
Abstract
Introduction
Acetaminophen (APAP) toxicity is the most common cause of acute liver failure in the US and Europe. Massive hepatocyte necrosis is the predominant feature of APAP-induced acute liver injury (ALI). Liver regeneration is a vital process for survival after a toxic insult, it occurs at a relative late time point after the injurious phase. Currently, N-acetylcysteine (NAC), a glutathione precursor, is the antidote for acetaminophen overdose. However, NAC is effective only for patients who present within hours of an acute overdose, and is less effective for late-presenting patients. It is possible that in delayed patients, previously reduced endogenous glutathione (GSH) level has restored and prolonged treatment with NAC might be toxic and impair liver regeneration. Therefore, we hypothesize that prolonged treatment with NAC impairs liver regeneration in ALI induced by APAP.
Methods
ALI was induced in C57BL/6 male mice by a single dose of APAP (350 mg/kg) by intraperitoneal injection. After two hours of APAP challenge, the mice were given 100 mg/kg NAC dissolved in 0.6 mL saline, or saline treatment every 12 hours for a total of 72 hours.
Results
Seventy-two hours after APAP challenge, compared with saline treatment, NAC treatment significantly increased serum transaminases (alanine transaminase/aspartate aminotransferase), induced evident hepatocyte vacuolation in the periportal area and delayed liver regeneration seen in histopathology. This detrimental effect was associated with reduced hepatic nuclear factor (NF)-κB DNA binding and decreased expression of cell cycle protein cyclin D1, two important factors in liver regeneration.
Conclusions
Prolonged treatment with NAC impairs liver regeneration in ALI induced by APAP.
Similar content being viewed by others

Introduction
Acetaminophen (APAP) toxicity is the most common cause of acute liver failure in the US and Europe [1]; however, the underlying mechanisms of APAP-induced hepatotoxicity are still not completely understood. The toxic response to APAP is triggered by a highly reactive metabolite, N-acetyl-p-benzoquinone imine (NAPQI), which reacts with and depletes glutathione (GSH), after which it forms covalent adducts and initiates mitochondrial oxidative stress [2, 3]. This increases the membrane permeability transition and causes the collapse of the mitochondrial membrane potential, which results in a diminished mitochondrial capacity to synthesize ATP [4], and ATP depletion leads to cell death. Massive necrosis of the hepatocyte is a characteristic feature of APAP-induced acute lung injury (ALI) [5].
Liver regeneration is a vital process for survival after a toxic insult [6, 7]. Regeneration ensures the replacement of necrotic cells and the full recovery of organ function. As hepatocytes are mostly in a quiescent state (G0), the regeneration process requires entry into the highly regulated cell cycle [8]. The induction of cyclin D1 is the most reliable marker for cell cycle (G1 phase) progression in hepatocytes. Once hepatocytes express cyclin D1, they have passed the G1 restriction point and are committed to DNA replication [8].
Many factors can influence liver regeneration. Currently, nuclear factor (NF)-κB is thought to play a major role in the initiation of liver regeneration after cell or tissue loss (such as by hepatotectomy) [8]. In addition, nutrients and metabolic status can also influence regeneration, because APAP induces massive hepatocyte necrosis. After the loss of a large number of parenchymal cells, the metabolic work of surviving hepatocytes is increased and more ATP is needed for maintaining homeostasis and regeneration [8].
Currently, N-acetylcysteine (NAC), a GSH precursor, is the antidote for APAP overdose [9]. However, this antidotal therapy is effective only for patients who present within hours of an acute overdose, and is less effective for late-presenting patients [9, 10]. There are two possibilities: delayed treatment with NAC is not effective when massive hepatonecrosis has occurred; at a late hour, liver regeneration becomes evident and plays a crucial role in liver recovery; however, delayed NAC treatment does not maintain the proliferation of primary hepatocytes [11]. In addition, there is evidence showing that reduced endogenous GSH concentration gradually gets back to a normal level at the late time point [12], after which continuous treatment with NAC may no longer replenish endogenous GSH. Instead, it might interfere with glucose metabolism, because NAC is involved in glucose and mitochondrial tricarboxylic acid (TCA) metabolisms [13]; and a high dose of NAC (600 to 1200 mg/kg) impairs liver glucose metabolism [13]. The liver is responsible for the metabolism of carbohydrate, lipid and protein; these processes are all interlinked [14], therefore, prolonged treatment with a high dose of NAC may interfere with hepatic normal metabolic functions and impairs liver recovery from APAP hepatotoxicity.
