
Abstract
To evaluate the antioxidant activity of the glycosaminoglycans hyaluronic acid (HYA) and chondroitin-4-sulphate (C4S), we used a rat model of collagen-induced arthritis (CIA). Arthritis was induced in Lewis rats by multiple intradermal injections of 250 μl of emulsion containing bovine type II collagen in complete Freund's adjuvant at the base of the tail and into three to five other sites on the back. Rats were challenged again with the same antigen preparation 7 days later. Disease developed about 11 days after the second immunization. The effects of treatment in the rats were monitored by biochemical parameters and by macroscopic and histological evaluations in blood, synovial tissue and articular cartilage. Arthritis produced the following symptoms: severe periarticular erythema, edema and inflammation in the hindpaws; membrane peroxidation in the cartilage of the joints; endogenous antioxidant wasting; high tumour necrosis factor-α (TNF-α) plasma levels; and synovial neutrophil accumulation. Treatment with HYA and C4S, starting at the onset of arthritis for 10 days, limited the erosive action of the disease in the articular joints of knee and paw, reduced lipid peroxidation, restored the endogenous antioxidants reduced glutathione (GSH) and superoxide dismutase, decreased plasma TNF-α levels, and limited synovial neutrophil infiltration. These data confirm that erosive destruction of the joint cartilage in CIA is due at least in part to free radicals released by activated neutrophils and produced by other biochemical pathways. The beneficial effects obtained with the treatment suggest that HYA and C4S could be considered natural endogenous macromolecules to limit erosive damage in CIA or as a useful tool with which to study the involvement of free radicals in rheumatoid arthritis.
Similar content being viewed by others


Introduction
Rheumatoid arthritis (RA) is a common human autoimmune disease characterised by chronic inflammation of the synovial joints and by subsequent progressive, erosive destruction of articular tissue [1]. This disease affects about 1% of the human population. The aetiology and pathogenesis of this disease are not yet fully understood but it seems likely that an autoimmune-mediated attack on the joints has a crucial role in the pathogenesis of RA [2].
Collagen-induced arthritis (CIA) in Lewis rats is a widely used experimental animal model of inflammatory polyarthritis with clinical and pathological features similar to those of human RA that are dependent on both humoral and cellular immunity to the immunising antigen [3].
It has been suggested previously that the generation of free radicals and other reactive oxygen species (ROS) such as singlet oxygen and hypochlorous acid might have a role in the pathogenesis of degenerative joint disease [4]. ROS are highly reactive transient chemical species with the potential to initiate cellular damage in joint tissues. These reactive molecules are formed during normal aerobic metabolism in cells and after the activation of phagocytes during infection or inflammation; a consequence of the uncontrolled production of free radicals is damage to biomolecules leading to altered function and disease [5]. There are many pieces of evidence, both direct and indirect, implicating radicals in the pathogenesis of inflammatory synovitis, such as the capacity of several cells that are present in the inflamed joint (macrophages, neutrophils, lymphocytes and endothelial cells) to produce free radicals when isolated and stimulated [6]. Cells are normally protected from ROS-induced damage by a variety of endogenous scavenging proteins, enzymes and chemical compounds that constitute the endogenous antioxidant systems [7]. It has been reported that ROS destroy antioxidant systems (in fact the enzymatic and/or non-enzymatic antioxidant systems are impaired in RA) and that RA patients are thus exposed to oxidant stress and lipid peroxidation because of the reduced antioxidant defence system [8, 9].
Glycosaminoglycans (GAGs), a large family of heterogeneous polysaccharides, are linear sulphate-substituted polymers composed of alternating hexuronic acid and hexosamine units that are important in all living organisms [10]. Their structure and degree of heterogeneity seem to be highly specific; the ability of several proteins to bind GAGs might reflect functional relationships and is likely to be exploited physiologically in a variety of ways. Several reports have shown that during the progression of RA the physiological levels of blood GAGs are increased [11–13]. The obvious explanation is that GAGs originate from the metabolism of the joint cartilage damaged by erosion. Nevertheless, the exact meaning of their increase is still unclear.
Molecules able to limit the generation and the effects of ROS exert a protective action in a variety of experimental inflammatory diseases, including CIA [14–17]. Many investigators have described the antioxidant properties of some GAGs (mainly for hyaluronic acid [HYA] and chondroitin-4-sulphate [C4S]) in experimental models, both in vitro and in vivo [18–22].
