
Abstract
Introduction
Suppression of Bcl-2 expression can overcome cellular resistance to apoptosis induced by the adenovirus type 5 gene E1A in models of ovarian and breast cancer. Celecoxib, a cyclooxygenase-2 (COX-2) inhibitor, is known to downregulate Bcl-2 expression. We hypothesized that celecoxib would enhance E1A-induced apoptosis by suppressing Bcl-2 through suppressing COX-2 expression. If successful, this strategy could represent a means of overcoming resistance to E1A gene therapy.
Methods
We first established the cytotoxicity of celecoxib in two COX-2-overexpressing E1A-transfected breast cancer cell lines (MDA-MB-231 and MDA-MB-435) and in two low-COX-2-expressing E1A-transfected cell lines (MCF-7 (breast cancer) and SKOV3.ip1 (ovarian cancer)). We next tested whether higher sensitivity to celecoxib among these cell lines resulted from increased apoptosis by flow cytometry and western blotting. We further investigated whether suppression of Bcl-2 by celecoxib was involved in the apoptosis resulting from celecoxib treatment, and we explored whether the celecoxib-induced apoptosis in these cells depends on a COX-2 downstream pathway.
Results
The two COX-2-overexpressing cell lines MDA-MB-231-E1A and MDA-MB-435-E1A were more sensitive to celecoxib than the corresponding control cells, but the two low-COX-2-expressing cell lines MCF-7-E1A and SKOV3.ip1-E1A were no more sensitive than control cells to celecoxib. Therefore, we used the MDA-MB-231-E1A and MDA-MB-435-E1A cells for all further experiments. In both cell lines, sub-G1 fraction was increased, or cleavage of PARP and caspase-9 were increased after 5 days of exposure to 40 μM celecoxib. However, Bcl-2 was suppressed only in the MDA-MB-435-E1A cells and not in the MDA-MB-231-E1A cells. Restoring Bcl-2 expression in the MDA-MB-435-E1A stable transfectants did not affect their sensitivity to celecoxib. However, adding prostaglandin E2 (PGE2) or PGF2α blunted the sensitivity to celecoxib of both E1A stable transfectants.
Conclusion
We speculate that one mechanism by which celecoxib enhances E1A-induced apoptosis in cells that express high levels of COX-2 is through blocking PGE2 or PGF2α.
Similar content being viewed by others


Introduction
The adenovirus type 5 gene E1A is being developed as a therapeutic agent for breast, head and neck, and ovarian cancer [1–3]. The tumor-suppressive effect of E1A results from its induction of apoptosis, its inhibition of invasion and metastasis, and its suppression of proliferation [4]. Although the mechanism by which E1A induces apoptosis is not completely understood, accumulating evidence suggests that E1A exerts its apoptotic effect through several pathways. One such pathway involves binding of E1A to the retinoblastoma protein, which results in the release and activation of transcription factor E2F [4–6]. Activated E2F can induce apoptosis through p53-dependent and p53-independent pathways [7–9]. E1A is also known to induce apoptosis through p53-dependent and p53-independent pathways [10]. Several factors that modulate E1A-induced apoptosis have been reported, including tumor necrosis factor α (TNF-α) [11, 12], TNF-related apoptosis-inducing ligand (TRAIL) [13] or TRAIL accompanied by caspase activation [14], activation of the pro-apoptotic factor p38 or inactivation of Akt/PKB [15], inhibition of nuclear factor-kappa B [16], and suppression of the Axl-Gas6 interaction [17].
However, E1A does not induce apoptosis efficiently in all types of cancer cells. The reason for this is unclear but could reflect the existence of a resistance mechanism or the oncogenic effects of the E1A gene. We recently showed that resistance to E1A gene therapy in an ovarian cancer xenograft model could be overcome by downregulating Bcl-2 with a Bcl-2 antisense oligonucleotide [18]. Bcl-2 inhibits apoptosis by inhibiting the release of cytochrome c and the activation of caspase-9 in E1A-transfected cells. Bcl-2 was the only major mechanism blocking E1A-induced apoptosis in our previous model. Theoretically, a means of downregulating Bcl-2 would overcome resistance to the apoptosis induced by E1A gene therapy. Because the US Food and Drug Administration has yet to approve Bcl-2 antisense oligonucleotide for clinical use, we are exploring other drugs that can also downregulate Bcl-2. Recent reports that cyclooxygenase-2 (COX-2) inhibitors could induce apoptosis through Bcl-2 downregulation led us to consider the role of COX-2 inhibitors in the E1A-induced apoptosis of cancer cells. In those reports, the selective COX-2 inhibitor NS-398 was found to downregulate Bcl-2 in LNCaP prostate cancer cells [19]; another COX-2 inhibitor, celecoxib, downregulated Bcl-2 in K562 chronic myeloid leukemia cells [20] and in MPP89 malignant mesothelioma cells [21]. In vivo investigations have shown that Bcl-2 downregulation by COX-2 inhibitors is accompanied by downregulation of the COX-2 protein. In one study of mice implanted with the hepatoma cell line H22, treatment with the COX-2 inhibitor nimesulide led to reductions in both COX-2 and Bcl-2 expression [22]. Similarly, celecoxib also reduced both COX-2 and Bcl-2 expression in an MTag mouse model of breast cancer [23]. Conversely, another group has proposed that COX-2 overexpression increases resistance to apoptosis through the upregulation of Bcl-2 [24]. Still others have shown that forced COX-2 overexpression or treatment with prostaglandins induces Bcl-2 expression [25, 26]. Collectively, these reports suggest that COX-2 is upstream of Bcl-2 and led us to propose that suppressing COX-2 expression with the COX-2 inhibitor celecoxib will suppress Bcl-2 expression, thereby enhancing E1A-induced apoptosis. If successful, this strategy could represent a means of overcoming resistance to E1A gene therapy.
