
Summary
Cryptochromes are photoreceptors that regulate entrainment by light of the circadian clock in plants and animals. They also act as integral parts of the central circadian oscillator in animal brains and as receptors controlling photomorphogenesis in response to blue or ultraviolet (UV-A) light in plants. Cryptochromes are probably the evolutionary descendents of DNA photolyases, which are light-activated DNA-repair enzymes, and are classified into three groups - plant cryptochromes, animal cryptochromes, and CRY-DASH proteins. Cryptochromes and photolyases have similar three-dimensional structures, characterized by an α/β domain and a helical domain. The structure also includes a chromophore, flavin adenine dinucleotide (FAD). The FAD-access cavity of the helical domain is the catalytic site of photolyases, and it is predicted also to be important in the mechanism of cryptochromes.
Similar content being viewed by others


Gene organization and evolutionary history
Cryptochromes are receptors for blue and ultraviolet (UV-A) light that share sequence similarity to DNA photolyases, DNA-repair enzymes that use blue light to repair UV-induced DNA damage by removing pyrimidine dimers from DNA [1]; cryptochromes have no photolyase activity, however [1–4]. There are two types of DNA photolyase, which repair different types of damage: CPD photolyases repair cyclobutane pyrimidine dimers (CPDs), and 6-4 photolyases repair 6-4 pyrimidine pyrimidone photoproducts. These photolyases together with the cryptochromes make up the photolyase/cryptochrome superfamily [5]. According to their sequence similarities, cryptochromes from a range of organisms can be clustered, more or less, into three subfamilies (Figure 1): plant cryptochromes, animal cryptochromes and cryptochrome-DASH proteins (CRY-DASH; see below).

An unrooted phylogenetic tree of the photolyase/cryptochrome superfamily, with subfamilies indicated on the right. Abbreviations: A, archaea; B, bacteria; F, fungi I; insects; P, plants; S, sponges; V, vertebrates.
Cryptochromes are widely distributed in bacteria and eukaryotes but are not found in archaea, although archaea do have a CPD photolyase (see Figure 1). The first cryptochrome gene to be identified was Arabidopsis CRY1 [6], and cryptochromes were soon found by homology in other plant species and in animals. Soon after the cloning of the first 6-4 photolyase from Drosophila [7, 8], a related sequence was discovered in the human expressed sequence tag (EST) databases that proved to encode human cryptochrome 1 (hCry1) [8, 9]. Cryptochromes have now been found in various animal lineages, including insects, fish, amphibians, and mammals. Animal cryptochromes act as components of the circadian clock that control daily physiological and behavioral rhythms and as photoreceptors that mediate entrainment of the circadian clock to light [3].
It was initially thought that only higher eukaryotes had cryptochromes and that prokaryotes had photolyases but not cryptochromes, but further searches of the more recently available genome databases revealed the presence of a cryptochrome gene in cyanobacteria (Synechocystis) [10]. This new type of cryptochrome was referred to as CRY-DASH, to underscore its relationship with cryptochromes found in Drosophila, Arabidopsis, Synechocystis, and Homo (although CRY-DASH itself is not found in Drosophila or humans) [11]. CRY-DASH proteins have been found not only in the photosynthetic cyanobacteria but also in non-photosynthetic bacteria, fungi, plants and animals, including Arabidopsis, Neurospora, zebrafish, and Xenopus (see Figure 1). The biological function of CRY-DASH proteins remains unknown at present.
The phylogenetic tree shown in Figure 1 illustrates the evolutionary relationships of photolyases and cryptochromes from different organisms. According to the tree topology, the photolyase/cryptochrome superfamily contains four subfamilies: animal cryptochromes/6-4 photolyase, plant cryptochromes, CRY-DASH proteins, and CPD photolyases. It is intriguing that animal cryptochromes and 6-4 photolyases are clustered together into the same clade, suggesting a close evolutionarily relationship between them even though they perform very different functions.
Characteristic structural features
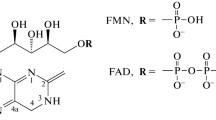
Cryptochromes show an overall structural similarity to DNA photolyases (Figure 2a, b), despite the fact that cryptochromes possess no photolyase activity. Most cryptochromes, with the exception of CRY-DASH proteins, are composed of two domains, an amino-terminal photolyase-related (PHR) region and a carboxy-terminal domain of varying size [4] (Figure 2c). The PHR region of cryptochromes appears to bind two chromophores, cofactors that absorb light; one chromophore is flavin adenine dinucleotide (FAD) and the other 5,10-methenyltetrahydrofolate (pterin or MTHF) [9, 12, 13]. Photolyases also have FAD, and the second chromophore can be either pterin or deazaflavin [9, 13]. The carboxy-terminal domain of cryptochromes is generally less conserved than the PHR region [2]; it is longer in most plant cryptochromes than animal cryptochromes, and CRY-DASH proteins lack this domain.

