
Abstract
Background
Dinosaur eggs containing embryos are rare, limiting our understanding of dinosaur development. Recently, a clutch of subspherical dinosaur eggs was discovered while blasting for a construction project in the Upper Cretaceous red beds (Hekou Formation) of the Ganzhou Basin, Jiangxi Province, China. At least two of the eggs contain identifiable hadrosauroid embryos, described here for the first time.
Results
The eggs, attributable to Spheroolithidae indet., are thin-walled and small (~ 660 mL) compared to those of Lambeosaurinae. The shape of the embryonic squamosal is reminiscent of that seen in the Late Cretaceous hadrosauroids Levnesovia transoxiana, Tanius sinensis, and Nanningosaurus dashiensis, suggestive of possible affinities.
Conclusion
The small size of the eggs and embryos, similar to those of Hadrosaurinae, indicates that the larger eggs and hatchlings typical of Lambeosaurinae are evolutionarily derived.
Similar content being viewed by others


Background
Dinosaur eggs are common worldwide, but embryos are rare [1]. Among the diverse duck-billed dinosaurs and their nearest relatives (Hadrosauroidea), just three identifiable species are known from perinatal material: Hypacrosaurus stebingeri [2], Maiasaura peeblesorum [3, 4], and Saurolophus angustirostris [5]. Accordingly, the early ontogeny of hadrosauroids is poorly understood, which hinders determination of skeletal development and allometric trends across the clade.
Recently, a construction project in the Upper Cretaceous red beds of the Ganzhou Basin, Jiangxi Province, China revealed a fossilized clutch of spheroolithid eggs. In this contribution, we briefly describe two of these eggs and their embryonic contents, accessioned at the Yingliang Stone Natural History Museum (YLSNHM) in Fujian Province, China. The embryos (YLSNHM 01328 and 01373) share several features in common with hadrosauroids, but otherwise lack the more derived features present in embryonic hadrosaurids. The squamosal is distinctive and recalls that of some other Late Cretaceous hadrosauroids, the implications of which we explore here. We end this contribution with a consideration of the evolutionary and taphonomic implications of these eggs and their contents.
A comment on taxonomy
Hadrosauridae is traditionally split into two subfamilies, the solid-crested Hadrosaurinae and hollow-crested Lambeosaurinae [6], the former named after the eponymous Hadrosaurus foulkii. However, in a relatively recent phylogenetic analysis, Prieto-Marquez [7] recovered Hadrosaurus as the sister taxon to all other hadrosaurids, and so, abiding by the regulations of the International Code of Zoological Nomenclature, renamed the clade of solid-crested hadrosaurids Saurolophinae. Although this new name has gained some acceptance in the literature (e.g., [8,9,10]), not all current phylogenies recover Hadrosaurus apart from the solid-crested hadrosaurids (e.g., [11, 12]). Therefore, in keeping with tradition and some recent phylogenies, we use the clade name Hadrosaurinae, which is practically synonymous with Saurolophinae, except for the inclusion of Hadrosaurus [13].
Geological provenance
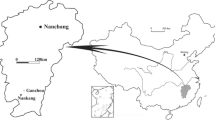
The red beds of the Ganzhou area of Jiangxi Province are divided into: (1) the Upper Cretaceous Ganzhou Group, consisting of the Maodian and Zhoutian formations, and (2) the Cretaceous-Paleogene Guifeng Group, consisting of the Upper Cretaceous Hekou and Tangbian formations and the Cretaceous-Paleogene Lianhe Formation [14]. The eggs and embryos reported here come from the Hekou Formation of the Guifeng Group in the Ganzhou area (Fig. 1).

Map showing location of the embryo-bearing egg specimens, YLSNHM 01328 and 01373 (“Fossil site”) in southern China
The Hekou Formation varies in thickness between 200 and 1600 m, depending on where it is exposed. It consists of channelized, fining-upward conglomerates, sieve deposits, and alternating sandstone and mudstone beds. The depositional setting is interpreted as primarily fluvially-derived within a proximal alluvial fan system. Mudcracks and caliche deposits are common, and the prevailing palaeoclimate is interpreted as subhumid [15].
The age of the Hekou Formation is debated. Some have argued for a Coniacian-Santonian age for the formation [16], but a Maastrichtian age is more commonly accepted. The latter interpretation is based partly on palaeomagnetic studies that have dated the host Guifeng Group to 71.4–65.0 Ma [17, 18]. Some have also argued that the Guifeng Group is penecontemporaneous with the red beds of the nearby Nanxiong Group [19], which is purportedly of Maastrichtian age [20]. In this model, the Hekou Formation is thought to correlate with the Dafeng and Yuanpu formations of the lower Nanxiong Group [21]. Indeed, the preserved theropod assemblage of the Nanxiong Group, including alioramine tyrannosaurids and oviraptorosaurs [22, 23], agrees well with the dinosaur assemblage of the Nemegt Formation in Mongolia, which itself is often considered Maastrichtian in age [24]. For these reasons, and in keeping with the recent literature (e.g., [25, 26]), we accept a probable Maastrichtian age for the Hekou Formation.