Materials and methods
Cell lines and reagents
Three human breast cancer cell lines (MDA-MB-231, MDA-MB-435, and MCF-7) and one ovarian cancer cell line (SKOV3.ip1, a subline of SKOV3 cells) were maintained in Dulbecco's modified Eagle's medium/Ham's F-12 medium (DMEM/F12; Gibco-BRL, Grand Island, NY, USA) supplemented with 10% fetal bovine serum and penicillin/streptomycin, and maintained in a humidified atmosphere of 5% CO2 at 37°C. The parental cell lines, the vector only transfection control cells, and the E1A-transfected cells were all kindly provided by Dr Mien-Chie Hung (The University of Texas M D Anderson Cancer Center, Houston, TX, USA) [15, 27, 28]. The E1A stable transfectants were selected by growing them in DMEM/F12 medium containing 500 μg/ml G418. All experiments were conducted under the guidelines of the M D Anderson Cancer Center.
Celecoxib (LKT Laboratories, St Paul, MN, USA) was dissolved in dimethylsulfoxide to 100 mM and stored at -20°C. For the cell culture experiments, celecoxib was diluted with DMEM/F12 in various concentrations. The final concentration of dimethylsulfoxide in the DMEM/F12 medium was kept at less than 0.1%. PGE2 and PGF2α (Cayman Chemical, Ann Arbor, MI, USA) were dissolved in dimethlysulfoxide to 10 mM and stored at -20°C.
Cell viability assays
Cell viability was assessed with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) [29] and trypan blue exclusion assays. For the MTT assay, cells were plated in 96-well plates (1.2 × 104 cells/well for MDA-MB-231 cells or 8.0 × 103 cells/well for MDA-MB-435, MCF-7, and SKOV3.ip1 cells) in 80 μl of medium and incubated for 24 h. Then, 20 μl of fresh medium containing celecoxib at 0–300 μM were added to each well, resulting in final celecoxib concentrations of 0–60 μM in five sequential dilutions. After cells had been cultured with celecoxib for 3 or 5 days, MTT (Sigma Chemical Co., St Louis, MO, USA) was added to a final concentration of 1 mg/ml. Reaction mixtures were incubated for 3 h. The resulting crystals were dissolved in dimethlysulfoxide (200 μl), and optical density was measured at 570 nm with a microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
We also used a trypan blue exclusion assay to confirm cell viability during treatment with 40 μM celecoxib, because the viability varied greatly among cell types. For that assay, MDA-MB-231 cells (4 × 105) or MDA-MB-435 cells (2 × 105) were plated in six-well plates in 2 ml of DMEM/F12 and incubated for 24 h. Then, 500 μl of fresh medium containing 200 μM celecoxib was added to each well, resulting in a final concentration of 40 μM, and the cells were incubated for 5 days. To determine the effect of PGE2 or PGF2α on celecoxib sensitivity, exogenous PGE2 or PGF2α (10 μM) was added to the celecoxib solution. After being cultured with celecoxib in the presence or absence of PGE2 or PGF2α for 5 days, cells were harvested by trypsinization and incubated with 0.4% trypan blue (Sigma). Cell viability was calculated as the percentage of viable (nonstaining) cells.
Flow cytometry
For flow-cytometric analysis of apoptosis, cells were harvested by trypsinization, washed twice with ice-cold PBS, and fixed with cold 70% ethanol at -20°C overnight. The fixed cells were washed twice with PBS and suspended in 1 ml of PBS containing Tween-20 (0.5%), RNase (10 μg/ml), and propidium iodide (10 μg/ml). The sub-G1 (apoptotic) cell population was measured with a FACScan cytofluorometer (Becton Dickinson, San Jose, CA, USA).
Western blotting
For western blot analyses, cells were washed with PBS and lysed in lysis buffer (20 mM Na2PO4, 150 mM NaCl, 1% Triton X-100, 1% aprotinin, 1 mM phenylmethylsulfonyl fluoride, 100 mM NaF, and 2 mM Na3VO4) as described previously [18]. Proteins were separated by polyacrylamide gel electrophoresis on a sodium dodecyl sulfate-polyacrylamide gel and transferred to a polyvinylidene difluoride membrane (Bio-Rad). Membranes were incubated with primary antibodies specific for COX-2 (1:1000; Cayman Chemical); Bcl-2 (1:500) and E1A (1:500) (both from BD PharMingen); poly-ADP ribosome polymerase (PARP) (1:1000) and cleaved caspase-9 (1:200) (both from Cell Signaling Technology, Beverly, MA, USA); caspase-8 (1:200) (from Oncogene Research Products, San Diego, CA, USA); and actin (1:5000) (Sigma). Then, membranes were incubated with fluorescent-conjugated mouse (1:5000) or rabbit (1:5000) secondary antibodies (IRdye; Li-Cor Biosciences, Lincoln, NE, USA). The membranes were scanned and relative protein expression levels estimated by using an Odyssey western blotting system (Li-Cor Biosciences). Downregulation was defined as a protein expression level at least 20% less than that of the control (untreated) cells.
Bcl-2 transfections
The Bcl-2 expression vector was constructed by inserting Bcl-2 cDNA at the EcoRI site of the pCl-neo mammalian expression vector (Promega, Madison, WI, USA). Briefly, MDA-MB-435 cells (4 × 105) were cultured for 24 h in six-well plates in 1 ml/well of DMEM/F12 with 10% fetal bovine serum until 60–70% confluence. The next day, the liposomal vector of the SN gene delivery system [30] was incubated with Bcl-2 DNA at a ratio of 4 μg DNA: 5 μl SN in 100 μl Opti-MEM in each well and added to the cultures. Bcl-2-overexpressing cells and control cells were plated 24 h later and tested for viability with a trypan blue exclusion assay as described above.
Prostaglandin analyses
MDA-MB-231-E1A cells (4 × 105) or MDA-MB-435-E1A cells (2 × 105) were plated in six-well plates in 2 ml of DMEM/F12, incubated for 24 h, and treated with celecoxib (0–40 μM) for 120 h. At that time, cellular supernatants were collected and analyzed for PGE2 and PGF2α by enzyme-linked immunosorbent assay (Cayman Chemical) according to the manufacturer's instructions.