The structure of cryptochromes. A comparison of the structures of the photolyase-related (PHR) regions of (a) Arabidopsis CRY1 and (b) E. coli DNA photolyase. White lines indicate the boundaries of the FAD-access cavity; red and blue represent areas of negative and positive electrostatic potential, respectively. Reproduced with permission from [15]; copyright 2004 National Academy of Sciences USA. (c) A schematic representation of a typical photolyase/cryptochrome superfamily protein. The parts of the PHR region bound by pterin and FAD are indicated with brackets and the domains are shown below the protein. (d) The overall fold of a CRY-DASH protein (Synechosystis sp. PCC6803 cryptochrome).
Crystal structures have been determined for three members of the photolyase/cryptochrome superfamily in various species: CPD photolyase (Escherichia coli (Figure 2b), Synechococcus sp. and Thermus thermophilus) [14], CRY-DASH (Synechocystis sp.; Figure 2d) [11], and a plant cryptochrome (the PHR region of Arabidopsis CRY1; Figure 2a) [15]. The PHR regions have remarkably similar three-dimensional structures: all three fold into two domains, an α/β domain and a helical domain, which are connected by a variable loop that wraps around the α/β domain (Figure 2c, d). The α/β domain adopts a dinucleotide-binding fold with five-strand parallel β sheets flanked on either side by α helices. Two lobes of the helical domain form a cavity called the FAD-access cavity in all three proteins [11, 14, 15]. FAD is buried deeply inside the FAD-access cavity, but it may be accessible to the solvent from the bottom of the cavity. FAD binds non-covalently to the protein in a U-shaped conformation, with its adenine and isoalloxazine rings positioned at the bottom of the cavity (Figure 2d).
Despite the overall structure similarity, the Arabidopsis CRY1 PHR region has several structural characteristics that differ from photolyase and CRY-DASH. First, photolyase has a generally positively charged groove running through the FAD-access cavity (Figure 2b), which is where DNA interacts, and CRY-DASH has a similar positive electrostatic potential on the surface around the cavity, consistent with the observation that CRY-DASH binds DNA [11]. The CRY1 PHR region does not have this DNA-binding groove (Figure 2a), indicating that it may not bind DNA directly. Second, the surface of CRY1 PHR is predominantly negatively charged, except for a small concentration of positive charges near the FAD-access cavity [15]. Another distinction between the PHR regions of photolyase/CRY-DASH and CRY1 is that the FAD-access cavity of the latter is larger and deeper (see white line in Figure 2a). This difference is consistent with the fact that unlike photolyase, cryptochrome does not bind a pyrimidine dimer in the cavity. Finally, a single molecule of an ATP analog (AMP-PNP) can bind in the FAD-access cavity of the CRY1 PHR region [15], whereas neither photolyase nor CRY-DASH is reported to bind ATP.
Localization and function
Circadian biological clocks are biochemical oscillators that cycle about every 24 hours and that can be reset (entrained) by exposure to light and other environmental signals. In animals there is a central oscillator in the brain that controls the circadian behavior of the whole organism as well as peripheral oscillators in some tissues. The oscillation arises from a transcriptional feedback loop involving a set of clock transcription factors, including timeless (Tim), period (Per), clock (Clk) and Bmal1 as well as cryptochromes. Cryptochromes are ubiquitously expressed in the organs and tissues of all organisms, and they are generally nuclear proteins that regulate gene expression. The best-studied animal cryptochromes are Drosophila cryptochrome Cry and mouse cryptochromes Cry1 and Cry2 [16–20], and the two Arabidopsis cryptochromes CRY1 and CRY2 have also been extensively studied.
Drosophilacryptochromes
Drosophila Cry is a predominantly nuclear protein that mediates regulation of the circadian clock by light [16, 17], although it can also be found in the cytosol [21]. It regulates the circadian clock by interacting directly with the protein Tim to suppress the clock's negative feedback loop (Figure 3a). Light stimulates the Cry-Tim interaction, which promotes ubiquitination and proteosome-dependent degradation of Tim and represses formation of the Per-Tim heterodimer. Inhibition of a heterodimer of the Clock and Cycle proteins by the Per-Tim heterodimer is thus released and the phase of circadian oscillation is reset [21] (Figure 3a). Cryptochrome is apparently not the only photoreceptor that entrains the circadian clock in Drosophila, however. The behavioral rhythmicity of the crybmutant fly, which lacks Cry function, can nevertheless be entrained in response to light unless the signal transduction by the visual pigment is also eliminated [17]. In addition to its role as a photoreceptor for the entrainment of the central oscillator of Drosophila, Cry also has a light-independent role in the function of the peripheral circadian oscillator [22].