The Hekou Formation has thus far yielded fossil algae, plants, dinosaur bones, and trackways, few of which have been formally described [14, 25,26,27,28,29]. Dinosaur eggs assigned to Oolithes sp., Oolithes spheroides [30], “Spheroolithus minor”, Ovaloolithus sp., Paraspheroolithus sp., Macroolithus rugustus, Coelurosauria fam. et gen. indet. [27], and Elongatoolithidae [26] are also known from the Hekou Formation.
Results
Description of eggs
Each embryo described here is preserved within its respective egg, both of which came from the same egg clutch. Field records note the presence of at least 13 eggs in the clutch at the time of excavation, but the original number may have been higher (S Miao, pers. comm. to LX, 2008).
The better-preserved egg of YLSNHM 01373 has the shape of a prolate spheroid (Fig. 2A), comparable to the spherical-subspherical eggs assigned to the oofamily Spheroolithidae [31], which has been reported previously from the Hekou Formation as “Spheroolithus minor” [27], a nomen nudum. Given the cross-sectional dimensions of the egg, we estimate a total egg volume of approximately 600 mL. Viewed in cross-section, the embryo fills approximately 40% of the egg by area.

Eggshell of Spheroolithidae sp. (YLSNHM 01373). A Overview of egg containing embryonic hadrosauroid; B cross-section of the YLSNHM 01373 eggshell under transmitted, unpolarized light. The dotted line marks the boundary between the mammillary (ML) and continuous (CL) layers. The white arrows indicate the locations of organic cores
The eggshell of YLSNHM 01373 is poorly preserved and thus hinders our detailed description and further assignment to an oogenus. While hadrosaurids have long been associated with spheroolithid eggs based on their embryonic contents [2, 4, 32, 33], the association remains questionable due to the poor preservation of the eggshell and the absence of detailed description of the eggshells in previous reports [e.g., 2, 4]. The YLSNHM 01373 egg reveals an indistinct boundary between the mammillary and continuous layers (Fig. 2B), indicating its two-layered structure—a common microscopic feature for dinosaurian eggs. The organic cores (nucleation site for the acicular radial crystal growth in the lower part) are poorly circumscribed, probably due to severe embryo-induced erosion or poor preservation (or both). The radial crystals merge into a tabular structure in the external quarter of the eggshell. The eggshell of YLSNHM 01373 ranges 0.32–0.42 mm thick, among the thinnest known for the oofamily [34].
Description of embryos
YLSNHM 01328
This partial, articulated skeleton consists of the posterior cranium (missing most of the snout), complete cervical series, and the anterior-most dorsal vertebrae and associated ribs (Fig. 3). The bridge of the rostrum has buckled; its original profile is obscured. The elongate, paired nasals are unfused and broken anteriorly where they reach their greatest transverse breadth. In lateral profile, the nasal is subtly bowed dorsally above the naris. The squamosal has disarticulated from the postorbital. The postorbital process of the squamosal is tall, blunt, and constricts where it meets the main body of the element. The pre- and postcotylar processes are subequal in length. The squamosal compares most favourably with those of the hadrosauroids Tanius sinensis [35], Levnesovia transoxiana [36], and Nanningosaurus dashiensis [37]. By contrast, the postorbital process of the squamosal is much longer and slenderer in most other hadrosauroids, including Gobihadros mongoliensis [38], Prosaurolophus maximus [39], and Corythosaurus casuarius [40] (Fig. 4). The anterior third of the left maxilla is missing, whereas the preserved portion is 14 mm long. Whether a palatal process of the maxilla (sensu [41])—whose absence is diagnostic of Lambeosaurinae—originally existed cannot be determined. The jugal facet of the maxilla faces laterally, and its long axis is horizontal as in the perinates of Maiasaura peeblesorum, whereas the same facet is distinctly angled in those of Hypacrosaurus stebingeri [2]. The partial maxilla has eight teeth in situ (there is a total 12 maxillary teeth in embryonic H. stebingeri; [2]). Each has a straight primary (median) ridge, offset slightly distally, and lacks both subsidiary ridges and marginal denticles (Fig. 5), as in the perinates of M. peeblesorum [42]. The occlusal surfaces of the teeth are not visible, hindering determination of whether they bear wear facets as they do in the embryos of H. stebingeri [2]. The quadrate (17 mm tall) is robust with a broadly rounded head that articulates dorsally with the squamosal in a hinge joint. The pterygoid flange is broad, and the quadratojugal notch, largely obscured by the quadratojugal, occurs in the lower half of the quadrate body. The quadrate is gently bowed anteriorly along its length; the dorsal and ventral halves form an angle of approximately 154°. The ossified braincase elements are unfused and have been displaced; the basisphenoid now rests against the skull roof and the orbitosphenoid has shifted posteriorly. The prootic is visible beneath the basisphenoid, where the single opening for c.n. V (trigeminal nerve) can be seen. The otoccipital (opisthotic + exoccipital) occurs further posteriorly. On the occiput, the well-developed paroccipital process is pendant and projects posterolaterally beneath the squamosal.