Statistical analyses
Two-tailed paired t-tests were used to compare data between groups. P < 0.05 was considered to be statistically significant. Data were expressed as means ± SD of at least three independent experiments, each of which was run in quadruplicate.
Results
COX-2 expression in E1A-transfected breast and ovarian cancer cell lines
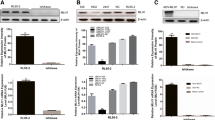
We first investigated the expression of COX-2 protein in three breast cancer cell lines (MDA-MB-231, MDA-MB-435, and MCF-7) and an ovarian cancer cell line (SKOV3.ip1), all stably transfected with E1A, to determine the relationship between COX-2 protein expression level and sensitivity to celecoxib. Western blot analyses showed that COX-2 expression was highest in the MDA-MB-231-E1A transfectants, followed closely by the MDA-MB-435-E1A transfectants. By contrast, COX-2 expression was low in the SKOV3.ip1-E1A and MCF-7-E1A transfectants (Figure 1A). If the COX-2 expression level of the MDA-MB-231-E1A stable transfectants is considered to be 100%, the relative percentage COX-2 expression for each stable transfectant is as follows: MDA-MB-435-E1A, 78%; SKOV3.ip1-E1A, 6%; and MCF-7-E1A, 6%. Thus, we defined MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants as high-COX-2-expressing cell lines, and SKOV3.ip1-E1A and MCF-7-E1A stable transfectants as low-COX-2-expressing cell lines.

COX-2 protein expression in breast and ovarian cancer cell lines stably transfected with E1A. (a) MDA-MB-231-E1A stable transfectants produced the greatest amounts of COX-2; MDA-MB-435-E1A cells produced 78% of that amount, but the SKOV3.ip1-E1A and MCF-7-E1A cells produced only 6% of that amount. MDA-MB-231-E1A cells expressed slightly more E1A than the other three cell lines. (a) COX-2 protein expression level between the E1A stable transfectants and their corresponding vector control cells or parent cells. If the COX-2 expression levels of each E1A transfectant is defined as 100%, the corresponding COX-2 expression levels of the vector controls were as follows: 65% for MDA-MB-231, 144% for MDA-MB-435, 71% for SKOV3.ip1 and 67% for MCF-7.
To rule out the possibility that E1A transfection affects COX-2 expression levels, we compared COX-2 levels in E1A stable transfectants with that in the corresponding vector control cells by western blotting analysis. If the COX-2 expression levels of each E1A transfectant is defined as 100%, the corresponding COX-2 expression levels of the vector controls were as follows: 65% for MDA-MB-231, 144% for MDA-MB-435, 71% for SKOV3.ip1 and 67% for MCF-7 (Figure 1B). Thus, we found that E1A transfection did not consistently affect COX-2 expression levels in these cell lines.
Celecoxib induces apoptosis in MDA-MB-231-E1A and MDA-MB-435-E1Astable transfectants
After establishing the relative amounts of COX-2 protein expressed by the E1A stable transfectants, we investigated whether those transfectants were more sensitive to celecoxib than their respective controls by using an MTT assay. The high-COX-2-expressing MDA-MB-231-E1A and MDA-MB-435-E1A cells were more sensitive to celecoxib (after 5 days of exposure to 0–60 μM) than the vector control or parental control cells (Figure 2A). By contrast, the low-COX-2-expressing cells (SKOV3.ip1 and MCF-7) showed no difference in celecoxib sensitivity between the E1A transfectants and the controls (Figure 2B).

Celecoxib decreases the viability of MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants. (b) MTT assays after a 5-day exposure to 0–60 μM celecoxib indicate substantial reductions in cell viability in all variants of the MDA-MB-231 and MDA-MB-435 cell lines (E1A stable transfectants, vector control, and parental control cells). Each point represents means from tests performed in quadruplicate; the bars are standard deviations. In both cell lines, the E1A transfectants were more sensitive than the vector control or parental control cells. (b) MTT assays in all variants of the SKOV3.ip1 and MCF-7 cell lines. The E1A transfectants have no difference to sensitivity for celecoxib in both cell lines. (c) Trypan blue assays after a 5-day exposure to 40 μM celecoxib show substantial reductions in viability of MDA-MB-231 and MDA-MB-435 cells. Values shown are normalized to the viability of the control (untreated) cells. Each bar represents means from tests performed in quadruplicate; bars are standard deviations. P values are from two-tailed paired t tests.
To quantitatively compare differences in sensitivity to celecoxib among cells, we tested the MDA-MB-231-E1A and MDA-MB-435-E1A transfectants and their controls with a trypan blue assay. After a 5-day exposure to 40 μM celecoxib, the MDA-MB-231-E1A cells were significantly more sensitive (mean 23.8% ± SD 2.2% viable cells) than the MDA-MB-231 vector control cells (82.3% ± 3.9%) (P = 0.0002) or the MDA-MB-231 parental cells (82.0% ± 1.4%) (P < 0.0001). Similarly, the MDA-MB-435-E1A stable transfectants were more sensitive to celecoxib (27.2% ± 1.2% viable cells) than the MDA-MB-435 vector control cells (91.2% ± 0.5%) (P < 0.0001) or the MDA-MB-435 parental cells (93.9% ± 3.5%) (P < 0.0001) (Figure 2C). We then investigated whether these differences in viability had been caused by apoptosis by using flow cytometry. A 5-day exposure to 40 μM celecoxib increased the sub-G1 fraction from 9.1% to 25.7% in MDA-MB-231-E1A cells and from 8.2% to 37.1% in MDA-MB-435-E1A cells (Figure 3A), indicating increases in apoptosis. We also tested cleavage of PARP, caspase-8, and caspase-9 as other indicators of apoptosis. Celecoxib treatment led to the disappearance of uncleaved PARP (116 kDa) in both cell lines; it also led to the appearance of cleaved PARP (89 kDa) in MDA-MB-435-E1A cells and the appearance of cleaved PARP expression in MDA-MB-231-E1A cells (Figure 3B). Celecoxib treatment also led to increases in cleaved caspase-9 (37 kDa) levels in both MDA-MB-231-E1A and MDA-MB-435-E1A transfectants but did not affect levels of cleaved caspase-8 (28 kDa). These results suggest that celecoxib treatment (5 days at 40 μM) induced apoptosis in MDA-MB-231 and MDA-MB-435 cells stably transfected with E1A.