Regulation of the circadian clock by animal cryptochromes. (a) In Drosophila, Cry suppresses the negative feedback loop of the circadian clock by binding to Tim in a light-dependent manner; this results in the proteosome-dependent ubiquitin-mediated degradation of Tim (Ubq, ubiquitination) and thus to inhibition of the action of the Per-Tim heterodimer. Without Cry, the Per-Tim heterodimer would enter the nucleus and inhibit the binding of clock-cycle proteins (Per, Clk and Bmal1) to the E-box in the promoters of clock genes, preventing their expression. (b) In mammals, cryptochromes are integral parts of the negative feedback loop. The Cry protein interacts with Per to repress the activity of the transcription factors Clk and Bmal1 and thus to repress transcription. Cryptochromes may also be involved in the photo-entrainment of the mammalian circadian clock; clock genes are known to be regulated in response to neural signals from the retina in response to light, but it is not yet clear whether this involves cryptochromes.
Mammalian cryptochromes
The two functions of Drosophila Cry - as a photoreceptor for the entrainment of the circadian clock together with visual pigments and as an integral component of the circadian oscillator protein complex - are also features of the mammalian cryptochromes. Mammalian cryptochromes are predominantly nuclear proteins [18, 20], but they can also be found in the cytosol [23]. Like Drosophila Cry, mammalian cryptochromes perform both light-dependent and light-independent functions in the regulation of the circadian clock. Several observations demonstrate the light-dependent role of mammalian Cry proteins. Knockout mice lacking one or both Cry genes have a reduced or abolished ability to induce expression of genes such as per and the protooncogene c-fos in response to light [18, 24]. Moreover, the pupils of mutant mice lacking both Cry1 and Cry2 have reduced reflex responses to light [25].
On the other hand, the cry1 cry2 double mutant mouse shows an apparently normal rhythmicity in light-dark cycling conditions, but it loses rhythmicity instantaneously and completely in free-running (always dark) conditions [19]. These observations indicate that the Cry proteins play an essential and light-independent function in the mammalian central circadian oscillator, and that cryptochromes are not the only photoreceptors mediating light control of the clock. The fact that cryptochromes are integral parts of the mouse central oscillator makes it almost impossible to test directly their role in the light entrainment of the clock. Nevertheless, it has been found that, somewhat analogous to the situation for Drosophila, the mouse cry mutant retains its ability to mediate light input unless the function of visual pigments is also disrupted at the same time. Triple-mutant mice carrying mutations of both cryptochromes together with a retinal-degenerative mutation are nearly arrhythmic under light-dark cycling conditions [24]. These results demonstrate that mammalian Cry proteins are indeed involved in regulation of the circadian clock by light, but their role in the light entrainment of the circadian clock is carried out redundantly by other photoreceptors. It seems clear now that the additional photoreceptors acting together with cryptochromes for the entrainment of the mammalian circadian oscillator are visual rod-cone opsins and the related protein melanopsin [26, 27].
Like Drosophila Cry, mammalian cryptochromes interact physically with clock proteins, including the promoter-binding transcription regulators Per, Clk and Bmal1 (Figure 3b). In contrast to Drosophila Cry, mammalian Cry proteins are components of the negative-feedback loop of the circadian clock (Figure 3b). The physical interaction of cryptochrome with other clock components affects their activity, interaction, degradation, or nuclear trafficking, and consequently alters the transcriptional regulation of the clock genes [28, 29]. But the interaction between cryptochromes and other clock proteins such as Per, Clk and Bmal1 appears not to be affected by light, suggesting that such interactions may not be the mechanism of photo-entrainment of the circadian clock, as they are in Drosophila. In addition to the direct regulation of transcription via physical interaction with promoter-binding transcription regulators, cryptochromes may also affect the circadian clock by participating in the regulation of histone modifications [30, 31], but how this works remains to be elucidated.
Arabidopsiscryptochromes
Arabidopsis CRY1 and CRY2 are predominantly nuclear proteins that mediate regulation of gene expression and entrainment of the circadian clock in response to light [32–35]. CRY1 and CRY2 play major roles in plant photomorphogenesis, such as inhibition of stem elongation by blue light, stimulation of leaf expansion by blue light, and regulation of floral initiation by day length [6, 36–38]. It appears that cryptochromes control developmental changes in plants via changes of gene expression in response to light. CRY1 and CRY2 together are responsible for blue-light-dependent changes in gene expression of up to 10-20% of the Arabidopsis genome [39].
There are at least two mechanisms by which cryptochromes may affect nuclear gene-expression changes in response to light. First, a cryptochrome molecule may interact with proteins associated with transcriptional machinery to affect transcription directly. Arabidopsis CRY2 binds to chromatin in a DNA sequence-independent manner ([40] and M. Maymon and C.L., unpublished observations), but it is unclear how a sequence-independent chromatin-interacting protein may regulate gene expression. Unlike the animal cryptochromes that have been shown to regulate transcription via physical interactions with promoter-binding transcription regulators, no such interaction has been reported for plant cryptochromes. An alternative model is that plant cryptochromes may interact with proteins exerting other cellular functions to regulate the stability, modification, cellular trafficking of the transcriptional regulators. For example, plant cryptochromes have been found to interact with an E3 ubiquitin ligase, COP1, suggesting that plant cryptochromes may act in the way not yet discovered for the animal cryptochromes [32, 41, 42]. Consistent with this view, it has also been found recently that Arabidopsis cryptochromes mediate suppression by blue light of the proteasome-dependent degradation of an important floral regulator, CONSTANS [43]. Exactly how cryptochromes do this needs to be investigated further.