Hadrosauroid partial embryonic skeleton (YLSNHM 01328). A Photograph, B interpretive drawing. See text for list of abbreviations

Hadrosauriform squamosals in left lateral view. Iguanodon bernissartensis after [58], Gobihadros mongoliensis after [38], Levnesovia transoxiana after [36], Nanningosaurus dashiensis after [37], Tanius sinensis after [35], Prosaurolophus maximus after [39], Corythosaurus casuarius after [40]. See text for list of abbreviations

Occluded teeth of YLSNHM 01328. Maxillary teeth (top row) exhibit a strong primary (median) ridge and no subsidiary ridges or marginal denticles
The cervical series is nearly complete and preserved in a gentle sigmoid along its length, being dorsiflexed anteriorly and ventroflexed posteriorly. The neurocentral sutures are unfused. The neural spines are low, whereas the postzygapophyses are strongly developed and hooked, both of which are common to all hadrosauroids. The postzygapophyses do not extend above the level of the neural spines as they do in adult hadrosaurids (e.g., [43, 44]). Many of the transverse processes have buckled ventrally, obscuring their corresponding centra. The few remaining cervical ribs are L-shaped, with elongate posterior processes. The transition between the cervical and anterior dorsal series is nearly indistinguishable. We tentatively identify the first dorsal as that bearing the first preserved elongate rib, which itself is broken along its length. As such, we identify 12 cervical vertebrae, although the count may have been higher by one or two vertebrae (depending on whether the first preserved long rib reached the sternum or not). For comparison, the cervical series contains 11 vertebrae in most known non-hadrosaurid hadrosauroids [38, 45, 46], and varies between 12 and 18 vertebrae in hadrosaurids [6, 47]. The neural spines of YLSNHM 01328 increase in size and posterior inclination by the 15th presacral vertebra, but the dorsal series is not preserved beyond the 17th presacral vertebra.
YLSNHM 01373
This articulated skeleton is lacking parts of the skull, distal limb elements, and tail (Fig. 6). The parietal, which is poorly visible in YLSNHM 01328, is elongate (2.7 times longer than wide), as in non-lambeosaurine hadrosauroids. A partial tooth row has been displaced from the jaws and now lies adjacent to the similarly displaced left ilium. The dissociation of the teeth from their host element makes it difficult to determine whether they originated within the maxilla or dentary. The morphology of the tooth crowns agrees with that of the maxillary teeth of YLSNHM 01328. The preserved cervical and dorsal vertebrae show unfused neural arches. We consider the first dorsal vertebra to be that bearing the first long rib that presumably connected with the sternum, in which case, we count 11 cervical vertebrae and 18 dorsal vertebrae. However, we can neither confirm the presence of the atlas/axis in the preserved series nor rule out the possibility that the posteriormost free dorsal vertebra would eventually become incorporated in the synsacrum as a dorsosacral [6]; the true cervical count could be higher. The few preserved caudal vertebrae show that the neural spines were low. The coracoid foramen is not enclosed but opens posteriorly to separate the contact surfaces for the humerus and scapula, as in Hypacrosaurus stebingeri perinates [2]. The scapular blade is slender and the caudal end is irregular and poorly ossified. The length of the deltopectoral crest of the humerus is moderate compared to the length of the humerus (ratio = 0.53), which contrasts with the condition of most hadrosaurids (ratio > 0.55) [6, 7]. The distal condyles of the humerus are poorly defined. The left ilium has drifted anteriorly to be preserved alongside the skull. It is low in lateral profile, with a preacetabular blade that is only weakly arched dorsally. The pubic and ischiadic penduncles are poorly defined. The femur is 26 mm long, with a well-developed greater trochanter and poorly defined distal condyles. The fourth trochanter is not visible, on account of the outward rotation of the femur. The preserved tibia and fibula are missing their extremities.