Starting from these findings, the aim of the present study was to assess the possible ability of HYA and C4S in limiting inflammation and joint cartilage erosion in an experimental model of CIA in Lewis rats.
Materials and methods
Animals
Male Lewis rats 6–7 weeks old, with a mean weight of 175–200 g, were used in our study. Rats, purchased from Charles River (Calco, Italy), were maintained under climate-controlled conditions in a 12 hours light : 12 hours dark cycle. The animals were fed with standard rodent chow and water ad libitum. The health status of the animal colony was monitored in accordance with the guidelines from the Italian Veterinary Board. Rats were subdivided into the following groups: (1) control (n = 7); (2) CIA plus vehicle (n = 7); (3) CIA plus HYA (25 mg/kg) (n = 7); (4) CIA plus C4S (25 mg/kg) (n = 7); (5) CIA plus HYA plus C4S (each at 25 mg/kg) (n = 7).
Materials
Hyaluronic acid from human umbilical cord, chondroitin sulphate A from bovine trachea and bovine type II collagen were purchased from Sigma-Aldrich Srl (Milan, Italy); complete Freund's adjuvant was obtained from Difco Laboratories (Detroit, MI, USA). All other reagents were purchased from Fluka (division of Sigma-Aldrich Srl, Milan, Italy).
Induction of arthritis
CIA was induced in rats as described previously [23], by multiple intradermal injections, at the base of the tail and into three to five other sites on the back, of 250 μg of bovine type II collagen in 125 μl of 0.1 M acetic acid emulsified in an equal volume of complete Freund's adjuvant containing 2 mg/ml Mycobacterium tuberculosis H37 RA [23]. Rats were challenged again with the same antigen preparation 7 days later. Before injection, animals were anaesthetised with ether and injections were performed with a 15 gauge needle. Disease developed about 11 days after the second immunisation.
Treatment with GAGs
At day 11, animals were randomised to receive treatments listed in the Animals section, timed to coincide approximately with the onset of arthritis pathology. GAGs were dissolved in physiological saline (0.9% NaCl) and administered daily, in a volume of 1 ml/kg body weight, intraperitoneally once a day until the 20th day.
Plasma GAG evaluation
Evaluation of plasma galactosamine and glucosamine was performed at day 21 to estimate indirectly the concentration of HYA and C4S in the blood of animals after the intraperitoneal treatment. Samples of blood (1.5 ml) were drawn, at the end of the experiment, from a tail vessel. The blood was collected in polyethylene tubes with the previous addition of 75 μl of heparin solution (4000 IU). The plasma samples obtained after centrifugation for 10 min at 3000 g and 4°C were frozen at -80°C until assay. On the day of analysis, GAGs were first isolated and purified and then hydrolysed into their constituent monosaccharides [24]. These amino sugars (glucosamine and galactosamine) were then assayed by a specific HPLC method [24].
Arthritis assessments
Evaluation of joint inflammation was performed by a blinded independent observer with no knowledge of the treatment protocol. The severity of the arthritis in each paw was quantified daily by a clinical score measurement [25] from 0 to 4 as follows: 0, no macroscopic signs of arthritis; 1, swelling of one group of joints (namely, wrist or ankle joints); 2, two groups of swollen joints; 3, three groups of swollen joints; 4; swelling of the entire paw. The maximum score for each rat was 16. Clinical severity was also assessed by the quantification of the paw volume changes. Measurements were performed with a dial gauge caliper. Changes in body weight were monitored to determine the rate of the increment in each rat.
Histological analysis
Rats were killed at day 21 by ether narcosis; hind limbs were removed and fixed in 10% buffered formalin. The limbs were decalcified in 5% formic acid, processed for paraffin embedding, sectioned at 5 μm thickness, and subsequently stained with haematoxylin-eosin for examination under a light microscope [26]. Sections were examined for the presence of hyperplasia of the synovium, pannus formation, and destruction of the joint space.