Mechanism
The catalytic mechanism of cryptochromes has not been fully elucidated, but some clues can be found in the mechanism of CPD photolyases, where FAD plays the main catalytic role [14]. In a DNA-repair reaction, CPD photolyase binds to the pyrimidine dimer of DNA and 'flips' it out from within the DNA duplex into the FAD-access cavity of the enzyme, to form a stable complex. The other chromophore (pterin or deazaflavin), which is also called the 'antenna' chromophore, absorbs photons of blue or UV-A light, and it transfers the excitation energy to the flavin of FAD. Flavin in the excited state donates an electron to the pyrimidine dimer to split the cyclobutane ring. The electron is transferred back to flavin in this process, resulting in regeneration of groundstate flavin. The repaired dinucleotide no longer fits into the FAD-access cavity, so it disassociates from the photolyase. The exact role of FAD and the FAD-access cavity in the function of cryptochromes remains unclear, but it is conceivable that it may also be involved in electron-transfer reactions.
Although the PHR region that contains the chromophore(s) is the most conserved part of the proteins, the carboxy-terminal domain has been shown to have a role in the function or regulation of both animal and plant cryptochromes. Expression of the carboxy-terminal domains of Arabidopsis cryptochromes fused to the marker enzyme b-glucuronidase confers a constitutive growth response to light even in darkness in the absence of the PHR region [32, 41]. In contrast, the PHR regions of the Drosophila and Xenopus cryptochromes are physiologically active in the absence of the carboxy-terminal domain [44, 45]. The carboxy-terminal domain of Drosophila Cry is important for protein stability, interaction with Tim, and sensitivity of the photoreceptor to circadian light signals [44], whereas the carboxy-terminal domain of Xenopus Cry is required for its nuclear localization [45].
Cryptochromes are regulated by phosphorylation. It has been shown that Arabidopsis cryptochromes are phosphorylated in response to blue light and that this is associated with the function and regulation of the photoreceptors [46, 47]. Moreover, when Arabidopsis CRY1 was expressed in insect cells, it was found to undergo ATP-dependent and blue-light-dependent autophosphorylation [15, 47, 48]. It is not known whether animal cryptochromes also bind to ATP, although it has been shown that mouse cryptochromes are phosphorylated [49].
The interaction between the Arabidopsis CRY1 PHR region and ATP has a few interesting features reminiscent of the interaction between pyrimidine dimer and photolyase [14]: the phosphate groups of ATP are exposed to solvent; the adenine and ribose moieties are buried deep within the FAD-access cavity; and ATP can have a water-mediated contact with FAD [15]. The interaction of the Arabidopsis CRY1 pHR region with ATP also lacks several features commonly found in protein-ATP interactions, such as protein-to-phosphate interaction, protein-to-Mg2+ contact, and a nearby serine residue for phosphotransfer [15]. An examination of the topology of the CRY1 PHR region structure shows, however, that all these features could potentially be provided by the carboxy-terminal domain of the cryptochrome (Figure 4). The observation that the serine-rich carboxy-terminal domains of Arabidopsis cryptochromes fused to β-glucuronidase are constitutively phosphorylated in vivo ([32, 46], suggests that a phosphotransfer may occur from ATP bound to the FAD-access cavity to the nearby carboxy-terminal domain (Figure 4a). It is also conceivable that photon-excited FAD may trigger electron transfer to the nucleotide and phosphotransfer from ATP to serine residues on the carboxy-terminal domain. Because the surface of the PHR region is predominantly negatively charged, especially in the place where the carboxy-terminal domain is likely to interact with it, the phosphorylated carboxy-terminal domain would then be repelled from the PHR region surface, resulting in a change of cryptochrome conformation. This conformational change would allow it to interact with other signaling proteins and to propagate the light signal (Figure 4a). Alternatively, another molecule of cryptochrome binding to the FAD-access cavity may also provide the missing features needed for a productive ATP-cryptochrome interaction. Indeed, both CRY2-CRY2 interaction and CRY1-CRY2 interactions can be detected in Arabidopsis (D. Shalitin, X. Yu, and C.L., unpublished observations). Formation of either a homo-oligomer or a hetero-oligomer of cryptochromes would provide a mechanism for intermolecular phosphotransfer, which may change the structure of the cryptochromes (Figure 4b, c).