Hadrosauroid partial embryonic skeleton (YLSNHM 01373). A Photograph, B interpretive drawing. See text for list of abbreviations
Discussion and conclusions
The embryos described here can be confidently assigned to Hadrosauroidea based on the following derived features (from [7]): (1) large jugal contact of maxilla faces strongly laterally; (2) deltopectoral crest of humerus wide relative to minimum width of humeral shaft (ratio = 1.71). Dentary teeth having a centrally located primary ridge and lacking subsidiary ridges are also considered derived for hadrosauroids [7], and while these features are present on the maxillary teeth of the embryos described here, we are unable to confirm their occurrence specifically on the dentary teeth (the dentary tooth crowns are not visible in YLSNHM 01328, and the loose teeth of YLSNHM 01373 are not definitively from the dentary).
Derived features that would allow the embryos to be assigned to Hadrosauridae (see synapomorphies listed in [7]) are lacking. For example, in hadrosaurid adults and embryos, the maxilla often features a well-defined ectopterygoid ridge [2, 36], which is missing in YLSNHM 01328. Hadrosaurids of all stages of development also possess a deltopectoral crest of the humerus that is both proximodistally elongate and exhibits a pointed distal corner [2, 5, 42, 48]; this is not the case in YLSNHM 01373. The parietal and nasals are also long compared to the condition seen in lambeosaurines (although parietal length is negatively allometric over the course of lambeosaurine ontogeny; [7]). Many definitive hadrosaurid characters are associated with the ilium. However, these characters typically relate to the pubic and ischial peduncles that bound the acetabulum, which is poorly preserved in YLSNHM 01373, and so these characters cannot be properly assessed. The relatively high cervical count (12 +) that we report in the embryos is consistent with that of most hadrosaurids [6], but cervical count does not unambiguously diagnose Hadrosauridae [7].
The squamosal of YLSNHM 01328 is distinctive in having a relatively tall, blunt postorbital process, which is also seen in the hadrosauroids Levnesovia transoxiana, Nanningosaurus dashiensis, and Tanius sinensis (Fig. 4). Further comparison with these taxa is complicated by the fact that none are very complete and that all are known from osteologically mature individuals. The holotype of Levnesovia transoxiana is represented by a partial skull and scattered postcranial elements [36]. Most notably, the teeth differ from those of the embryos described here in having denticulate crown margins and weak secondary ridges (the latter on the dentary teeth only). The holotype of Nanningosaurus dashiensis consists of scant skull and postcranial remains [37]. The dorsal process of the maxilla differs from that of YLSNHM 01328 in being sharply peaked. The maxillary teeth primarily differ from those reported here in being more numerous and narrower mesiodistally, both characters known to vary with age in the hadrosaurid Hypacrosaurus stebingeri [2]. The dentary teeth of N. dashiensis bear secondary ridges and marginal denticles, but these features cannot be confirmed in YLSNHM 01328 because the dentary tooth crowns are obscured by the overlying maxillary teeth. The isolated tooth row preserved with YLSNHM 01373 cannot be confidently ascribed to the dentary to facilitate comparison with N. dashiensis. The holotype of Tanius sinensis consists of a posterior cranium, varied appendicular bones, a series of ten cervical vertebrae (complete count unknown), and a few other axial elements [35]. No extant diagnosis exists for the cranium, and the postcranium is relatively conservative. The dorsal neural spines are purportedly tall for a non-hadrosaurid hadrosauroid [49], but neural spine height is known to increase with age in H. stebingeri [2]. The poor temporal resolution of the host Hekou Formation makes it difficult to determine whether the described embryos were penecontemporaneous with any of the other Late Cretaceous hadrosauroid taxa just mentioned (and whose temporal ranges are sometimes likewise poorly constrained). If we accept that the Hekou Formation is of Maastrichtian age (see ‘Geological provenance’ above), then the hadrosauroid embryos may only be penecontemporaneous with T. sinensis, which itself is from somewhere within the upper Campanian to lower Maastrichtian [49].The occurrence of hadrosauroids in the Hekou Formation is not unprecedented. Their presence was initially signaled following the description of large ornithopod tracks in Upper Cretaceous red beds elsewhere in southern China [50, 51]. More recently, Xing et al. [25] described a partial axial skeleton of a hadrosaurid from the Hekou Formation, identified on the basis of the long and robust postzygapophyses of the cervical vertebrae. Similarly developed processes are not present in the embryos described here, supporting the existence of at least two hadrosauroid taxa within the Hekou Formation—a hadrosaurid and a non-hadrosaurid hadrosauroid.