Lipid peroxidation analysis
Determination of malonaldehyde in the articular tissue was performed to estimate the extent of lipid peroxidation in the damaged cartilage. At the end of the experiment hind limbs were removed and maintained at 0°C, then the joint cartilage was quickly separated from the bone and muscular tissue and frozen at -80°C until assay. On the day of analysis, after thawing, cartilage samples were washed in ice-cold 20 mM Tris-HCl, pH 7.4, and blotted on absorbent paper. Each sample was then minced in ice-cold 20 mM Tris-HCl, pH 7.4 containing 1 mg/ml butylated hydroxytoluene and homogenised in a 1 : 10 (w/v) ratio with an Ultra-Turrax homogeniser. After centrifugation for 10 min at 3000 g and 4°C, the clear supernatant was used for biochemical assay. Analysis was performed with a colorimetric commercial kit (Lipid peroxidation assay kit, cat. no. 437634; Calbiochem, La Jolla, CA, USA). In brief, 0.65 ml of 10.3 mM N-methyl-2-phenylindole in acetonitrile was added to 0.2 ml of homogenate supernatant. After vortex-mixing for 3–4 s and the addition of 0.15 ml of 37% HCl, samples were well mixed, closed with a tight stopper and incubated for 60 min at 45°C. The samples were then cooled on ice and the absorbance was measured spectrophotometrically at 586 nm. A calibration curve of an accurately prepared standard malonaldehyde solution was also run for quantification. The malonaldehyde concentration was expressed as nmol/mg of protein.
Determination of GSH
Samples of joint cartilage obtained at the end of the experiment were frozen at -80°C until GSH assay. The analysis was performed with a spectrophotometric method [27]. In brief, tissue samples were homogenised in a solution containing 5% trichloroacetic acid and 5 mM EDTA at 4°C. Then each sample was centrifuged for 10 min at 15,000 g and 4°C. An aliquot of homogenate supernatant (0.4 ml) was added to a dark polyethylene tube containing 1.6 ml of 0.4 M Tris-EDTA buffer, pH 8.9. After vortex-mixing, 40 μl of 10 mM dithiobisnitrobenzoic acid in methanol was added. The samples were vortex-mixed again and the absorbance at 412 nm was read after 5 min. The values of unknown samples were drawn from a standard curve plotted by assaying different known concentrations of GSH. The amount of GSH was expressed as μmol/g of protein.
Evaluation of superoxide dismutase (SOD)
Samples of joint cartilage were washed with 0.9% NaCl containing 0.16 mg/ml heparin and homogenised with a Ultra-Turrax homogeniser in ice-cold 0.25 M sucrose containing 1 mM diethylenetriamine pentaacetic acid (1 : 1, w/v). Each sample was then centrifuged for 20 min at 20,000 g and 4°C. The supernatant was aspirated and the total SOD activity was assayed spectrophotometrically at 505 nm with a commercial kit (Ransod assay kit, cat. no. Sd 125; Randox Laboratories, Crumlin, UK). In brief, 50 μl of each diluted sample (1 : 10 [w/v] in 0.01 M potassium phosphate buffer, pH 7.0) was mixed with 1.7 ml of solution containing 0.05 mM xanthine and 0.025 mM iodonitrotetra-zolium chloride. After being mixed for 5 s, 250 μl of xanthine oxidase (80 U/l) was added. The time between reading the initial and final absorbances was 3 min. A standard curve of SOD solution (from 2 to 32 U/ml) was run for quantification. All standard and diluted sample rates were converted into a percentage of the buffer diluent rate and subtracted from 100% to give a percentage inhibition. Sample SOD activities were obtained from a plotted curve of the percentage inhibition for each standard. SOD values were expressed as units/mg of protein.
Plasma tumour necrosis factor-α (TNF-α) assay
Plasma TNF-α concentration was determined with an ELISA commercial kit (Rat TNF-α ELISA kit ultra sensitive, cat. no. 22079; NBS Biologicals Ltd, Huntingdon, UK). At the end of the experiment, samples of blood (0.5 ml) were drawn from a tail vessel. The blood was collected in polyethylene tubes with the previous addition of 25 μl of heparin solution (4000 IU). The plasma samples obtained after centrifugation for 10 min at 3000 g and 4°C were frozen at -80°C until assay. In brief, 100 μl of standards, samples and controls were added to each well of the coated microplate. After 3 hours of incubation at 24°C the microplate was decanted and the liquid discarded. Then, 100 μl of biotinylated anti-TNF-α antibody was added to each well. After 45 min of incubation at 24°C and a further elimination of the liquid from the wells, 100 μl of Streptavidin-horseradish peroxidase conjugate was added. After incubation for a further 45 min and a washing of the wells, 100 μl of chromogen were added. The absorbance of each well was read spectrophotometrically at 450 nm. TNF-α values were expressed as ng/ml.