Possible models of the phosphorylation-dependent structural changes of plant cryptochromes in response to blue light. The PHR region is predominantly negatively charged (-), and the carboxy-terminal domain (C) can be made negatively charged by phosphorylation (which requires ATP and releases inorganic phosphate, Pi). In all models, phosphorylation leads to binding of unknown signaling partners (X, Y, Z) and to regulation of plant development. (a) One model is that phosphorylation of the carboxy-terminal domain in response to light is performed by ATP bound to the PHR region; this leads to dissociation of the two domains. (b) A second possibility is that phosphotransfer in response to light involves the interaction of two cryptochromes encoded by the same gene. (c) Alternatively, intermolecular phosphotransfer could involve the interaction of different cryptochromes. All three scenarios may exist in plant cells, and the activity of a cryptochrome may be determined by the kinetics of the different reactions.
Frontiers
Despite recent progress in the study of cryptochromes, there are still many intriguing questions about their structure and mechanism. For example, what is the role of the FAD-access cavity in cryptochrome? Can a photon-excited FAD trigger a phosphotransfer from ATP to the carboxy-terminal domain? Elucidation of the structure of holocryptochromes including the carboxy-terminal domain may help us to answer these questions. For mammalian cryptochromes, it is unclear whether the interaction of Cry with other clock proteins is affected by light in vivo, and also how may cryptochromes mediate regulation of the circadian clock by light. The physiological function of CRY-DASH proteins is also not clear, although they may regulate gene expression directly. Elucidation of the function and mechanism of CRY-DASH would provide additional clues about the evolutionary history of the cryptochromes.
References
Sancar A: Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors. Chem Rev. 2003, 103: 2203-2237. 10.1021/cr0204348. A comprehensive review of the biochemical characteristics of photolyases and cryptochromes.
Lin C, Shalitin D: Cryptochrome structure and signal transduction. Annu Rev Plant Biol. 2003, 54: 469-496. 10.1146/annurev.arplant.54.110901.160901. A review of the structure and function of plant cryptochromes.
Cashmore AR: Cryptochromes: enabling plants and animals to determine circadian time. Cell. 2003, 114: 537-543. 10.1016/j.cell.2003.08.004. A review of the role of cryptochromes in the regulation of the circadian clock.
Todo T: Functional diversity of the DNA photolyase/blue light receptor family. Mutat Res. 1999, 434: 89-97. This review discusses the evolutionary relationship between DNA photolyase and cryptochromes.
Kanai S, Kikuno R, Toh H, Ryo H, Todo T: Molecular evolution of the photolyase-blue-light photoreceptor family. J Mol Evol. 1997, 45: 535-548. An analysis of the evolutionary history of DNA photolyases and cryptochromes.
Ahmad M, Cashmore AR: HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor. Nature. 1993, 366: 162-166. 10.1038/366162a0. The isolation of the first cryptochrome gene.
Todo T, Takemori H, Ryo H, Ihara M, Matsunaga T, Nikaido O, Sato K, Nomura T: A new photoreactivating enzyme that specifically repairs ultraviolet light-induced (6-4) phoptoproducts. Nature. 1993, 361: 371-374. 10.1038/361371a0. The first isolation and purification of 6-4 photolyase from Drosophila.
Todo T, Ryo H, Yamamoto K, Toh H, Inui T, Ayaki H, Nomura T, Ikenaga M: Similarity among the Drosophila (6-4)photolyase, a human photolyase homolog, and the DNA photolyase-blue-light photoreceptor family. Science. 1996, 272: 109-112. The isolation of the first cryptochrome gene from an animal.
Hsu DS, Zhao X, Zhao S, Kazantsev A, Wang RP, Todo T, Wei YF, Sancar A: Putative human blue-light photoreceptors hCRY1 and hCRY2 are flavoproteins. Biochemistry. 1996, 35: 13871-13877. 10.1021/bi962209o. The first description of the human cryptochromes, hCry1 and hCry2.
Hitomi K, Okamoto K, Daiyasu H, Miyashita H, Iwai S, Toh H, Ishiura M, Todo T: Bacterial cryptochrome and photolyase: characterization of two photolyase-like genes of Synechocystis sp. PCC6803. Nucleic Acids Res. 2000, 28: 2353-2362. 10.1093/nar/28.12.2353. The first report of the bacterial cryptochrome CRY-DASH.
Brudler R, Hitomi K, Daiyasu H, Toh H, Kucho K, Ishiura M, Kanehisa M, Roberts VA, Todo T, Tainer JA, et al: Identification of a new cryptochrome class. Structure, function, and evolution. Mol Cell. 2003, 11: 59-67. 10.1016/S1097-2765(03)00008-X. The crystal structure of bacterial CRY-DASH.
Lin C, Robertson DE, Ahmad M, Raibekas AA, Jorns MS, Dutton PL, Cashmore AR: Association of flavin adenine dinucleotide with the Arabidopsis blue light receptor CRY1. Science. 1995, 269: 968-970. The first report of noncovalent binding of FAD to Arabidopsis CRY1.