At present, NAC is also used to treat non-acetaminophen-induced hepatotoxicity [15, 16]. However, there are limited data available on the efficacy and safety of NAC. The doses vary from 100 mg/kg/20 hours to 300 mg/kg/24 hours in patients; the median duration of NAC administration in children with non-APAP-induced ALI is five days (range = 1 to 77 days) [16]. In an amatoxin (a toxic peptide from poisoning mushroom) induced hepatotoxicity model, NAC was administered at a dosage of 1200 mg/kg and repeated every four hours at a dosage of 600 mg/kg for a total of 48 hours. The NAC-treated mice have higher serum alanine transaminase (ALT)/aspartate aminotransferase (AST) than the amatoxin-challenged mice treated with saline [15], this indicates that prolonged treatment with a high dose of NAC might be toxic.
Based on this information, we hypothesized that NAC might impair liver regeneration in ALI induced by APAP. To evaluate this idea, ALI was induced in mice by APAP intraperitoneal injection, and the mice were observed over a 72-hour period. Compared with saline treatment, prolonged treatment with NAC increased serum ALT/AST and also induced evident hepatocyte vacuolation and delayed liver regeneration as shown by histopathology; This detrimental effect was associated with reduced hepatic NF-κB DNA binding and decreased expression of cell cycle protein cyclin D1, two important factors in liver regeneration. In conclusion, prolonged treatment with therapeutic dose of NAC impairs liver regeneration in ALI induced by APAP, reevaluation of optimal doses and duration of NAC therapy is needed.
Materials and methods
Materials
All chemicals were purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA) unless otherwise noted.
Animal model and experimental groups
This research protocol complied with the regulations regarding the care and use of experimental animals published by the National Institutes of Health and was approved by the Institutional Animal Use and Care Committee of the University of Pittsburgh Medical School. Male C57BL/6 mice weighing 20 to 25 g (Jackson Laboratories, Bar Harbor, ME, USA) were used in this study. The animals were maintained at the University of Pittsburgh Animal Research Center with a 12-hour light-dark cycle and free access to standard laboratory food and water. The animals were fasted over night prior to the experiments.
Experiment A
ALI was induced by a single dose of APAP (350 mg/kg dissolved in 1 mL sterile saline) administered by intraperitoneal injection. APAP-challenged mice were then randomized into the NAC (n = 7) group or the saline group (n = 7). Six mice injected with saline not containing APAP served as a control group. Two hours after APAP administration, each group was given the following treatments every 12 hours for a total of 72 hours: 100 mg/kg NAC dissolved in 0.6 mL sterile saline for the NAC group, and 0.6 mL saline for the saline group and the control group. Seventy-two hours after APAP injection, all surviving mice in each group were anaesthetized with sodium pentobarbital (90 mg/kg intraperitoneally), and the following procedures were performed: blood was aspirated from the heart for the subsequent measurements of ALT and AST; the left lobe of the liver was harvested for pathology (H&E staining); and the right lobe of the liver was harvested and frozen for measurement of hepatic NF-κB DNA binding by electrophoretic mobility shift assays (EMSA) and hepatic tissue cyclin D1 expression by western blot.
Experiment B
Three separate groups of mice were treated as described above with the exception that the treatment period was only 24 hours (n = 6 for each group).
Plasma aminotransferase measurements
Plasma levels of AST and ALT were measured at 37°C with a commercially available kit (Sigma Diagnostic, St Louis, MO, USA).
Histological analysis
Consecutive sections (5 μm) of paraffin-embedded liver were prepared for H&E staining. The percentage of necrosis was estimated by evaluating the number of microscopic fields with necrosis compared with the entire cross-section. In general, necrosis was estimated at low power (×100) and questionable areas were evaluated at higher magnification (×200 or ×400). The pathologist (XH) evaluated all histological sections in a blinded fashion. Inflammatory cell infiltration results were scored semi-quantitatively by averaging the number of inflammatory cells per microscopic field at a magnification of 200×. Five fields were evaluated per tissue sample, and six animals in each group were examined.