Articular neutrophil accumulation
Myeloperoxidase activity was analysed as an index of neutrophil infiltration in the synovial tissue, because it is closely correlated with the number of neutrophils present in the tissue [28]. We measured myeloperoxidase in the synovial tissue of joints by a specific assay for this enzyme [29]. In brief, synovial tissue samples were separated from rat joints and were first homogenised in a solution containing 20 mM potassium phosphate buffer, pH 7.0, to 1 : 10 (w/v) and then centrifuged for 30 min at 20,000 g and 4°C. The supernatant of each sample was discarded and the resulting pellet was added to a buffer solution consisting of 0.5% hexadecyltrimethylammonium bromide dissolved in 50 mM potassium phosphate buffer, pH 6, containing 50 μl of protease and phosphatase inhibitor cocktails. Samples were then sonicated for 1 min and centrifuged for 30 min at 20,000 g and 4°C. An aliquot of the supernatant was allowed to react with a solution of o-dianisidine dihydrochloride (0.167 mg/ml) and 0.0005% hydrogen peroxide. The rate of change in absorbance was measured spectrophotometrically at 405 nm. Myeloperoxidase activity has been defined as the concentration of enzyme degrading 1 μmol of peroxide/min at 37°C and was expressed as U/g of protein.
Statistical analysis
Data are expressed as means ± SD. The difference between the means of two groups was evaluated with an ANOVA and was considered significant at P < 0.05.
Statement of animal care
The studies reported in this manuscript have been performed in accordance with the declaration of Helsinki and with the Guide for the Care and Use of Laboratory Animals.
Results
Effects of HYA and C4S on clinical signs of CIA
Four days after the second immunisation, animals began to show evidence of clinical inflammation in one or more hindpaws. The first manifestation of disease was erythema of one or more ankle joints, followed by involvement of the metatarsal and interphalangeal joints. In Figure 1A we show the incidence of CIA through the 21-day study period. The initial signs of arthritis in all groups were evident at day 11 and the incidence was about 55%. In the vehicle group the incidence was 100% at day 14; the same incidence was maintained until the end of the experiment. The treatment with GAGs exerted a significant attenuation in the incidence of CIA: 70% in HYA treatment, 60% in C4S treatment and 42% in HYA plus C4S treatment (Fig. 1A).

Effect of hyaluronic acid (HYA) and chondroitin-4-sulphate (C4S) on the time course of the development and progression of arthritis. (A) Cumulative incidence of arthritis and day of onset of arthritis. (B) Arthritis severity scores in rats during 21 day period after immunisation. Values are means ± SD for seven animals for each group.
The typical time course of the development and progression of disease, as assessed by mean arthritis severity score and paw diameter, is shown in Figures 1B and 2A. By 11 days all animals showed evidence of disease, predominantly in the hindpaws. The disease was always progressive, with joint recruitment following the same pattern: tarsal, metatarsophalangeal and then interphalangeal. The interphalangeal joints were never solely involved, and inflammation in these joints was invariably associated with inflammation in the tarsal joint. The mean arthritis severity score in the CIA plus vehicle group was progressive from day 11 and achieved values of about 10 in the last four days (Fig. 1B). The same variations were observed in the hindpaw diameter of CIA rats given vehicle alone. In fact, at the end of the experiment, the increase in the hindpaw diameter due to oedema and inflammation was about 1 mm (Fig. 2A). Administration of HYA and C4S attenuated both the mean arthritis severity score and the change in hindpaw diameter (Figs 1B and 2A). Only the treatment with HYA reduced the hindpaw diameter but it was not statistically significant (Fig. 2A).

Effect of hyaluronic acid (HYA) and chondroitin-4-sulphate (C4S) on time course of change in paw diameter (A) and body weight increment (B) in rats with collagen-induced arthritis during the 21 days after immunisation. Values are means ± SD for seven animals for each group.
Changes in body weight
In the first 2 weeks the absolute increment in body weight was similar in all groups, and no significant differences were seen between them. After day 15 a significant loss in body weight was observed in the CIA rats given vehicle alone compared with the control rats. This weight loss increased until the end of the experiment (Fig. 2B). Treatment with GAGs ameliorated the decrease in body weight; the greatest effect was achieved after treatment with both polymers (Fig. 2B).