Malhotra K, Kim ST, Batschauer A, Dawut L, Sancar A: Putative blue-light photoreceptors from Arabidopsis thaliana and Sinapis alba with a high degree of sequence homology to DNA photolyase contain the two photolyase cofactors but lack DNA repair activity. Biochemistry. 1995, 34: 6892-6899. 10.1021/bi00020a037. The first report of a second chromophore for cryptochromes.
Park HW, Kim ST, Sancar A, Deisenhofer J: Crystal structure of DNA photolyase from Escherichia coli. Science. 1995, 268: 1866-1872. The first crystal structure of a DNA photolyase.
Brautigam CA, Smith BS, Ma Z, Palnitkar M, Tomchick DR, Machius M, Deisenhofer J: Structure of the photolyase-like domain of cryptochrome 1 from Arabidopsis thaliana. Proc Natl Acad Sci USA. 2004, 101: 12142-12147. 10.1073/pnas.0404851101. The crystal structure of the PHR region of an Arabidopsis cryptochrome, which was the first plant cryptochrome structure to be solved.
Emery P, So WV, Kaneko M, Hall JC, Rosbash M: CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell. 1998, 95: 669-679. 10.1016/S0092-8674(00)81637-2. The circadian photosensitivity of a Drosophila strain overexpressing Cry is shown to be increased.
Stanewsky R, Kaneko M, Emery P, Beretta B, Wager-Smith K, Kay SA, Rosbash M, Hall JC: The cryb mutation identifies cryptochrome as a circadian photoreceptor in Drosophila. Cell. 1998, 95: 681-692. 10.1016/S0092-8674(00)81638-4. The authors show that a Drosophila cry mutant, cryb, exhibits poor synchronization to light-dark cycles and shows no response to brief light pulses.
Thresher RJ, Vitaterna MH, Miyamoto Y, Kazantsev A, Hsu DS, Petit C, Selby CP, Dawut L, Smithies O, Takahashi JS, et al: Role of mouse cryptochrome blue-light photoreceptor in circadian photoresponses. Science. 1998, 282: 1490-1494. 10.1126/science.282.5393.1490. The first report of a phenotype for the cry2 knockout mouse. The mutant has a intrinsic circadian period that is only an hour long.
van der Horst GT, Muijtjens M, Kobayashi K, Takano R, Kanno S, Takao M, de Wit J, Verkerk A, Eker AP, van Leenen D, et al: Mammalian Cry1 and Cry2 are essential for maintenance of circadian rhythms. Nature. 1999, 398: 627-630. 10.1038/19323. The first report of phenotype of cry1 and cry1/cry2 double knockout mutants in mouse. The mice lacking cry1 protein display accelerated free-running periodicity of locomotor activity, but mice lacking both cry proteins show complete loss of free-running periodicity.
Kume K, Zylka MJ, Sriram S, Shearman LP, Weaver DR, Jin X, Maywood ES, Hastings MH, Reppert SM: mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell. 1999, 98: 193-205. 10.1016/S0092-8674(00)81014-4. The authors show that Cry1 and Cry2 inhibit transcription mediated by the CLOCK and BMAL1 proteins in mammalian cultured cell line.
Ceriani MF, Darlington TK, Staknis D, Mas P, Petti AA, Weitz CJ, Kay SA: Light-dependent sequestration of Timeless by Cryptochrome. Science. 1999, 285: 553-556. 10.1126/science.285.5427.553. Drosophila Cry blocks the function of Per:Tim heterodimer complex in a light-dependent fashion.
Krishnan B, Levine JD, Lynch MK, Dowse HB, Funes P, Hall JC, Hardin PE, Dryer SE: A new role for cryptochrome in a Drosophila circadian oscillator. Nature. 2001, 411: 313-317. 10.1038/35077094. A Drosophila cry mutant renders the peripheral circadian oscillator largely arrhythmic but leaves the central circadian oscillator intact in pacemaker neurons.
Thompson CL, Rickman CB, Shaw SJ, Ebright JN, Kelly U, Sancar A, Rickman DW: Expression of the blue-light receptor cryptochrome in the human retina. Invest Ophthalmol Vis Sci. 2003, 44: 4515-4521. 10.1167/iovs.03-0303. The authors show that Cry2 is localized throughout the cytoplasm as well as the nucleus in the ganglion-cell layer of human retina.
Selby CP, Thompson C, Schmitz TM, Van Gelder RN, Sancar A: Functional redundancy of cryptochromes and classical photoreceptors for nonvisual ocular photoreception in mice. Proc Natl Acad Sci USA. 2000, 97: 14697-14702. 10.1073/pnas.260498597. In a triple mutant mouse lacking Cry1, Cry2 and rhodopsin, the light induction of c-fos transcription in the suprachiasmatic nuclei is markedly reduced, suggesting redundant function of cryptochromes and visual pigments in the photo-entrainment of the circadian clock in the mouse.