Tissue myeloperoxidase
Neutrophil infiltration was measured at 72 hours by determining myeloperoxidase (MPO) activity in liver tissue homogenates and was used as an index of neutrophil infiltration in all groups. At the time of sample determination, liver tissue samples (400 mg) were harvested and snap frozen immediately in liquid nitrogen and stored at -70°C until analysis was performed. Samples were homogenized in suspension buffer (50 mmol/L potassium phosphate/0.5% hexadecyltrimethylammonium bromide; pH 6.0), sonicated on ice, freeze-thawed twice, resonicated and then centrifuged for 15 minutes as 20,000 g at 4°C. Liver supernatants were heated for two hours at 60°C, and recentrifuged for 15 minutes at 20,000 g at 4°C.
To determine the MPO activity, 100 μL of the supernatants was incubated for three minutes at 37°C in a reaction solution containing 40% PBS, 8% N,N-dimethylformamide, 1.6 mmol/L 3,3', 5,5'-tetramethylbenzidine, 0.3 mmol/L hydrogen peroxide and 80 mmol/L sodium phosphate (pH 5.4). The reaction was stopped by adding ice-cold 800 mmol/L acetic acid solution (pH 3.0) and by placing the assay tubes on ice. Samples were read at 655 nm and total MPO activity calculated as the change in absorbency per minute per gram of tissue multiplied by the dilution factor. The levels were expressed as units per gram of tissue (U/g) [17].
EMSA
NF-κB activation was determined by EMSA as previously described [18]. The gels were dried and exposed to Biomax film (Kodak, Rochester, NY, USA) at -70°C overnight with use of an intensifying screen.
Western blot
Liver protein was extracted as previously described [19]. Equivalent amounts of protein were boiled in sample buffer and separated on 7.5% pre-cast SDS-PAGE (Bio-Rad, Hercules, CA, USA) and transferred to nylon membranes. Membranes were then probed with a specific antibody against cyclin D1 (Cell signaling Technology, Lexington, KY, USA) protein, visualized with an Enhanced Chemiluminescence substrate (ECL, Amersham Pharmacia Biotech, Piscataway, NJ, USA) and exposed to x-ray film according to the manufacturer's instructions.
Statistical methods
Results are presented as means ± standard error of the mean (SEM). Continuous data were analyzed using student's t-test or analysis of variance followed by Fisher's least significant difference test. P values less than 0.05 were considered significant. Summary statistics are presented for densitometry results from studies using western blot for cyclin D1 expression but these results were not subjected to statistical analysis because the method employed was only semi-quantitative (n = 6).
Results
Prolonged treatment with NAC increases serum ALT/AST at 72-hour time point
Twenty-four hours after APAP injection, compared with saline treatment, NAC therapy significantly decreased serum concentrations of ALT/AST (Figure 1a, b). However, 72 hours after APAP challenge, compared with saline treatment, NAC therapy significantly increased serum ALT/AST concentrations (Figure 1c, d).

Effect of treatment with NAC or saline on serum ALT/AST in an APAP-induced ALI model. (a, b) Acute liver injury (ALI) was induced in C57Bl/6 male mice with a single dose of acetaminophen (APAP) (350 mg/kg) by intraperitoneal injection. Two hours after APAP injection, the animals were treated with 100 mg/kg N-acetyl-cysteine (NAC) dissolved in 0.6 mL saline or 0.6 mL saline every 12 hours. Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were measured 24 hours after APAP injection (n = 6 surviving mice for each group). Results are means ± standard error of the mean (SEM). * P < 0.05 versus control; † P < 0.05 vs. the NAC group. (c, d) Three separate groups of mice were used. ALI was induced as described above. Two hours after APAP challenge, the animals were given the same treatment every 12 hours for a total of 72 hours. ALT and AST were measured 72 hours after APAP injection (n = 6 to 7 surviving mice for each group). Results are means ± SEM. * P < 0.05 vs. control; † P < 0.05 vs. the NAC group.