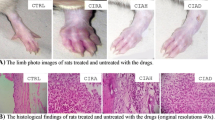
Histology
Representative joint histopathology of the experimental groups is shown in Figure 3. A characteristic of arthritic joints in rats with CIA is synovial hyperplasia, pannus formation, exudation of cells into the joint space, and erosion of bone and cartilage. A massive influx of inflammatory cells, synovial hyperplasia, and accumulation of abundant monomorphonuclear and polymorphonuclear cells in the joint space are evident (Fig. 3B) compared with a normal control group (Fig. 3A). By comparison, rats treated with HYA plus C4S revealed minimal evidence of inflammation or joint destruction. In fact the synovial membrane in the joints was like normal synovium, with few signs of synovial hyperplasia or other characteristics of inflammation (Fig. 3E). A reduced degree of arthritis severity was also observed in the rats that received only HYA or C4S (Fig. 3C,3D).

Representative joint histopathology of the groups with collagen-induced arthritis (CIA) administered vehicle alone and administered hyaluronic acid (HYA), chondroitin-4-sulphate (C4S) or both glycosaminoglycans, at the end of the experiment. (A) control; (B) CIA plus vehicle; (C) CIA plus HYA; (D) CIA plus C4S; (E) CIA plus HYA plus C4S. (Original magnification × 100.)
Plasma GAG levels
Because HYA contains the amino sugar glucosamine as a component, whereas C4S contains galactosamine, Table 1 reports the concentrations of GAGs, expressed in terms of glucosamine and galactosamine, in rats that underwent CIA after 10 days of treatment with HYA, C4S or both substances. In the control group, galactosamine levels were 9.81 ± 1.11 mg/l while glucosamine levels were 10.35 ± 1.71 mg/l; these values were considered physiological. However, in CIA rats treated with vehicle alone there was a significant increase in both amino sugars. The increase was even more significant in the three groups administered with HYA, C4S or both GAGs. The intraperitoneal administration of HYA increased glucosamine levels by about 31%; treatment with C4S increased galactosamine levels by about 30%. The administration of both GAGs increased both glucosamine and galactosamine (30% and 22% respectively; Table 1).
Assessment of malonaldehyde (MAL)
Determination of malonaldehyde in the articular cartilage was performed to estimate free-radical damage to biological membranes (Fig. 4). Low levels of malonaldehyde were seen in the control group at the end of the experiment (day 21); these values were considered normal. In contrast, a significant increase in malonaldehyde production was found in the joints of CIA rats given vehicle alone. Treatment with GAGs decreased malonaldehyde concentrations by inhibiting lipid peroxidation in the cartilage tissue. Treatment with HYA was at the limit of significance, whereas the maximum effect was observed by administering HYA plus C4S (Fig. 4).

Articular cartilage malonaldehyde (MAL) content of joints of animals with RA treated with hyaluronic acid (HYA), chondroitin-4-sulphate (C4S) or both glycosaminoglycans. Values are means ± SD for seven animals for each group.
GSH assay
The concentration of GSH was evaluated to estimate endogenous defences against hydrogen peroxide formation. Figure 5A shows the changes in GSH content evaluated in the joint articular cartilage (day 21) in the experimental groups. In normal control rats GSH levels ranged between 5.0 and 7.0 μmol/g of protein. In contrast, a marked decrease in GSH concentrations was found in the joint articular cartilage of CIA rats given vehicle alone. Treatment with each of the two polymers significantly inhibited the decrease in GSH levels. In this case, too, the maximum effect was obtained in the group treated with both polymers.

Articular cartilage antioxidant content of joints of animals with RA treated with hyaluronic acid (HYA), chondroitin-4-sulphate (C4S) or both glycosaminoglycans. (A) Articular reduced glutathione (GSH) levels. (B) Articular superoxide dismutase (SOD) activity. Values are means ± SD for seven animals for each group.
SOD activity
SOD activity was evaluated to estimate endogenous defences against superoxide anions. Figure 5B summarises the articular cartilage content of SOD in the experimental groups. In control animals normal SOD activities ranged between 10.0 and 15.0 U/mg of protein. In contrast, a significant decrease in this antioxidant was seen in CIA rats treated only with vehicle. As previously, chronic administration of the two GAGs significantly limited the decline in SOD.
TNF-α levels
Figure 6 reports the changes in TNF-α concentration assayed in plasma of rats at the end of the experiment. In the control group the normal levels of TNF-α ranged between 20.0 and 40.0 pg/ml. A marked increase in TNF-α concentration was found in the plasma of CIA rats given vehicle alone. Administration of GAGs significantly inhibited the increase in the cytokine in the three other groups.

Plasma TNF-α concentrations assayed in rats with RA treated with hyaluronic acid (HYA), chondroitin-4-sulphate (C4S) or both glycosaminoglycans. Values are means ± SD for seven animals for each group.