Celecoxib enhances apoptosis of MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants. (a) Cell cycle distribution of MDA-MB-231-E1A and MDA-MB-435-E1A cells was detected by fluorescence-activated cell sorting after a 5-day exposure to 0 or 40 μM celecoxib. The percentage of cells in sub-G1 (apoptosis) appears at the upper right of each graph. (b) Western blots of MDA-MB-231 and MDA-MB-435 cells treated with 0 or 40 μM celecoxib for 5 days and tested for cleaved caspase-9 (cl-cas-9), uncleaved and cleaved caspase-8 (cl-cas-8), PARP (uncleaved and cleaved), E1A, and actin. Cleaved PARP and cleaved caspase-9 levels were higher after celecoxib treatment in the MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants, but expression of cleaved caspase-8 (cl-cas-8) did not change.
Celecoxib downregulates COX-2 protein expression in MDA-MB-231 and MDA-MB-435 cells
Evidence that nonsteroidal anti-inflammatory drugs can suppress COX-2 by transcriptional regulation [31] as well as suppressing both COX-2 and Bcl-2 protein expression [22, 23] led us to investigate whether celecoxib would change the amount of COX-2 protein expressed by MDA-MB-231 and MDA-MB-435 cells. Indeed, COX-2 protein expression was downregulated in all MDA-MB-231 cell variants; the percentage decreases were 32% for the MDA-MB-231 parental cells, 34% for the vector control cells, and 58% for the E1A stable transfectants (Figure 4A). COX-2 protein expression was also decreased in all MDA-MB-435 variants, with the percentages being 39% for MDA-MB-435 parental cells, 25% for the vector control cells, and 60% for the E1A stable transfectants (Figure 4A). In both MDA-MB-231 and MDA-MB-435 cells, COX-2 was suppressed to a greater extent in the E1A transfectants than in the corresponding parental or vector control cells. These results suggest that celecoxib-induced apoptosis in cells expressing E1A involves the suppression of COX-2.

Celecoxib downregulated COX-2 protein expression in all MDA-MB-231 and MDA-MB-435 variants, but celecoxib downregulated Bcl-2 expression in only the MDA-MB-435-E1A stable transfectants. (a) Western blots of MDA-MB-231 and MDA-MB-435 cells treated with 0 or 40 μM celecoxib for 5 days and tested for COX-2 and Bcl-2. Percentages indicate differences relative to the 0 μM control samples. Protein expression was considered to be downregulated if the treated condition was at least 20% less than the control (untreated) condition. (b) Time course of Bcl-2 expression after treatment with 0 or 40 μM celecoxib in MDA-MB-435-E1A and MDA-MB-231-E1A stable transfectants. Bcl-2 was suppressed at both 72 and 96 h in the MDA-MB-435-E1A stable transfectants but was not suppressed in the MDA-MB-231-E1A stable transfectants. (c) Transfection of MDA-MB-435 cells with Bcl-2 DNA (+) or a control DNA (-) led to overexpression of Bcl-2 in all variants. (d) MDA-MB-435-E1A cells made to overexpress Bcl-2 and non-Bcl-2-overexpressing cells were treated with 0 or 40 μM celecoxib for 5 days, and cell viability was determined with a trypan-blue assay. Bcl-2 overexpression did not restore sensitivity to celecoxib (P = 0.11).
Bcl-2 suppression does not contribute to celecoxib-induced apoptosis in E1Astable transfectants
Next, we investigated whether suppression of Bcl-2 by celecoxib is involved in the apoptosis resulting from celecoxib treatment. After a 5-day treatment with 40 μM celecoxib, Bcl-2 levels in the MDA-MB-435 cells were decreased (by 27%) only in the E1A transfectant; Bcl-2 level did not change in any of the MDA-MB-231 variants (Figure 4A). In timecourse experiments with the same celecoxib concentration (40 μM), Bcl-2 was suppressed by 37% at 72 h and by 52% at 96 h in MDA-MB-435-E1A cells. However, Bcl-2 was not suppressed at either measurement time in the MDA-MB-231-E1A cells (Figure 4B).
As a further step in determining the contribution of Bcl-2 suppression to celecoxib-induced apoptosis, we transfected Bcl-2 DNA into the MDA-MB-435 variants (Figure 4C) to see if restoring Bcl-2 expression would affect sensitivity to celecoxib. Bcl-2 restoration did not affect the viability of MDA-MB-435-E1A stable transfectants after a 5-day treatment with 40 μM celecoxib (Figure 4D). These results suggest that celecoxib induces apoptosis in MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants regardless of Bcl-2 expression.
Celecoxib enhances apoptosis of MDA-MB-231-E1A and MDA-MB-435-E1A cells via prostaglandins E2 or F2α
Given our findings that celecoxib induced apoptosis in the E1A stable transfectants and that COX-2 downregulation is involved in this apoptosis but Bcl-2 suppression is not, we next explored whether the celecoxib-induced apoptosis in these cells depends on a pathway downstream of COX-2. For these experiments, we tested the effects of a 5-day treatment with 40 μM celecoxib on cell viability with or without the addition of 10 μM prostaglandin (PG) E2 or PGF2α, two molecules located downstream of COX-2. In the MDA-MB-231-E1A cells, treatment with celecoxib alone produced a mean of 35.9% viable cells (± 2.7% SD); the addition of either prostaglandin substantially improved cell viability (62.1% ± 3.9% viable cells for PGE2 (P = 0.0005), 63.0% ± 3.8% for PGF2α (P = 0.0026). Results were similar for the MDA-MB-435-E1A cells (33.3% ± 7.5% for celecoxib only, 58.6% ± 7.7% for PGE2 (P = 0.041), and 60.2% ± 6.7% for PGF2α (P = 0.0030)) (Figure 5). These results suggest that celecoxib enhances apoptosis of cells that stably express E1A in part by blocking PGE2 or PGF2α.