Prolonged treatment with NAC impairs liver regeneration in histopathology
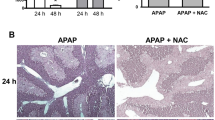
In histological evaluation 72 hours after ALI induction, compared with control animals, saline-treated mice demonstrated a small number of scattered necrotic hepatocytes, evident regeneration and extensive infiltration of inflammatory cells (240 ± 40 per high power field, n = 6) in the centrilobular region. In contrast, NAC-treated mice demonstrated 10% necrotic hepatocytes and extensive inflammatory cell infiltration (250 ± 35 per high power field, n = 6) in the centrilobular region; however, no evident regeneration was seen (Figure 2; arrows indicate periportal areas shown in Figure 3). In addition, prolonged treatment with NAC also induced evident hepatocyte vacuolation in the periportal area (Figure 3).

Effect of treatment with NAC or saline on pathology in mice with ALI. H&E staining was assessed 72 hour after induction of acute liver injury (ALI) or sham procedure (n = 6 for each group). Typical picture is shown. Arrows indicate periportal areas shown in Figure 3. NAC = N-acetyl-cysteine.

Prolonged treatment with NAC induces periportal hepatocytes vacuolation in mice with ALI. H&E staining was assessed 72 hour after induction of acute liver injury (ALI) or sham procedure (n = 6 for each group). Typical picture is shown. NAC = N-acetyl-cysteine.
Prolonged treatment with NAC increases hepatic tissue MPO level
Tissue MPO activity was determined as an index of neutrophil infiltration after the APAP injection in the liver. Liver MPO activity values for the control group were 4.2 ± 0.29 U/g (Figure 4). Seventy-two hours after ALI, these values increased to 5.88 ± 1.19 U/g in the saline group. The liver MPO activity was significantly increased in the NAC therapy group, to a value of 9.5 ± 0.70 U/g (P < 0.05), when compared with the saline group (n = six for each group, data were shown as mean ± SEM).

Effect of treatment with NAC or saline on hepatic MPO activity in mice with ALI. Liver myeloperoxidase (MPO) was assessed 72 hours after induction of acute liver injury (ALI) or sham procedure. Results are means ± standard error of the mean (SEM). * P < 0.05 versus control; † P < 0.05 vs. saline. NAC = N-acetyl-cysteine.
Prolonged NAC therapy decreases hepatic NF-κB DNA binding
NF-κB is a pleiotropic transcription factor whose activation has been linked to inflammatory and destructive processes, as well as initiation of regenerative programs in the injured liver. Blockade of HMGB1 protects against ischemia-reperfusion (I/R)-induced liver injury; this protection is associated with increased NF-κB DNA binding activity [20]. Enhanced NF-κB activation is seen in mice that are protected from hepatic I/R following blockade of the HMGB1 receptor for advanced glycation end products [21]. Therefore, we examined the impact of APAP on activation of NF-κB 72 hours after APAP injection and tested the effect of NAC treatment. There was a low basal level of NF-κB DNA binding in the hepatic tissue samples in the control group. In the saline group, there was a marked increase in NF-κB DNA binding. Prolonged treatment of mice after APAP challenge with NAC clearly decreased NF-κB DNA binding relative to the degree observed in mice treated with saline (Figure 5).

Effect of treatment with NAC or saline on NF-κB DNA binding in nuclear extracts prepared from hepatic tissue samples from mice with ALI. Nuclear factor (NF) κB DNA binding was assessed 72 hours after induction of acute liver injury (ALI) or sham procedure. The figure depicts results from five representative assays. Typical gels are depicted. NAC = N-acetyl-cysteine.
Prolonged treatment with NAC decreases hepatic cyclin D1 expression
The timely onset of tissue repair processes can limit liver injury and promote regeneration of lost tissue mass [7]. The induction of cyclin D1 is the most reliable marker for cell cycle (G1 phase) progression in hepatocytes [8]. Western blot was performed using whole-cell extracts prepared from liver tissue to assess expression of cyclin D1 in mice subjected to ALI or the control procedure. Cyclin D1 expression in the control group and the NAC group was minimal (Figure 6). In contrast, cyclin D1 expression was clearly observed in saline-treated animals at 72 hours after APAP administration, which indicates that prolonged treatment with NAC inhibits hepatic cyclin D1 expression.