Myeloperoxidase analysis
Very low myeloperoxidase activity was measured in control rats (Fig. 7). In contrast, elevated myeloperoxidase levels were measured in the vehicle-administered CIA group. However, treatment with HYA and C4S decreased neutrophil accumulation by reducing myeloperoxidase activity in the synovial tissue of the joints. The decrease in myeloperoxidase activity was similar in all GAG-treated groups (Fig. 7).

Articular myeloperoxidase (MPO) content of joints of animals with RA treated with hyaluronic acid (HYA), chondroitin-4-sulphate (C4S) or both glycosaminoglycans. Values are means ± SD for seven animals for each group.
Discussion
Free radicals have long been implicated as mediators of tissue damage in RA patients [30]. Correspondingly, it has been shown that affected articulations are infiltrated by blood-derived cells, mainly neutrophils, macrophages and dendritic cells [31]. In response to activation, these cells are responsible for the generation of ROS [32, 33], which are released in large amounts into the surrounding tissue. When the endogenous antioxidant defences are overcome, the resulting production of free radicals induces impairment and destruction of the affected joint constituents such as synovial fluid, cartilage and other articular constituents [30]. One of several approaches to reduce oxidative stress is treatment with antioxidant compounds as therapeutic agents [34–38].
Acid GAGs are present in blood, usually in proteoglycan form. The main GAG of normal human plasma is C4S, which is mostly in a low-sulphate form [39, 40]. Keratan sulphate, heparan sulphate and HYA are the other GAG structures usually detected in human plasma [39, 40]. In animals, the total amounts of GAGs in plasma [41] are similar to those measured in humans [42]. Nevertheless, significantly increased plasma concentrations of GAGs have been observed in a wide range of diseases [39, 43–46] including RA in humans [47, 48] and experimental arthritis in rat and mouse [49, 50]. These changes in circulating GAGs in RA are thought to represent products of the connective tissue metabolism, and some circulating GAG structures are probably degradation products originating from articular cartilage [51]. However, the high levels of HYA and other GAGs found in RA [48] could not be explained only by the erosion of articular cartilage. In fact, the biological role, the real sites of origin and the metabolic fate of these amino-sugar-containing polysaccharides are not clearly understood.
Another approach to explaining the increased presence of GAG in the plasma comes from evidence that these molecules might function as carriers or modulators between adjacent cells [52]. It has been shown that GAGs can alter the binding with selectins because the latter bind several glycoproteins [53]. HYA and other GAGs could decrease cytokine gene expression directly [54] or indirectly by binding the CD44 receptor [55]. GAGs might also stimulate or inhibit cell proliferation in different mesothelioma cell lines [56]. In addition, some GAGs possess antioxidant activity capable of reducing free radicals and inhibiting lipid peroxidation [57–59]. The use of these molecules as therapeutic agents has shown some positive outcomes both in humans and in experimental animal models [36, 60–64]. The present study was designed to evaluate the effect of chronic treatment with HYA and C4S in a rat model of CIA. The choice of these two compounds from other GAGs was made on the basis of the evidence that they show the best antioxidant activity [36, 60–64].
The data obtained in the groups treated with HYA, C4S or both GAGs were positive in all parameters considered. Nevertheless, treatment with HYA alone showed less protection than that with C4S, and often the data were not significant (Fig. 2A,2B) or at the limit of significance (Figs 3C, 4, 5A,5B and 7). In contrast, the administration of both compounds showed maximal effect in limiting the CIA damage in all parameters. The beneficial effect of HYA and C4S was made evident by measuring the incidence of CIA. Similar results were obtained for the mean arthritis severity score. HYA and C4S were also able to reduce the hindpaw diameter and the body weight decrease observed in the CIA rats receiving vehicle alone.
Lipid peroxidation is considered a critical mechanism of the injury that occurs during RA [9, 65]. The evidence supporting these biochemical changes is based on the analysis of a large number of intermediate products [66]. An indicative method, extensively used, of evaluating lipid peroxidation is analysis of tissue malonaldehyde [66]. The large amount of malonaldehyde found in the CIA plus vehicle group at day 21 is consistent with the occurrence of damage mediated by free radicals. Treatment with the two GAGs produced a significant attenuation of cartilage injury.
The production of oxygen free radicals that occurs with the development of arthritis in the articular cartilage leads to decreased GSH and SOD levels as a consequence of their consumption during oxidative stress and cellular lysis [67, 68]. This decrease contributes to increased cellular damage by favouring attack by free radicals. HYA and C4S blunted the depletion of GSH and SOD, probably by competing in scavenging for free radicals, and as a result helped to preserve the integrity of cellular membranes in the injured cartilage.