In describing hadrosaurine and lambeosaurine eggs and embryos from Montana, Horner [4] noted that those of hadrosaurines tend to be much smaller than those of lambeosaurines. Hadrosaurine embryos (Maiasaura peeblesorum) have femora that vary in length between 35 and 40 mm, and their subspherical eggs were calculated to have a volume of approximately 900 mL. Lambeosaurine embryos (Hypacrosaurus stebingeri and an indeterminate form), by contrast, have femora that vary between 60 and 80 mm long, and are derived from eggs approaching 4000 mL [2, 4]. Horner [4] hypothesized that these differences were typical of their respective subfamilies, and further suggested that the smaller hadrosaurine hatchlings were altricial, based on their poorly ossified epiphyses [3, 52].
The hadrosauroid eggs and embryos reported here facilitate character polarization of egg and hatchling size among hadrosaurids. The 26 mm femur of YLSNHM 01373 is closer to M. peeblesorum perinates in size, and the corresponding egg (660 mL) similarly is more like those of M. peeblesorum than to those of any known lambeosaurine [4]. Importantly, the YLSNHM 01373 embryo is not fully developed, evidenced by the fact that the skeleton does not entirely fill the egg [cf. 54]. This likely explains the missing ends of the tibia-fibula, which ossify from the diaphyses outward, and the absence of many of the manual and pedal elements in an otherwise mostly undisturbed skeleton; they may simply not have ossified by the time of death [54]. It is probable, therefore, that these embryos were yet several embryonic stages away from hatching (Fig. 7). Consequently, we are unable to determine whether the corresponding neonates were altricial, given the incomplete development of the embryo. However, in the closely related Telmatosaurus transsylvanicus, the hatchlings are similarly small and their limb bone epiphyses are poorly formed [55], as in M. peeblesorum. These observations strongly imply that the lambeosaurine condition of having larger eggs and precocial hatchlings is an evolutionarily derived trait.

Hadrosauroid embryo, based on YLSNHM 01373
Dinosaur eggs and embryos commonly occur in semi-arid, upland palaeoenvironments [1]. Palaeontologists once maintained that these were the preferred nesting grounds of dinosaurs [32, 56], but rare perinatal bones have also been found in lowland deposits (e.g., [48, 57], and it is now widely held that the wet, acidic conditions of such palaeoenvironments exerted a bias against the preservation of eggs and their contents [57]. The Hekou Formation of China, with its diversity of fossil dinosaur eggs combined with sedimentological indicators of moderately dry, well-drained conditions (e.g., caliche, mudcracks, carbonate nodules), is entirely consistent with this framework of understanding. These strata promise to reveal many more clues about early ontogenetic development in dinosaurs.
Methods
Eggshell histology
A piece of eggshell was removed from YLSNHM 01373 with an Engraving Pen AT-310. The shell was embedded in Araldite 2020, cut with a STX-202A diamond wire cutting machine, and then polished with P400 to P4000 abrasive paper to approximately 30 μm thick for microscopic observation under normal and cross-polarized light with a Zeiss Primotech microscope.
Availability of data and materials
All data generated or analysed during this study are included in this published article.
Abbreviations
- atic:
-
Atlas intercentrum
- ax:
-
Axis
- bsp:
-
Basisphenoid
- ca:
-
Caudal vertebrae
- CL:
-
Continuous layer
- cnV:
-
Cranial nerve V exit
- co:
-
Coracoid
- cv:
-
Cervical vertebrae
- ?d1:
-
First dorsal vertebra (possibly)
- dto:
-
Dentary tooth
- dr:
-
Dorsal rib
- dv:
-
Dorsal vertebrae
- f:
-
Frontal
- fe:
-
Femur
- fl:
-
Fibula
- h:
-
Humerus
- il:
-
Ilium
- is:
-
Ischium
- j:
-
Jugal
- mt:
-
Metatarsal
- ML:
-
Mammillary layer
- mx:
-
Maxilla
- mxto:
-
Maxillary tooth
- n:
-
Nasal
- oc:
-
Occipital condyle
- osp:
-
Orbitosphenoid
- oto:
-
Otoccipital
- p:
-
Parietal
- pl:
-
Palatine
- po:
-
Postorbital
- poc:
-
Postorbital contact
- pocp:
-
Postcotyloid process
- prcp:
-
Precotyloid process
- prf:
-
Prefrontal
- pro:
-
Prootic
- pt:
-
Pterygoid
- pu:
-
Pubis
- q:
-
Quadrate
- qj:
-
Quadratojugal
- r:
-
Radius
- sa:
-
Sacrum
- sc:
-
Scapula
- sp:
-
Swollen process of squamosal
- sq:
-
Squamosal
- t:
-
Tibia
- to:
-
Tooth
- ul:
-
Ulna
- YLSNHM:
-
Yingliang Stone Natural History Museum
References
Carpenter K, Alf K. Global distribution of dinosaur eggs, nests, and babies. In: Carpender K, Hirsch KF, Horner JR, editors. Dinosaur eggs and babies. Cambridge: Cambridge University Press; 1994. p. 15–30.