Effect of treatment with NAC or saline on the expression of cyclin D1 in the hepatic tissue. Western blot was performed using hepatic extracts prepared from tissues obtained 72 hours after acetaminophen injection. The figure depicts results from five representative assays. Typical gels are depicted. NAC = N-acetyl-cysteine.
Discussion
The purpose of this study was to test the hypothesis that prolonged treatment with NAC impairs liver regeneration in ALI induced by APAP. The major and the novel findings of this investigation are: prolonged treatment with NAC increases serum ALT/AST and delays liver regeneration 72 hours after APAP administration; prolonged treatment with a therapeutic dose of NAC induces evident hepatocyte vacuolation around periportal area; the detrimental effect is associated with reduced NF-κB DNA binding; and prolonged NAC therapy significantly decreases the expression of cell cycle protein cyclin D1 in liver tissue.
In this study, prolonged NAC treatment demonstrated a large amount of hepatonecrosis in the centrilobular region and evident hepatocyte vacuolation in the periportal area; and also induced increased neutrophil infiltration as determined by measuring myeloperoxidase activity in total liver extracts. In addition, no evident regeneration was seen in the NAC group by histopathology. In contrast, saline treatment showed only a small number of scattered necrotic hepatocytes and evident regeneration. These data indicate that liver regeneration is delayed by prolonged NAC therapy and support the notion that timely onset of tissue repair processes can limit liver injury [7].
To elucidate the molecular basis of liver recovery in prolonged NAC therapy, we investigated its effect on the NF-κB signaling pathway because activation of NF-κB is linked strongly not only to the inflammatory response [20], but also to liver regeneration [8]. In addition, NF-κB is currently thought to play a major role in the initiation of liver regeneration after cell or tissue loss (such as partial hepatectomy) [8, 22]. NF-κB activation also induces increased expression of survival genes, including BCLXL and A1 [23]. Our data suggested that prolonged treatment with NAC is associated with a detrimental effect characterized by reduced NF-κB DNA binding. Although NF-κB activation modulates inflammation [24], it is also known to protect hepatocytes from cell death, and inhibition of NF-κB after partial hepatectomy results in massive hepatocyte apoptosis, worsens liver injury and decreases survival [25]. Enhanced NF-κB activation is also seen in mice that are protected from hepatic I/R following blockade of the receptor for advanced glycation end products [21]. There is evidence suggesting that the impact of APAP toxicity ensues, at least in part, by dramatic modulation of inflammatory and/or regeneration programs [26]. After massive hepatonecrosis induced by APAP overdose, liver regeneration is a vital process for survival after a toxic insult [6, 7], and NF-κB activation plays an important role in liver regeneration [8]. Prolonged treatment with NAC impairs liver regeneration, at least partly by inhibiting the NF-κB activation pathway.
Massive hepatocyte necrosis is the predominant feature of APAP-induced ALI. Tissue repair is an important determinant of final outcome of toxicant-induced injury [7], and cyclin D1 is an important cell cycle protein. In the current investigation, our western blot data showed that prolonged NAC treatment markedly decreased the level of cyclin D1 expression in the APAP-challenged liver tissue. The change in cyclin D1 expression was associated with increased serum ALT/AST and delayed liver regeneration in NAC-treated mice receiving APAP, suggesting that prolonged NAC therapy likely inhibits cyclin D1-mediated regeneration pathway, and the reduced cyclin D1 expression might be modulated by decreased NF-κB DNA binding [8].
Conclusions
Prolonged treatment with NAC impairs liver regeneration in ALI induced by APAP, reevaluation of optimal doses and duration of NAC therapy is needed.
Key messages
-
Prolonged treatment with NAC increases serum ALT/AST in APAP-challenged mice.
-
Prolonged treatment with NAC induces hepatocyte vacuolation in APAP-challenged mice.
-
Prolonged treatment with NAC delays hepatocyte regeneration in APAP-challenged mice.