Celecoxib-induced apoptosis of MDA-MB-231-E1A and MDA-MB-435-E1A cells depends on PGE2 or PGF2α. Treatment of MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants with 0 or 40 μM celecoxib (CLX) plus 10 μM of either PGE2or PGF2α for 5 days blunted sensitivity to celecoxib in both cell lines.
To test the effect of celecoxib on prostaglandin synthesis, we assessed PGE2 and PGF2α levels in MDA-MB-231-E1A and MDA-MB-435-E1A cells treated for 5 days with 40 μM celecoxib (data not shown). Enzyme-linked immunosorbent assay showed that in the MDA-MB-231-E1A cells, celecoxib treatment significantly inhibited PGE2 (mean ± SD 0.20 pg/ml ± 0.08 pg/ml vs 2.16 pg/ml ± 1.25 pg/ml for dimethylsulfoxide control; P = 0.002), but celecoxib had no effect on PGE2 synthesis in the MDA-MB-435-E1A cells. Conversely, celecoxib inhibited PGF2α in MDA-MB-435-E1A cells (121.7 pg/ml ± 12.3 pg/ml vs 173.6 pg/ml ± 7.4 pg/ml for control, P = 0.003) but did not affect PGF2α synthesis in MDA-MB-231-E1A cells.
Discussion
Our hypothesis was that celecoxib would enhance E1A-induced apoptosis by suppressing COX-2 expression and thereby suppressing Bcl-2 expression. In exploring the possibility that the COX-2 inhibitor celecoxib would downregulate Bcl-2, we found that celecoxib did enhance E1A-induced apoptosis in cells that express high levels of COX-2 protein. We also found that PGE2 or PGF2α are involved in this apoptotic pathway. However, celecoxib-induced apoptosis did not depend on suppression of Bcl-2.
With regard to suppression of Bcl-2 by COX-2 inhibitors, some previous studies have shown that the ability of COX-2 inhibitors to induce apoptosis in cancer cells depends on the downregulation of Bcl-2 [19–23]. However, others have reported that COX-2 inhibitors can induce apoptosis in cancer cells independently of Bcl-2 [29, 32]. Cao and Prescott [33] proposed that Bcl-2 overexpression is probably caused by reductions in arachidonic acid and increases in PGE2 levels. Nevertheless, our finding that COX-2 suppression did not suppress Bcl-2 leads us to propose that other mechanisms exist by which celecoxib induces apoptosis, at least in the breast cancer cell lines we tested. In other words, it is unclear how much suppression of Bcl-2 is enough to induce significant apoptosis, or, indeed, if celecoxib has the capacity to downregulate Bcl-2. It is possible that only the reduction in Bcl-2 led to apoptosis in the MDA-MB-435-E1A stable transfectants; however, that reduction would have to have been substantial. We showed that 40 μM of celecoxib was enough to enhance E1A-induced apoptosis in MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants regardless of Bcl-2 suppression. This result suggests that other molecules than Bcl-2 could be critical for celecoxib to enhance E1A-induced apoptosis.
We then investigated the involvement of PGE2 or PGF2α, two molecules located downstream of COX-2. The synthesis of prostaglandins is known to depend on COX-2 activity. In one study, the synthesis of PGE2 or PGF2α was enhanced by the overexpression of COX-2 in the mammary glands of COX-2 transgenic mice [34]. In addition, PGF2α has been shown to promote tumorigenesis in endometrial cancer cells [35]. Several reports indicate that the reduction of COX-2 inhibits the release of prostaglandins. For example, treatment of the COX-2-overexpressing myeloma cell line ARH-77 with indomethacin led to the reduction of PGD2, PGE2, and PGF2α [36]. Celecoxib has been shown to inhibit the release of PGE2 or PGF2α from ureteral segments in swine [37]. NS-398 inhibited the production of prostaglandins, including PGD2, PGE2 and PGF2α, in the prostate cancer cell line PC-3 [38]. We speculate that both PGE2 and PGF2α derived from COX-2 could be key factors in modulating the apoptotic effect in E1A-transfected cell lines. Indeed, we found that blocking PGE2 and PGF2α production was crucial for celecoxib-induced apoptosis in the E1A stable transfectants. We confirmed here that both PGE2 and PGF2α influenced apoptosis, but celecoxib suppressed different prostaglandins in the two different cell lines. This difference could reflect the involvement of other pathways that modulate apoptosis in E1A-transfected cell lines.
Davis et al. [39] reported that the COX-2 inhibitor NS-398 was more cytotoxic in a prostate epithelial cell line in which E2F1 had been activated than in the original prostate epithelial cell lines, speculating that the reason for the greater cytotoxicity was a disruption in the retinoblastoma/E2F complexes. We showed that caspase-9 and PARP were activated, but caspase-8 was not activated, in celecoxib-treated MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants (Figure 3B). These findings are consistent with others showing that celecoxib enhances caspase-3 or caspase-9 activation through PGE2 inhibition [42, 43]. We speculate that PGE2 or PGF2α (or both) inhibit intrinsic apoptotic pathway induced by E1A. This speculation is, to the best of our knowledge, the first to link PGE2 or PGF2α in the apoptosis associated with E1A.