Van Gelder RN, Wee R, Lee JA, Tu DC: Reduced pupillary light responses in mice lacking cryptochromes. Science. 2003, 299: 222-10.1126/science.1079536. This report showed a defect of the cry mutant in the pupillary light response, suggesting a role for cryptochromes as photoreceptors in the mouse.
Hattar S, Lucas RJ, Mrosovsky N, Thompson S, Douglas RH, Hankins MW, Lem J, Biel M, Hofmann F, Foster RG, et al: Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature. 2003, 424: 76-81. 10.1038/nature01761. Mice lacking melanopsin coupled with disabled rod and cone phototransduction mechanisms fail to show any circadian phase-shifting and other nonvisual photoreception, demonstrating the role of melanopsin as a photoreceptor for the circadian clock.
Panda S, Sato TK, Castrucci AM, Rollag MD, DeGrip WJ, Hogenesch JB, Provencio I, Kay SA: Melanopsin (Opn4) requirement for normal light-induced circadian phase shifting. Science. 2002, 298: 2213-2216. 10.1126/science.1076848. The authors show that mice with both degeneration of the outer retina and a deficiency in melanopsin exhibit complete loss of photoentrainment of the circadian oscillator and of other nonvisual photoreception.
Griffin EA, Staknis D, Weitz CJ: Light-independent role of CRY1 and CRY2 in the mammalian circadian clock. Science. 1999, 286: 768-771. 10.1126/science.286.5440.768. The authors show that mammalian cryptochromes act as light-independent inhibitors of the CLOCK:BMAL1 heterodimer.
Lee C, Etchegaray JP, Cagampang FR, Loudon AS, Reppert SM: Post-translational mechanisms regulate the mammalian circadian clock. Cell. 2001, 107: 855-867. 10.1016/S0092-8674(01)00610-9. This report shows that mammalian Cry proteins are necessary for stabilizing phosphorylated Per2 and for the nuclear accumulation of Per1, Per2 and CKI3.
Etchegaray JP, Lee C, Wade PA, Reppert SM: Rhythmic histone acetylation underlies transcription in the mammalian circadian clock. Nature. 2003, 421: 177-182. 10.1038/nature01314. Acetylation of histone H3 is a potential target of the inhibitory action of Cry.
Naruse Y, Oh-hashi K, Iijima N, Naruse M, Yoshioka H, Tanaka M: Circadian and light-induced transcription of clock gene Per1 depends on histone acetylation and deacetylation. Mol Cell Biol. 2004, 24: 6278-6287. 10.1128/MCB.24.14.6278-6287.2004. The authors show that mammalian Cry1 represses transcription through histone deacetylases and the protein mSin3B that is associated with them, indicating a role for histone modification in the circadian clock regulation.
Yang H-Q, Wu Y-J, Tang R-H, Liu D, Liu Y, Cashmore AR: The C termini of Arabidopsis cryptochromes mediate a constitutive light response. Cell. 2000, 103: 815-827. 10.1016/S0092-8674(00)00184-7. The authors show that the carboxy-terminal domain of Arabidopsis CRY1 and CRY2 is responsible for the signaling function of the proteins in response to light.
Guo H, Duong H, Ma N, Lin C: The Arabidopsis blue light receptor cryptochrome 2 is a nuclear protein regulated by a blue light-dependent post-transcriptional mechanism. Plant J. 1999, 19: 279-287. 10.1046/j.1365-313X.1999.00525.x. This paper and [34] show that the carboxy-terminal domain of Arabidopsis CRY2 has a functional nuclear-localization signal.
Kleiner O, Kircher S, Harter K, Batschauer A: Nuclear localization of the Arabidopsis blue light receptor cryptochrome 2. Plant J. 1999, 19: 289-296. 10.1046/j.1365-313X.1999.00535.x. See [33].
Somers DE, Devlin PF, Kay SA: Phytochromes and cryptochromes in the entrainment of the Arabidopsis circadian clock. Science. 1998, 282: 1488-1490. 10.1126/science.282.5393.1488. This report shows that Arabidopsis CRY1, but not CRY2, is the blue light photoreceptor for circadian period length control.
Guo H, Yang H, Mockler TC, Lin C: Regulation of flowering time by Arabidopsis photoreceptors. Science. 1998, 279: 1360-1363. 10.1126/science.279.5355.1360. This report shows the function of Arabidopsis CRY2 in the control of photoperiodic flowering.
Mockler T, Yang H, Yu X, Parikh D, Cheng YC, Dolan S, Lin C: Regulation of photoperiodic flowering by Arabidopsis photoreceptors. Proc Natl Acad Sci USA. 2003, 100: 2140-2145. 10.1073/pnas.0437826100. This report shows the redundant as well as the antagonistic function of cryptochromes and phytochromes in the control of photoperiodic flowering.
El-Assal SE-D, Alonso-Blanco C, Peeters AJ, Raz V, Koornneef M: A QTL for flowering time in Arabidopsis reveals a novel allele of CRY2. Nat Genet. 2001, 29: 435-440. 10.1038/ng767. A QTL identified from natural populations is CRY2; a missense defect in the CRY2 gene causes photoperiod-insensitive flowering in an Arabidopsis accession found near the equator.