-
Prolonged treatment with NAC delays liver recovery from APAP hepatotoxicity, reevaluation of optimal doses and duration of NAC therapy is needed.
Abbreviations
- APAP:
-
acetaminophen
- ALI:
-
acute liver injury
- ALT:
-
alanine aminotransferase
- AST:
-
aspartate aminotransferase
- EMSA:
-
electrophoretic mobility shift assays
- GSH:
-
glutathione
- H&E:
-
haematoxylin and eosin
- I/R:
-
ischemia-reperfusion
- MPO:
-
myeloperoxidase
- NAC:
-
N-acetyl-cysteine
- NAPQI:
-
N-acetyl-p-benzoquinone imine
- NF-κB:
-
nuclear factor κB
- PBS:
-
phosphate-buffered saline
- SEM:
-
standard error of the mean
- TCA:
-
tricarboxylic acid.
References
Lee WM: Acetaminophen and the U.S. Acute liver failure study group: lowering the risks of hepatic failure. Hepatology 2004, 40: 6-9. 10.1002/hep.20293
Nelson SD: Molecular mechanisms of the hepatotoxicity caused by acetaminophen. Semin Liver Dis 1990, 10: 267-278. 10.1055/s-2008-1040482
Cohen SD, Khairallah EA: Selective protein arylation and acetaminophen-induced hepatotoxicity. Drug Metab Rev 1997, 29: 59-77. 10.3109/03602539709037573
Cressman DE, Greenbaum LE, DeAngelis RA, Ciliberto G, Furth EE, Poli V, Taub R: Liver failure and defective hepatocyte regeneration in interleukin-6 deficient mice. Science 1996, 274: 1379-1383. 10.1126/science.274.5291.1379
Jaeschke H, Bajt ML: Intracellular signaling mechanisms of acetaminophen-induced liver cell death. Toxicol Sci 2006, 89: 31-41. 10.1093/toxsci/kfi336
Chanda S, Mehendale HM: Hepatic cell division and tissue repair: a key to survival after liver injury. Mol Med Today 1996, 2: 82-89. 10.1016/1357-4310(96)88743-0
Mehendale HM: Tissue repair: An important determinant of final outcome of toxicant-induced injury. Toxicol Pathol 2005, 33: 41-51. 10.1080/01926230590881808
Fausto N: Liver regeneration. J Hepatol 2000, 32: 19-31. 10.1016/S0168-8278(00)80412-2
Whyte IM, Francis B, Dawson AH: Safety and efficacy of intravenous N-acetylcystine for acetaminophen overdose: analysis of the Hunter Area Toxicity service (HATS) Database. Curr Med Res Opin 2007, 23: 2359-2368. 10.1185/030079907X219715
Kerr F, Dawson A, Whyte IH, Buckley N, Murray L, Graudins A, Chan B, Trudinger B: The australasian clinical toxicology investigators collaboration. Randomized trial of different loading infusion rates of N-acetylcystine. Ann Emerg Med 2005, 45: 402-408. 10.1016/j.annemergmed.2004.08.040
Hogaboam CM, Bone-Larson CL, Steinhauser ML, Lukacs NW, Colletti LM, Simpson KJ, Strieter RM, Kunkel SL: Novel CXCR2-dependent liver regenerative qualities of ELR-containing CXC chemokines. FASEB J 1999, 13: 1565-1574.
Su GL, Gong KQ, Fan MH, Kelley WM, Hsieh J, Sun JM, Hemmila MR, Arbabi S, Remick DG, Wang SC: Lipopolysaccharide-binding protein modulates acetaminophen-induced liver injury in mice. Hepatology 2005, 41: 187-195. 10.1002/hep.20533
Zwingmann G, Bilodeau M: Metabolic insights into the hepatoprotective role of N-acetylcysteine in mouse model. Hepatology 2006, 43: 454-463. 10.1002/hep.21075
Sheetal Vaghjiani: Functions of the liver. In Drugs and the liver. Edited by: North-Lewis P. Chicago: Pharmaceutical Press; 2008:24-48.