Generally speaking, COX-2 inhibitors, including celecoxib, are thought to block prostaglandin synthesis by inhibiting the enzymatic activity of COX-2. However, COX-2 inhibitors (including celecoxib) might also act by suppressing production of the COX-2 protein [31, 44, 45]. We showed that the celecoxib-sensitive MDA-MB-231-E1A and MDA-MB-435-E1A stable transfectants originally overexpressed COX-2, and they produced less COX-2 protein in the presence of celecoxib (Figures 1 and 4A). Our finding that COX-2 levels were highest in the MDA-MB-231-E1A cells was confirmed by others' reports that the MDA-MB-231 cell line expresses high levels of COX-2 [46–48]. Moreover, our findings regarding sensitivity to celecoxib imply that celecoxib inhibited the production of prostaglandins not only by inhibiting COX-2 enzymatic activity but also by modulating COX-2 protein expression. A previous report indicated that modulating COX-2 protein expression with a structural analog of celecoxib (sc-236) or with the pharmacologic COX-2 protein-suppressing agent curcumin affected apoptosis in the COX-2-positive colon cancer cell line HT-29 [49]; another report showed that modulating COX-2 protein expression with the retinoid X receptor-selective retinoid LGD1069 decreased PGE2 production in normal human mammary epithelial cells [50]. We speculate that the modulation of COX-2 protein expression by celecoxib is a significant part of its effect in reducing prostaglandin levels.
Our study had some limitations. For one thing, although we did confirm that all four E1A stable transfectants expressed E1A (Figure 1), the level of expression was not uniform among the cell lines. Specifically, MDA-MB-231-E1A cells expressed slightly more E1A than the other three cell lines, a result that has also been shown by others [15, 28]. Thus, we were not able to exclude the possibility that E1A expression level could affect sensitivity to celecoxib. In addition, we examined only one COX-2 inhibitor, celecoxib. COX-2 inhibitors other than celecoxib can downregulate Bcl-2 [19, 22]; whether other COX-2 inhibitors would downregulate Bcl-2 expression to a greater extent than celecoxib remains to be seen. Elucidation of the precise mechanism by which COX-2 inhibitors downregulate Bcl-2 will help to clarify the appropriate uses of COX-2 inhibitors in preclinical settings for treating cancer.
Conclusion
We found that celecoxib enhanced E1A-induced apoptosis in breast cancer cells that express high levels of COX-2 protein and that this effect depended, at least in part, on blocking the production of PGE2 or PGF2α. Further studies exploring the precise mechanism by which prostaglandins influence E1A-induced apoptosis, and confirmation of synergistic effects between E1A gene therapy and COX-2 inhibitor treatment in cancer xenograft models, are needed to establish conclusively that COX-2 inhibitors can overcome resistance to E1A-induced apoptosis.
Abbreviations
- COX-2:
-
cyclooxygenase-2
- DMEM/F12:
-
Dulbecco's modified Eagle's medium/Ham's F-12 medium
- E1A :
-
adenovirus type 5 gene E1A
- MTT = 3-(4:
-
5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- PARP:
-
poly ADP-ribosome polymerase
- PBS:
-
phosphate-buffered saline
- PGE2 :
-
prostaglandin E2
- PGF2α :
-
prostaglandin F2α
- TNF-α:
-
tumor necrosis factor-α
- TRAIL:
-
TNF-related apoptosis-inducing ligand.
References
Xing X, Yujiao Chang J, Hung M: Preclinical and clinical study of HER-2/neu-targeting cancer gene therapy. Adv Drug Deliv Rev. 1998, 30: 219-227. 10.1016/S0169-409X(97)00118-X.
Hortobagyi GN, Ueno NT, Xia W, Zhang S, Wolf JK, Putnam JB, Weiden PL, Willey JS, Carey M, Branham DL, et al: Cationic liposome-mediated E1A gene transfer to human breast and ovarian cancer cells and its biologic effects: a phase I clinical trial. J Clin Oncol. 2001, 19: 3422-3433.
Villaret D, Glisson B, Kenady D, Hanna E, Carey M, Gleich L, Yoo GH, Futran N, Hung MC, Anklesaria P, et al: A multicenter phase II study of tgDCC-E1A for the intratumoral treatment of patients with recurrent head and neck squamous cell carcinoma. Head Neck. 2002, 24: 661-669. 10.1002/hed.10107.
Frisch SM, Mymryk JS: Adenovirus-5 E1A: paradox and paradigm. Nat Rev Mol Cell Biol. 2002, 3: 441-452. 10.1038/nrm827.
Berk AJ: Recent lessons in gene expression, cell cycle control, and cell biology from adenovirus. Oncogene. 2005, 24: 7673-7685. 10.1038/sj.onc.1209040.
Gomez-Manzano C, Yung WK, Alemany R, Fueyo J: Genetically modified adenoviruses against gliomas: from bench to bedside. Neurology. 2004, 63: 418-426.
Trimarchi JM, Lees JA: Sibling rivalry in the E2F family. Nat Rev Mol Cell Biol. 2002, 3: 11-20. 10.1038/nrm714.
Evan GI, Vousden KH: Proliferation, cell cycle and apoptosis in cancer. Nature. 2001, 411: 342-348. 10.1038/35077213.
Wu X, Levine AJ: p53 and E2F-1 cooperate to mediate apoptosis. Proc Natl Acad Sci USA. 1994, 91: 3602-3606. 10.1073/pnas.91.9.3602.
Teodoro JG, Shore GC, Branton PE: Adenovirus E1A proteins induce apoptosis by both p53-dependent and p53-independent mechanisms. Oncogene. 1995, 11: 467-474.
Chen MJ, Holskin B, Strickler J, Gorniak J, Clark MA, Johnson PJ, Mitcho M, Shalloway D: Induction by E1A oncogene expression of cellular susceptibility to lysis by TNF. Nature. 1987, 330: 581-583. 10.1038/330581a0.
Duerksen-Hughes P, Wold WS, Gooding LR: Adenovirus E1A renders infected cells sensitive to cytolysis by tumor necrosis factor. J Immunol. 1989, 143: 4193-4200.