Horner JR, Currie PJ. Embryonic and neonatal morphology and ontogeny of a new species of Hypacrosaurus (Ornithischia, Lambeosauridae) from Montana and Alberta. In: Carpender K, Hirsch KF, Horner JR, editors. Dinosaur eggs and babies. Cambridge: Cambridge University Press; 1994. p. 312–36.
Horner JR, Weishampel DB. A comparative embryological study of two ornithischian dinosaurs. Nature. 1988;332:256–7.
Horner JR. Egg clutches and embryos of two hadrosaurian dinosaurs. J Vertebr Paleontol. 1999;19:607–11.
Dewaele L, Tsogtbaatar K, Barsbold R, Garcia G, Stein K, Escuillié F, Godefroit P. Perinatal specimens of Saurolophus angustirostris (Dinosauria: Hadrosauridae), from the Upper Cretaceous of Mongolia. PLoS ONE. 2015;10: e0138806.
Horner JR, Weishampel DB, Forster CA. Hadrosauridae. In: Weishampel DB, Dodson P, Osmólska H, editors. The Dinosauria. 2nd ed. Berkeley: University of California Press; 2004. p. 438–63.
Prieto-Marquez A. Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods. Zool J Linn Soc. 2010;159:435–502.
Gates TA, Scheetz R. A new saurolophine hadrosaurid (Dinosauria: Ornithopoda) from the Campanian of Utah. North America J Syst Palaeontol. 2015;13:711–25.
Zhang X-Q, Zhang X-M, Hou M-C, Li G, Li H-M. Lithostratigraphic subdivision of red beds in Nanxiong Basin, Guangdong, China. J Stratigr. 2017;37:441–51.
McDonald AT, Wolfe DG, Freedman Fowler EA, Gates TA. A new brachylophosaurin (Dinosauria: Hadrosauridae) from the Upper Cretaceous Menefee Formation of New Mexico. PeerJ. 2021;9: e11084.
Xing H, Mallon JC, Currie ML. Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae. PLoS ONE. 2017;12: e0175253.
Kobayashi Y, Nishimura T, Takasaki R, Chiba K, Fiorillo AR, Tanaka K, et al. A new hadrosaurine (Dinosauria: Hadrosauridae) from the marine deposits of the Late Cretaceous Hakobuchi Formation, Yezo Group. Japan Sci Rep. 2019;9:12389.
Madzia D, Arbour VM, Boyd CA, Farke AA, Cruzado-Caballero P, Evans DC. The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ. 2021;9: e12362.
Wen C, Liu X, Lü B, Mao X, Chen J, Hou S, Zhou Z, Hou J, Wu H. The Cretaceous redbeds in Shicheng Basin, Jiangxi province: pedogenic and paleoenvironmental characteristics. Quat Sci. 2016;36:1403–16.
Chen LQ, Steel RJ, Guo FS, Olariu C, Gong CL. Alluvial fan facies of the Yongchong Basin: implications for tectonic and paleoclimatic changes during Late Cretaceous in SE China. J Asian Earth Sci. 2017;134:37–54.
Cao K. Cretaceous terrestrial deposits in China. China Geol. 2018;3:402–14.
Gu XR. The magnetic stratum of Late Cretaceous redbeds in Jitai Basin. Jiangxi Province Geol Sci Technol Jiangxi. 1991;18:185–8.
Zuo Y, Wu J, Zhou W. Lithostratigraphic division of volcanic terrain in Late Mesozoic in Jiangxi. Geol Rev. 1999;45:742–50.
Fang X, Li P, Zhang Z, Zhang X, Lin Y, Guo S, Cheng Y, Li Z, Zhang X, Cheng Z. Cretaceous strata in Nanxiong Basin of Guangdong and the evolution from the dinosaur egg to the bird egg. Acta Geol Sin. 2009;30:167–86.
Jiangxi Bureau of Geology and Mineral Resources. Regional geology of Jiangxi Province. Beijing: Geological Publishing House; 1984.
Zhang XQ, Zhang SJ, Lin XY, Huan QJ, Li HL. Red beds and paleontology in the Nanxiong Basin. Beijing: Huaxia Literature and Art Publishing House; 2021.