The myeloperoxidase results demonstrated that a strong decrease in infiltration of polymorphonuclear cells occurred in the synovial tissue of joints. This decrease and the other biochemical parameters were evaluated by histological analysis, confirming the protective effects of the two polymers. We suggest that the decrease in neutrophil accumulation induced by GAGs might be due to the inhibition of lipid peroxidation and the consequent decrease in the chemotactic reduction of peroxide [66].
Several areas of investigation have indirectly implicated TNF-α as a contributor to cellular damage in CIA. The high levels of this cytokine can be interpreted as a progression of cartilage cell injury [69]. The antioxidant activity of GAGs might have lowered plasma TNF-α concentrations and consequently mitigated articular cell damage.
Which, then, is the mechanism by which HYA and C4S protect the cartilage against free radical attack? HYA and C4S are linear polymers formed by alternating hexuronic acid and hexosamine units. HYA is non-sulphated compound whereas C4S is sulphated in position 4 of the amino sugar. One plausible explanation for the antioxidant activity of HYA and C4S is the presence in their structure of a carboxylic group that might bind transition metals such as Cu2+ or Fe2+ [59, 70], which are in turn responsible for the initiation of Fenton's reaction. In this reaction the oxidation of Fe2+ or Cu2+ to Fe3+ or Cu3+ leads to the formation of the detrimental hydroxyl radical (OH) from hydrogen peroxide. In this way these molecules might function as metal chelators like the antioxidant deferoxamine or the calcium chelator EDTA. Another antioxidant mechanism might be the direct scavenging effect of HYA and C4S on free-radical molecules, especially the OH radical or other Fenton's reaction intermediates such as the superoxide anion [19, 61].
These hypotheses could explain the increased levels of GAGs during RA. In fact, elevated circulating levels of GAGs might be a biological response to the production of free radicals. The aim of our work was to increase the physiological levels of HYA and C4S by administering these compounds endogenously. Several studies have previously reported an increase in blood and tissue distribution of HYA and C4S after their administration in rats [71, 72]. We suggest that, after intraperitoneal administration, these polymers might be absorbed by the lymphatic system and blood vessels and then they may accumulate at the sites of production of free radicals. In addition, the amounts of HYA and C4S might cause an increase in total negative charge with a consequent inhibition of lymphocyte interactions with the target cell surface. In this way GAGs could exert a positive anti-inflammatory effect.
Treatment was performed intraperitoneally and not by the oral route, as previously reported by other investigators [64, 73]. We suggest that this different mode of administration, as demonstrated by our results, is preferable to the oral route because a large amount of the two polymers can reach the inflamed cartilage and they are available locally to neutralise transition metals or other reactive species.
Conclusions
The evidence of benefits obtained in this study show that GAGs are neither a specific drug nor an alternative to actual therapies for RA, but they represent a small step in our understanding of this complex pathology. The hypotheses about the mechanism of action in our model, reported above, need several further investigations for confirmation. GAGs might then be a useful tool in the study of the involvement of free radicals in CIA or in assessing other models of damage induced by free radicals.
Abbreviations
- CIA:
-
collagen-induced arthritis
- C4S:
-
chondroitin-4-sulphate
- GAG:
-
glycosaminoglycan
- GSH:
-
reduced glutathione
- HYA:
-
hyaluronic acid
- RA:
-
rheumatoid arthritis
- ROS:
-
reactive oxygen species
- SOD:
-
superoxide dismutase
- TNF-α:
-
tumour necrosis factor-α.
References
Feldmann M, Brennan FM, Maini RN: Rheumatoid arthritis. Cell. 1996, 85: 307-310. 10.1016/S0092-8674(00)81109-5.
Jawaheer D, Thomson W, MacGregor AJ, Carthy D, Davidson J, Dyer PA, Silman AJ, Ollier WE: 'Homozygosity' for the HLA-DR shared epitope contributes the highest risk for rheumatoid arthritis concordance in identical twins. Arthritis Rheum. 1994, 37: 681-6866.
Wooley PH, Luthra HS, Stuart JM, David CS: Type II collagen-induced arthritis in mice. I. Major histocompatibility complex (I region) linkage and antibody correlates. J Exp Med. 1981, 154: 688-700. 10.1084/jem.154.3.688.