Routes JM, Ryan S, Clase A, Miura T, Kuhl A, Potter TA, Cook JL: Adenovirus E1A oncogene expression in tumor cells enhances killing by TNF-related apoptosis-inducing ligand (TRAIL). J Immunol. 2000, 165: 4522-4527.
Shao R, Lee DF, Wen Y, Ding Y, Xia W, Ping B, Yagita H, Spohn B, Hung MC: E1A sensitizes cancer cells to TRAIL-induced apoptosis through enhancement of caspase activation. Mol Cancer Res. 2005, 3: 219-226.
Liao Y, Hung MC: Regulation of the activity of p38 mitogen-activated protein kinase by Akt in cancer and adenoviral protein E1A-mediated sensitization to apoptosis. Mol Cell Biol. 2003, 23: 6836-6848. 10.1128/MCB.23.19.6836-6848.2003.
Shao R, Karunagaran D, Zhou BP, Li K, Lo SS, Deng J, Chiao P, Hung MC: Inhibition of nuclear factor-kappaB activity is involved in E1A-mediated sensitization of radiation-induced apoptosis. J Biol Chem. 1997, 272: 32739-32742. 10.1074/jbc.272.52.32739.
Lee WP, Liao Y, Robinson D, Kung HJ, Liu ET, Hung MC: Axl-gas6 interaction counteracts E1A-mediated cell growth suppression and proapoptotic activity. Mol Cell Biol. 1999, 19: 8075-8082.
Bartholomeusz C, Itamochi H, Yuan LX, Esteva FJ, Wood CG, Terakawa N, Hung MC, Ueno NT: Bcl-2 antisense oligonucleotide overcomes resistance to E1A gene therapy in a low HER2-expressing ovarian cancer xenograft model. Cancer Res. 2005, 65: 8406-8413. 10.1158/0008-5472.CAN-05-1754.
Liu XH, Yao S, Kirschenbaum A, Levine AC: NS398, a selective cyclooxygenase-2 inhibitor, induces apoptosis and down-regulates bcl-2 expression in LNCaP cells. Cancer Res. 1998, 58: 4245-4249.
Subhashini J, Mahipal SV, Reddanna P: Anti-proliferative and apoptotic effects of celecoxib on human chronic myeloid leukemia in vitro. Cancer Lett. 2005, 224: 31-43.
Catalano A, Graciotti L, Rinaldi L, Raffaelli G, Rodilossi S, Betta P, Gianni W, Amoroso S, Procopio A: Preclinical evaluation of the nonsteroidal anti-inflammatory agent celecoxib on malignant mesothelioma chemoprevention. Int J Cancer. 2004, 109: 322-328. 10.1002/ijc.11710.
Li XH, Li JJ, Zhang HW, Sun P, Zhang YL, Cai SH, Ren XD: Nimesulide inhibits tumor growth in mice implanted hepatoma: overexpression of Bax over Bcl-2. Acta Pharmacol Sin. 2003, 24: 1045-1050.
Basu GD, Pathangey LB, Tinder TL, Lagioia M, Gendler SJ, Mukherjee P: Cyclooxygenase-2 inhibitor induces apoptosis in breast cancer cells in an in vivo model of spontaneous metastatic breast cancer. Mol Cancer Res. 2004, 2: 632-642.
Pereg D, Lishner M: Non-steroidal anti-inflammatory drugs for the prevention and treatment of cancer. J Intern Med. 2005, 258: 115-123. 10.1111/j.1365-2796.2005.01519.x.
Tsujii M, DuBois RN: Alterations in cellular adhesion and apoptosis in epithelial cells overexpressing prostaglandin endoperoxide synthase 2. Cell. 1995, 83: 493-501. 10.1016/0092-8674(95)90127-2.
Sheng H, Shao J, Morrow JD, Beauchamp RD, DuBois RN: Modulation of apoptosis and Bcl-2 expression by prostaglandin E2 in human colon cancer cells. Cancer Res. 1998, 58: 362-366.
Shao R, Hu MC, Zhou BP, Lin SY, Chiao PJ, von Lindern RH, Spohn B, Hung MC: E1A sensitizes cells to tumor necrosis factor-induced apoptosis through inhibition of IkappaB kinases and nuclear factor kappaB activities. J Biol Chem. 1999, 274: 21495-21498. 10.1074/jbc.274.31.21495.
Meric F, Liao Y, Lee WP, Pollock RE, Hung MC: Adenovirus 5 early region 1A does not induce expression of the ewing sarcoma fusion product EWS-FLI1 in breast and ovarian cancer cell lines. Clin Cancer Res. 2000, 6: 3832-3836.
Kern MA, Schubert D, Sahi D, Schoneweiss MM, Moll I, Haugg AM, Dienes HP, Breuhahn K, Schirmacher P: Proapoptotic and antiproliferative potential of selective cyclooxygenase-2 inhibitors in human liver tumor cells. Hepatology. 2002, 36: 885-894.
Zou Y, Peng H, Zhou B, Wen Y, Wang SC, Tsai EM, Hung MC: Systemic tumor suppression by the proapoptotic gene bik. Cancer Res. 2002, 62: 8-12.
Tuynman JB, Peppelenbosch MP, Richel DJ: COX-2 inhibition as a tool to treat and prevent colorectal cancer. Crit Rev Oncol Hematol. 2004, 52: 81-101. 10.1016/S1040-8428(04)00139-8.
Hsu AL, Ching TT, Wang DS, Song X, Rangnekar VM, Chen CS: The cyclooxygenase-2 inhibitor celecoxib induces apoptosis by blocking Akt activation in human prostate cancer cells independently of Bcl-2. J Biol Chem. 2000, 275: 11397-11403. 10.1074/jbc.275.15.11397.
Cao Y, Prescott SM: Many actions of cyclooxygenase-2 in cellular dynamics and in cancer. J Cell Physiol. 2002, 190: 279-286. 10.1002/jcp.10068.