Lü JC, Yi L, Brusatte SL, Yang L, Li H, Chen L. A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. Nat Commun. 2014;5:3788.
Bi SD, Romain A, de Fabrègues CP, Pittman M, Lamanna MC, Yu YL, Yu CY, et al. An oviraptorid preserved atop an embryo-bearing egg clutch sheds light on the reproductive biology of non-avialan theropod dinosaurs. Sci Bull. 2021;66:947–54.
Funston GF, Mendonca SE, Currie PJ, Barsbold R. Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin. Palaeogeogr Palaeoclimatol Palaeoecol. 2018;494:101–20.
Xing L, Niu K, Wang D, Prieto-Marquez A. A partial articulated hadrosaurid skeleton from the Maastrichtian (Upper Cretaceous) of the Ganzhou area, Jiangxi Province. China Hist Biol. 2021;33:2256–9.
Xing L, Niu K, Ma W, Zelenitsky DK, Yang T-R, Brusatte SL. An exquisitely preserved in-ovo theropod dinosaur embryo sheds light on avian-like prehatching postures. iScience. 2021;25:103516.
Peng H. Longhushan, Jiangxi. In: Peng H, editor. China Danxia. Singapore: Springer; 2020. p. 299–343.
Xing L, Li D, Klein H, Lockley MG, Liang Q, Yang J, Li L, You W. Lower Cretaceous turtle tracks from Hekou Group of northwest China. Cret Res. 2019;99:269–74.
Xing L, Niu K, Zhang L, Yang T-R, Zhang J, Persons WS IV, Romilio A, Zhuang Y, Ran H. Dinosaur eggs associated with crustacean trace fossils from the Upper Cretaceous of Jiangxi, China: evidence for foraging behavior? Biosis Biol Syst. 2020;1:54–9.
Mikhailov KE, Bray ES, Hirsch KF. Parataxonomy of fossil egg remains (Veterovata): basic principles and applications. J Vertebr Paleontol. 1996;16:763–9.
Mikhailov KE. Classification of fossil eggshells of amniotic vertebrates. Acta Palaeontol Pol. 1991;36:193–238.
Horner JR, Makela R. Nest of juveniles provides evidence of family structure among dinosaurs. Nature. 1979;282:296–8.
Hirsch KF, Quinn B. Eggs and eggshell fragments from the Upper Cretaceous Two Medicine Formation of Montana. J Vertebr Paleontol. 1990;10:491–511.
Moreno-Azanza M, Canudo JI, Gasca JM. Spheroolihid eggshells in the Lower Cretaceous of Europe: implications for eggshell evolution in ornithischian dinosaurs. Cret Res. 2014;51:75–87.
Wiman C. Die Kreide-Dinosaurier aus Shantung. Palaeontol Sin Ser C. 1929;6:1–67.
Sues HD, Averianov A. A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs. Proc R Soc B. 2009;276:2549–55.
Mo J, Zhao Z, Wang W, Xu X. The first hadrosaurid dinosaur from southern China. Acta Geol Sin. 2007;81:550–4.
Tsogtbaatar K, Weishampel DB, Evans DC, Watabe M. A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia). PLoS ONE. 2019;14: e0208480.
Horner JR. Cranial morphology of Prosaurolophus (Ornithischia: Hadrosauridae) with descriptions of two new hadrosaurid species and an evaluation of hadrosaurid phylogenetic relationships. Mus Rockies Occ Pap. 1992;2:1–119.
Ostrom JH. Cranial morphology of the hadrosaurian dinosaurs of North America. Bull Am Mus Nat Hist. 1961;122:1–186.
Wagner JR, Lehman TM. An enigmatic new lambeosaurine hadrosaur (Reptilia: Dinosauria) from the upper shale member of the Campanian Aguja Formation of Trans-Pecos Texas. J Vertebr Paleontol. 2009;29:605–11.
Prieto-Marquez A, Guenther MF. Perinatal specimens of Maiasaura from the Upper Cretaceous of Montana (USA): insights into the early ontogeny of saurolophine hadrosaurid dinosaurs. PeerJ. 2018;6: e4734.
Cuthbertson RS, Holmes RB. The first complete description of the holotype of Brachylophosaurus canadensis Sternberg, 1953 (Dinosauria: Hadrosauridae) with comments on intraspecific variation. Zool J Linn Soc. 2010;159:373–97.
Campione NE. Postcranial anatomy of Edmontosaurus regalis (Hadrosauridae) from the Horseshoe Canyon Formation, Alberta, Canada. In: Eberth DA, Evans DC, editors. Hadrosaurs: Indiana University Press; 2015. p. 208–44.