Ma L, Li J, Qu L, Hager J, Chen Z, Zhao H, Deng XW: Light control of Arabidopsis development entails coordinated regulation of genome expression and cellular pathways. Plant Cell. 2001, 13: 2589-2607. 10.1105/tpc.13.12.2589. This study investigates genome-wide mRNA expression in Arabidopsis in response to light quality, showing that cryptochromes are the major photoreceptors mediating regulation of gene expression by blue light.
Cutler SR, Ehrhardt DW, Griffitts JS, Somerville CR: Random GFP::cDNA fusions enable visualization of subcellular structures in cells of Arabidopsis at a high frequency. Proc Natl Acad Sci USA. 2000, 97: 3718-3723. 10.1073/pnas.97.7.3718. A report that the Arabidopsis CRY2 carboxy-terminal domain can be associated with chromosomes.
Wang H, Ma LG, Li JM, Zhao HY, Deng XW: Direct interaction of Arabidopsis cryptochromes with COP1 in light control development. Science. 2001, 294: 154-158. 10.1126/science.1063630. This report shows that Arabidopsis cryptochromes interact with the E3 ubiquitin ligase COP1.
Sullivan JA, Deng XW: From seed to seed: the role of photoreceptors in Arabidopsis development. Dev Biol. 2003, 260: 289-297. 10.1016/S0012-1606(03)00212-4. A review of the regulation of plant development by light and the role of COP proteins in this process.
Valverde F, Mouradov A, Soppe W, Ravenscroft D, Samach A, Coupland G: Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science. 2004, 303: 1003-1006. 10.1126/science.1091761. This paper reports the ubiquitination of CRY2 and degradation of a zinc-finger protein by the proteasome in blue light.
Busza A, Emery-Le M, Rosbash M, Emery P: Roles of the two Drosophila cryptochrome structural domains in circadian photoreception. Science. 2004, 304: 1503-1506. 10.1126/science.1096973. The PHR region of Drosophila Cry is active alone in vivo.
Zhu H, Conte F, Green CB: Nuclear localization and transcriptional repression are confined to separable domains in the circadian protein cryptochrome. Curr Biol. 2003, 13: 1653-1658. 10.1016/j.cub.2003.08.033. This paper reports that the carboxy-terminal domain of animal cryptochromes directs nuclear localization.
Shalitin D, Yang H, Mockler TC, Maymon M, Guo H, Whitelam GC, Lin C: Regulation of Arabidopsis cryptochrome 2 by blue-light-dependent phosphorylation. Nature. 2002, 417: 763-767. 10.1038/nature00815. A report of the blue-light-specific phosphorylation of Arabidopsis CRY2.
Shalitin D, Yu X, Maymon M, Mockler T, Lin C: Blue light-dependent in vivo and in vitro phosphorylation of Arabidopsis cryptochrome 1. Plant Cell. 2003, 15: 2421-2429. 10.1105/tpc.013011. A report of a blue-light-dependent phosphorylation of Arabidopsis CRY1 and its autophosphorylation activity in vitro.
Bouly JP, Giovani B, Djamei A, Mueller M, Zeugner A, Dudkin EA, Batschauer A, Ahmad M: Novel ATP-binding and autophosphorylation activity associated with Arabidopsis and human cryptochrome-1. Eur J Biochem. 2003, 270: 2921-2928. 10.1046/j.1432-1033.2003.03691.x. Characterization of the autophosphorylation activity of Arabidopsis CRY1 in vitro.
Eide EJ, Vielhaber EL, Hinz WA, Virshup DM: The circadian regulatory proteins BMAL1 and cryptochromes are substrates of casein kinase Iepsilon. J Biol Chem. 2002, 277: 17248-17254. 10.1074/jbc.M111466200. The authors show that mammalian Cry1 is phosphorylated by CKIε.
Acknowledgements
We thank the previous and current members of our laboratories for discussions and preparation of Figures. Figures 1 and 2d were prepared with the help of H. Daiyasu (Kyoto University) and K. Hitomi (The Scripps Research Institute), respectively; Figure 4 was prepared with the help of John Klejnot. We also thank Johann Deisenhofer and The National Academy of Sciences USA for the use of their published material in Figure 2a, b, and Johann Deisenhofer and A. Sancar for helpful discussions. Research in the authors' laboratories is partially supported by NIH (GM56265 to C.L.), NSF (MCB-0091391 to C.L.), and by Grant-in-Aid for Scientific Research from the Japanese Ministry of Education, Cultures, Sports, Science and Technology of Japan (to T.T.).
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
About this article
Cite this article
Lin, C., Todo, T. The cryptochromes. Genome Biol 6, 220 (2005). https://doi.org/10.1186/gb-2005-6-5-220
Published:
DOI: https://doi.org/10.1186/gb-2005-6-5-220