RS, Richard S. Arginine methylation: the coming of Age. Mol Cell. 2017;65(1):8–24. https://doi.org/10.1016/j.molcel.2016.11.003.
Maron MI, Casill AD, Gupta V, Roth JS, Sidoli S, Query CC, Gamble MJ, Shechter D. Type I and II PRMTs inversely regulate post-transcriptional intron detention through sm and CHTOP methylation. Elife. 2022;11. https://doi.org/10.7554/eLife.72867.
Li WJ, He YH, Yang JJ, Hu GS, Lin YA, Ran T, Peng BL, Xie BL, Huang MF, Gao X, et al. Profiling PRMT methylome reveals roles of hnRNPA1 arginine methylation in RNA splicing and cell growth. Nat Commun. 2021;12(1):1946. https://doi.org/10.1038/s41467-021-21963-1.
Tewary SK, Zheng YG, Ho MC. Protein arginine methyltransferases: insights into the enzyme structure and mechanism at the atomic level. Cell Mol Life Sci. 2019;76(15):2917–32. https://doi.org/10.1007/s00018-019-03145-x.
Hou W, Nemitz S, Schopper S, Nielsen ML, Kessels MM, Qualmann B. Arginine methylation by PRMT2 controls the functions of the actin Nucleator Cobl. Dev Cell. 2018;45(2):262–275e268. https://doi.org/10.1016/j.devcel.2018.03.007.
Dong F, Li Q, Yang C, Huo D, Wang X, Ai C, Kong Y, Sun X, Wang W, Zhou Y, et al. PRMT2 links histone H3R8 asymmetric dimethylation to oncogenic activation and tumorigenesis of glioblastoma. Nat Commun. 2018;9(1):4552. https://doi.org/10.1038/s41467-018-06968-7.
Li J, Pan X, Ren Z, Li B, Liu H, Wu C, Dong X, de Vos P, Pan LL, Sun J. Protein arginine methyltransferase 2 (PRMT2) promotes dextran sulfate sodium-induced Colitis by inhibiting the SOCS3 promoter via histone H3R8 asymmetric dimethylation. Br J Pharmacol. 2022;179(1):141–58. https://doi.org/10.1111/bph.15695.
Ganesh L, Yoshimoto T, Moorthy NC, Akahata W, Boehm M, Nabel EG, Nabel GJ. Protein methyltransferase 2 inhibits NF-kappaB function and promotes apoptosis. Mol Cell Biol. 2006;26(10):3864–74. https://doi.org/10.1128/MCB.26.10.3864-3874.2006.
Dalloneau E, Pereira PL, Brault V, Nabel EG, Herault Y. Prmt2 regulates the lipopolysaccharide-induced responses in lungs and macrophages. J Immunol. 2011;187(9):4826–34. https://doi.org/10.4049/jimmunol.1101087.
Vurusaner B, Thevkar-Nages P, Kaur R, Giannarelli C, Garabedian MJ, Fisher EA. Loss of PRMT2 in myeloid cells in normoglycemic mice phenocopies impaired regression of Atherosclerosis in diabetic mice. Sci Rep. 2022;12(1):12031. https://doi.org/10.1038/s41598-022-15349-6.
Wang J, Hua H, Wang F, Yang S, Zhou Q, Wu X, Feng D, Peng H. Arginine methylation by PRMT2 promotes IFN-beta production through TLR4/IRF3 signaling pathway. Mol Immunol. 2021;139:202–10. https://doi.org/10.1016/j.molimm.2021.08.014.
Rendra E, Riabov V, Mossel DM, Sevastyanova T, Harmsen MC, Kzhyshkowska J. Reactive oxygen species (ROS) in macrophage activation and function in Diabetes. Immunobiology. 2019;224(2):242–53. https://doi.org/10.1016/j.imbio.2018.11.010.
Wang S, Cao M, Xu S, Shi J, Mao X, Yao X, Liu C. Luteolin alters macrophage polarization to inhibit inflammation. Inflammation. 2020;43(1):95–108. https://doi.org/10.1007/s10753-019-01099-7.
Cura V, Cavarelli J, Structure. Activity and function of the PRMT2 protein arginine methyltransferase. Life (Basel). 2021;11(11). https://doi.org/10.3390/life11111263.
Furqan M, Akinleye A, Mukhi N, Mittal V, Chen Y, Liu D. STAT inhibitors for cancer therapy. J Hematol Oncol. 2013;6:90. https://doi.org/10.1186/1756-8722-6-90.
Ashrafizadeh M, Ahmadi Z, Kotla NG, Afshar EG, Samarghandian S, Mandegary A, Pardakhty A, Mohammadinejad R, Sethi G. Nanoparticles targeting STATs in Cancer Therapy. Cells. 2019;8(10). https://doi.org/10.3390/cells8101158.