Liu CH, Chang SH, Narko K, Trifan OC, Wu MT, Smith E, Haudenschild C, Lane TF, Hla T: Overexpression of cyclooxygenase-2 is sufficient to induce tumorigenesis in transgenic mice. J Biol Chem. 2001, 276: 18563-18569. 10.1074/jbc.M010787200.
Jabbour HN, Sales KJ, Boddy SC, Anderson RA, Williams AR: A positive feedback loop that regulates cyclooxygenase-2 expression and prostaglandin F2α synthesis via the F-series-prostanoid receptor and extracellular signal-regulated kinase 1/2 signaling pathway. Endocrinology. 2005, 146: 4657-4664. 10.1210/en.2005-0804.
Ding J, Tsuboi K, Hoshikawa H, Goto R, Mori N, Katsukawa M, Hirai E, Yamamoto S, Abe M, Ueda N: Cyclooxygenase isozymes are expressed in human myeloma cells but not involved in anti-proliferative effect of cyclooxygenase inhibitors. Mol Carcino. 2006, 45: 250-259. 10.1002/mc.20175.
Jerde TJ, Calamon-Dixon JL, Bjorling DE, Nakada SY: Celecoxib inhibits ureteral contractility and prostanoid release. Urology. 2005, 65: 185-190. 10.1016/j.urology.2004.08.057.
Nithipatikom K, Isbell MA, Lindholm PF, Kajdacsy-Balla A, Kaul S, Campell WB: Requirement of cyclooxygenase-2 expression and prostaglandins for human prostate cancer cell invasion. Clin Exp Metastasis. 2002, 19: 593-601. 10.1023/A:1020915914376.
Davis JN, McCabe MT, Hayward SW, Park JM, Day ML: Disruption of Rb/E2F pathway results in increased cyclooxygenase-2 expression and activity in prostate epithelial cells. Cancer Res. 2005, 65: 3633-3642. 10.1158/0008-5472.CAN-04-3129.
Nevins JR: The Rb/E2F pathway and cancer. Hum Mol Genet. 2001, 10: 699-703. 10.1093/hmg/10.7.699.
Tsantoulis PK, Gorgoulis VG: Involvement of E2F transcription factor family in cancer. Eur J Cancer. 2005, 41: 2403-2414. 10.1016/j.ejca.2005.08.005.
Dandekar DS, Lopez M, Carey RI, Lokeshwar BL: Cyclooxygenase-2 inhibitor celecoxib augments chemotherapeutic drug-induced apoptosis by enhancing activation of caspase-3 and -9 in prostate cancer cells. Int J Cancer. 2005, 115: 484-492. 10.1002/ijc.20878.
Zhang Z, Lai GH, Sirica AE: Celecoxib-induced apoptosis in rat cholangiocarcinoma cells mediated by Akt inactivation and Bax translocation. Hepatology. 2004, 39: 1028-1037. 10.1002/hep.20143.
Narayanan BA, Condon MS, Bosland MC, Narayanan NK, Reddy BS: Suppression of N-methyl-N-nitrosourea/testosterone-induced rat prostate cancer growth by celecoxib: effects on cyclooxygenase-2, cell cycle regulation, and apoptosis mechanism(s). Clin Cancer Res. 2003, 9: 3503-3513.
Yao M, Kargman S, Lam EC, Kelly CR, Zheng Y, Luk P, Kwong E, Evans JF, Wolfe MM: Inhibition of cyclooxygenase-2 by rofecoxib attenuates the growth and metastatic potential of colorectal carcinoma in mice. Cancer Res. 2003, 63: 586-592.
Basu GD, Pathangey LB, Tinder TL, Gendler SJ, Mukherjee P: Mechanisms underlying the growth inhibitory effects of the cyclo-oxygenase-2 inhibitor celecoxib in human breast cancer cells. Breast Cancer Res. 2005, 7: 422-435. 10.1186/bcr1019.
Liu XH, Rose DP: Differential expression and regulation of cyclooxygenase-1 and -2 in two human breast cancer cell lines. Cancer Res. 1996, 56: 5125-5127.
Timoshenko AV, Xu G, Chakrabarti S, Lala PK, Chakraborty C: Role of prostaglandin E2 receptors in migration of murine and human breast cancer cells. Exp Cell Res. 2003, 289: 265-274. 10.1016/S0014-4827(03)00269-6.
Agarwal B, Swaroop P, Protiva P, Raj SV, Shirin H, Holt PR: Cox-2 is needed but not sufficient for apoptosis induced by Cox-2 selective inhibitors in colon cancer cells. Apoptosis. 2003, 8: 649-654. 10.1023/A:1026199929747.
Kong G, Kim HT, Wu K, DeNardo D, Hilsenbeck SG, Xu XC, Lamph WW, Bissonnette R, Dannenberg AJ, Brown PH: The retinoid X receptor-selective retinoid, LGD down-regulates cyclooxygenase-2 expression in human breast cells through transcription factor crosstalk: implications for molecular-based chemoprevention. Cancer Res. 1069, 65: 3462-3469.
Acknowledgements
We thank Dr Uma Raju for technical support and reagents, Dr Mien-Chie Hung for the E1A transfectants and assistance with the Bcl-2 transfections, and Christine F Wogan at the Department of Scientific Publications at M D Anderson Cancer Center for editorial suggestions. No grant funding supported this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
TS participated in conceiving and designing the study, carried out many of the experiments, and drafted the manuscript. CB participated in the collection, analysis, and interpretation of findings, provided procedural guidance and expertise, and participated in revising the manuscript. AMT, an expert in COX and COX inhibition in cancer, conceived the prostaglandin experiments. NTU led the conception and design of the study, the analysis and interpretation of the findings, and the revisions to the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Sugimoto, T., Bartholomeusz, C., Tari, A.M. et al. Adenovirus type 5 E1A-induced apoptosis in COX-2-overexpressing breast cancer cells. Breast Cancer Res 9, R41 (2007). https://doi.org/10.1186/bcr1739
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/bcr1739