Tong TC, Hernandez M, Richardson WM, Betten DP, Favata M, Riffenburgh RH, Clark RF, Tanen DA: Comparative treatment of α-amanitin poisoning with N-acetylcysteine, benzylpenicillin, cimetidine, thioctic acid and silybin in a murine model. Ann Emerg Med 2007, 50: 282-288. 10.1016/j.annemergmed.2006.12.015
Kortsalioudaki C, Taylor RM, Cheeseman P, Bansal S, Mieli-Vergani G, Dhawan A: Safety and efficacy of N-acetylcysteine in children with non-acetaminophen-induced acute liver failure. Liver transpl 2008, 14: 25-30. 10.1002/lt.21246
Martinez-Mier G, Toledo-Pereyra LH, McDuffie E, Warner RL, Ward PA: L-selectin and chemokine response after liver ischemia and reperfusion. J Surg Res 2000, 93: 156-162. 10.1006/jsre.2000.5954
Yang R, Gallo DJ, Baust JJ, Uchiyama T, Watkins SK, Delude RL, Fink MP: Ethyl pyruvate modulates inflammatory gene expression in mice subjected to hemorrhagic shock. Am J Physiol Gastrointest Liver Physiol 2002, 283: G212-G222.
Yang R, Harada T, Mollen KP, Prince JM, Levy RM, Englert JA, Galloitsch-Peerta M, Yang L, Yang H, Tracey KJ, Harbrecht BG, Billiar TR, Fink MP: Anti-HMGB1 neutralizing antibody ameliorates gut barrier Dysfunction and improves survival after hemorrhagic shock. Mol Med 2006, 12: 105-114. 10.2119/2006-00010.Yang
Tsung A, Sahai R, Tanaka H, Nakao A, Fink MP, Lotze MT, Yang H, Li J, Tracey KJ, Geller DA, Billiar TR: The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J Exp Med 2005, 201: 1135-1143. 10.1084/jem.20042614
Zeng S, Feirt N, Goldstein M, Guarrera J, Ippagunta N, Ekong U, Dun H, Lu Y, Qu W, Schmidt AM, Emond JC: Blockade of receptor for advanced glycation end product (RAGE) attenuates ischemia and reperfusion injury to the liver in mice. Hepatology 2004, 39: 422-432. 10.1002/hep.20045
Cataldegirmen G, Zeng S, Feirt N, Ippagunta N, Dun H, Qu W, Lu Y, Rong L, Hofmann MA, Kislinger T, Pachydaki SI, Jenkins DG, Weinberg A, Lefkowitch J, Rogiers X, Yan S, Schmidt AM, Emond JC: RAGE limits regeneration after massive liver injury by coordinated suppression of TNF-α and NF-κB. J Exp Med 2005, 201: 473-484. 10.1084/jem.20040934
Malhi H, Gores GJ, Lemasters JJ: Apoptosis and necrosis in the liver: a tale of two deaths? Hepatology 2006, 43: S31-44. 10.1002/hep.21062
Ulloa L, Ochani M, Yang H, Tanovic M, Halperin D, Yang R, Czura CJ, Fink Mp, Tracey KJ: Ethyl pyruvate prevents lethality in mice with established lethal sepsis and systemic inflammation. Proc Natl Acad Sci 2002, 99: 12351-12356. 10.1073/pnas.192222999
Iimuro Y, Nishiura T, Hellerbrand C, Behrns KE, Schoonhoven R, Grishma JW, Brenner DA: NF kappa B prevents apoptosis and liver dysfunction during liver regeneration. J Clin Invest 1998, 101: 802-811. 10.1172/JCI483
Kaplowitz N: Acetaminophen hepatotoxicity. What do we know, what don't we know, and what do we do next? Hepatology 2004, 40: 23-26. 10.1002/hep.20312
Acknowledgements
This study was supported by funding from the Department of Critical Care Medicine, University of Pittsburgh.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
RKY designed the study. All authors participated in the animal handling and procedures. MPF helped to draft the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Yang, R., Miki, K., He, X. et al. Prolonged treatment with N-acetylcystine delays liver recovery from acetaminophen hepatotoxicity. Crit Care 13, R55 (2009). https://doi.org/10.1186/cc7782
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/cc7782