Hu X, Herrero C, Li WP, Antoniv TT, Falck-Pedersen E, Koch AE, Woods JM, Haines GK, Ivashkiv LB. Sensitization of IFN-gamma Jak-STAT signaling during macrophage activation. Nat Immunol. 2002;3(9):859–66. https://doi.org/10.1038/ni828.
Yu T, Gan S, Zhu Q, Dai D, Li N, Wang H, Chen X, Hou D, Wang Y, Pan Q, et al. Modulation of M2 macrophage polarization by the crosstalk between Stat6 and Trim24. Nat Commun. 2019;10(1):4353. https://doi.org/10.1038/s41467-019-12384-2.
Gong M, Zhuo X, Ma A. STAT6 Upregulation promotes M2 macrophage polarization to suppress Atherosclerosis. Med Sci Monit Basic Res. 2017;23:240–9. https://doi.org/10.12659/msmbr.904014.
Szanto A, Balint BL, Nagy ZS, Barta E, Dezso B, Pap A, Szeles L, Poliska S, Oros M, Evans RM, et al. STAT6 transcription factor is a facilitator of the nuclear receptor PPARgamma-regulated gene expression in macrophages and dendritic cells. Immunity. 2010;33(5):699–712. https://doi.org/10.1016/j.immuni.2010.11.009.
Piccolo V, Curina A, Genua M, Ghisletti S, Simonatto M, Sabo A, Amati B, Ostuni R, Natoli G. Opposing macrophage polarization programs show extensive epigenomic and transcriptional cross-talk. Nat Immunol. 2017;18(5):530–40. https://doi.org/10.1038/ni.3710.
Mowen KA, Tang J, Zhu W, Schurter BT, Shuai K, Herschman HR, David M. Arginine methylation of STAT1 modulates IFNalpha/beta-induced transcription. Cell. 2001;104(5):731–41. https://doi.org/10.1016/s0092-8674(01)00269-0.
Chen W, Daines MO, Hershey GK. Methylation of STAT6 modulates STAT6 phosphorylation, nuclear translocation, and DNA-binding activity. J Immunol. 2004;172(11):6744–50. https://doi.org/10.4049/jimmunol.172.11.6744.
Rho J, Choi S, Seong YR, Choi J, Im DS. The arginine-1493 residue in QRRGRTGR1493G motif IV of the Hepatitis C virus NS3 helicase domain is essential for NS3 protein methylation by the protein arginine methyltransferase 1. J Virol. 2001;75(17):8031–44. https://doi.org/10.1128/jvi.75.17.8031-8044.2001.
Honda M, Nakashima K, Katada S. PRMT1 regulates astrocytic differentiation of embryonic neural stem/precursor cells. J Neurochem. 2017;142(6):901–7. https://doi.org/10.1111/jnc.14123.
Igarashi H, Kuwahara K, Yoshida M, Xing Y, Maeda K, Nakajima K, Sakaguchi N. GANP suppresses the arginine methyltransferase PRMT5 regulating IL-4-mediated STAT6-signaling to IgE production in B cells. Mol Immunol. 2009;46(6):1031–41. https://doi.org/10.1016/j.molimm.2008.08.272.
Hu G, Yan C, Xie P, Cao Y, Shao J, Ge J. PRMT2 accelerates tumorigenesis of hepatocellular carcinoma by activating Bcl2 via histone H3R8 methylation. Exp Cell Res. 2020;394(2):112152. https://doi.org/10.1016/j.yexcr.2020.112152.
Acknowledgments
Not applicable.
Funding
This work was supported by National Natural Science Foundation of China [Grant No. 82101840, HJ.H., No. 82102259, YJ.L. and No. 81871101, Y.L.].
Author information
Authors and Affiliations
Contributions
Y.L. and HJ.H. conceived and supervised the study. T.L., YJ.L. and MQ.X. performed the experiments. Y.L., HJ.H., and T.L. analyzed the data and wrote the manuscript. All authors contributed to discussing the results.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal care and experimental procedures were conducted according to the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals, with the approval of the Animal Experimentation Ethics Committee of Ruijin Hospital (Shanghai, China). All methods were performed in accordance with the relevant guidelines and regulations and were reported in compliance with the ARRIVE guidelines (https://arriveguidelines.org).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, T., Li, Y., Xu, M. et al. PRMT2 silencing regulates macrophage polarization through activation of STAT1 or inhibition of STAT6. BMC Immunol 25, 1 (2024). https://doi.org/10.1186/s12865-023-00593-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12865-023-00593-w