You H, Li D, Liu W. A new hadrosauriform dinosaur from the Early Cretaceous of Gansu Province. China Acta Geol Sin. 2011;85:51–7.
Dalla Vecchia FM. Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy. J Vertebr Paleontol. 2009;29:1100–16.
Godefroit P, Bolotsky Y, Alifanov V. A remarkable hollow-crested hadrosaur from Russia: an Asian origin for lambeosaurines. C R Palevol. 2003;2:143–51.
Tanke DH, Brett-Surman MK. Evidence of hatchling and nesting-size hadrosaurs (Reptilia: Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta. In: Tanke DH, Carpenter K, editors. Mesozoic vertebrate life. Bloomington: Indiana University Press; 2001. p. 206–18.
Borinder NH, Poropat SF, Campione NE, Wigren T, Kear BP. Postcranial osteology of the basally branching hadrosauroid dinosaur Tanius sinensis from the Upper Cretaceous Wangshi Group of Shandong. China J Vertebr Paleontol. 2021;41: e1914642.
Xing LD, Harris JD, Dong ZM, Lin YL, Chen W, Guo SB, Ji Q. Ornithopod (Dinosauria: Ornithischia) tracks from the Upper Cretaceous Zhutian Formation in Nanxiong Basin, China and general observations on large Chinese ornithopod footprints. Geol Bull China. 2009;28:829–43.
Xing LD, Lockley MG, Li DL, Klein H, Persons WSIV, Ye Y, Zhang JP, Ran H. Late Cretaceous ornithopod-dominated, theropod, and pterosaur track assemblages from the Nanxiong Basin, China: new discoveries, ichnotaxonomy, and palaeoecology. Palaeogeogr Palaeoclimatol Palaeoecol. 2017;466:303–13.
Horner JR. Dinosaur reproduction and parenting. Ann Rev Earth Planet Sci. 2000;28:19–45.
Ferguson M. Reproductive biology and embryology of the crocodilians. In: Gans C, Billett F, Maderson PFA, editors. Biology of the Reptilia, vol. 14. New York: Wiley; 1985. p. 329–491.
Rieppel O. Studies on skeleton formation in reptiles. V. Patterns of ossification in the skeleton of Alligator mississippiensis Daudin (Reptilia, Crocodylia). Zool J Linn Soc. 1993;109:301–25.
Grigorescu D, Csiki Z. Ontogenetic development of Telmatosaurus transsylvanicus (Ornitischia: Hadrosauria) from the Maastrichtian of the Haţeg Basin Romania-evidence from the limb bones. Hantkeniana. 2006;5:20–6.
Sternberg CMA. juvenile hadrosaur from the Oldman Formation of Alberta. Natl Mus Can Bull. 1955;136:120–2.
Carpenter K. Baby dinosaurs from the Late Cretaceous Lance and Hell Creek formations and a description of a new species of theropod. Rocky Mt Geol. 1982;20:123–34.
Norman DB. On the taxonomy and diversity of Wealden iguanodontian dinosaurs (Ornithischia: Ornithopoda). Rev Paléobiol. 2013;32:385–404.
Acknowledgements
Thanks to T. Gates, Mo J.-Y., Wu X.-C., Xu X., and D. Zelenitsky for helpful discussion. Wu R. prepared the eggshell sections. Ren W.-Y. expertly crafted the restoration in Fig. 7. Thanks also to two anonymous reviewers for constructive criticism of the manuscript, and to G. Carnevale for editorial assistance.
Funding
LX is supported by the National Natural Science Foundation of China (No. 41888101, 41790455), the 111 project (B20011), and the Fundamental Research Funds for Central Universities (265QZ201903). T-RY is supported by the research grant of Ministry of Science and Technology, Taiwan (MOST 110-2116-M-178-002). TM is supported by a Discovery Launch Supplement from the Natural Sciences and Engineering Research Council of Canada (DGECR-2021-00421). Both TM and JCM are supported by funding from the Canadian Museum of Nature.
Author information
Authors and Affiliations
Contributions
LX managed the project and photographed the eggs and embryos. KN obtained the specimens, conducted the geological survey, supervised specimen preparation, and contributed to discussions of morphology and taphonomy. T-RY contributed the eggshell histological analysis. TM produced the anatomical drawings in Figs. 3 and 6. JCM designed the figures. LX, T-RY, DW, TM, and JCM wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xing, L., Niu, K., Yang, TR. et al. Hadrosauroid eggs and embryos from the Upper Cretaceous (Maastrichtian) of Jiangxi Province, China. BMC Ecol Evo 22, 60 (2022). https://doi.org/10.1186/s12862-022-02012-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-022-02012-x