Blake DR, Merry P, Unsworth J, Kidd BL, Outhwaite JM, Ballard R, Morris CJ, Gray L, Lunec J: Hypoxic-reperfusion injury in the inflamed human joint. Lancet. 1989, i: 289-293. 10.1016/S0140-6736(89)91305-6.
Lunec J: Free radicals: their involvement in disease processes. Ann Clin Biochem. 1990, 27: 173-182.
Dormandy TL: Free-radical pathology and medicine. A review. J R Coll Physicians Lond. 1989, 23: 221-227.
Halliwell B, Gutteridge JM: The antioxidants of human extracellular fluids. Arch Biochem Biophys. 1990, 280: 1-8. 10.1016/0003-9861(90)90510-6.
Heliovaara M, Knekt P, Aho K, Aaran RK, Alfthan G, Aromaa A: Serum antioxidants and risk of rheumatoid arthritis. Ann Rheum Dis. 1994, 53: 51-53.
Gambhir JK, Lali P, Jain AK: Correlation between blood antioxidant levels and lipid peroxidation in rheumatoid arthritis. Clin Biochem. 1997, 30: 351-355. 10.1016/S0009-9120(96)00007-0.
Iozzo RV: Matrix proteoglycans: from molecular design to cellular function. Annu Rev Biochem. 1998, 67: 609-652. 10.1146/annurev.biochem.67.1.609.
Friman C, Eronen I, Videman T: Plasma glycosaminoglycans in experimental osteoarthritis caused by immobilization. J Rheumatol. 1982, 9: 292-294.
Reddy GK, Dhar SC: Metabolism of glycosaminoglycans in tissue of arthritic rat. Mol Cell Biochem. 1991, 106: 117-124.
Caterson B, Flannery CR, Hughes CE, Little CB: Mechanisms involved in cartilage proteoglycan catabolism. Matrix Biol. 2000, 19: 333-344. 10.1016/S0945-053X(00)00078-0.
Campo GM, Squadrito F, Altavilla D, Squadrito Giov, Avenoso A, Canale P, Ioculano M, Sperandeo A, Caputi AP: Protection of ischemic and reperfused rat myocardium by the nonglucocorticoid 21-aminosteroid U-74389G, a new inhibitor of lipid peroxidation. J Pharmacol Exp Ther. 1996, 277: 333-340.
Kroger H, Miesel R, Dietrich A, Ohde M, Altrichter S, Braun C, Ockenfels H: Suppression of type II collagen-induced arthritis by N-acetyl-L-cysteine in mice. Gen Pharmacol. 1997, 29: 671-674. 10.1016/S0306-3623(96)00570-8.
Campo GM, Squadrito F, Campo S, Altavilla D, Quartarone C, Ceccarelli S, Ferlito M, Avenoso A, Squadrito G, Saitta A, Caputi AP: Beneficial effect of Raxofelast, an hydrophilic vitamin E analogue, in the rat heart after ischemia and reperfusion injury. J Mol Cell Cardiol. 1998, 30: 1493-1503. 10.1006/jmcc.1998.0713.
Campo GM, Squadrito F, Ceccarelli S, Calò M, Avenoso A, Campo S, Squadrito G, Altavilla D: Reduction of Carbon tetrachloride-induced rat liver injury by IRFI 042, a novel dual vitamin E-like antioxidant. Free Radic Res. 2001, 34: 379-393.
Sato H, Takahashi T, Ide H, Fukushima T, Tabata M, Sekine F, Kobayashi K, Negishi M, Niwa Y: Antioxidant activity of synovial fluid, hyaluronic acid, and two subcomponents of hyaluronic acid. Synovial fluid scavenging effect is enhanced in rheumatoid arthritis patients. Arthritis Rheum. 1988, 31: 63-71.
Presti D, Scott JE: Hyaluronan-mediated protective effect against cell damage caused by enzymatically produced hydroxyl (OH) radicals is dependent on hyaluronan molecular mass. Cell Biochem Funct. 1994, 12: 281-288.
Cortivo R, Brun P, Cardarelli L, O'Regan M, Radice M, Abatan-gelo G: Antioxidant effects of hyaluronan and its alpha-methyl-prednisolone derivative in chondrocyte and cartilage cultures. Semin Arthritis Rheum. 1996, 26: 492-501. 10.1016/S0049-0172(96)80030-8.
Albertini R, De Luca G, Passi A, Moratti R, Abuja PM: Chondroitin-4-sulfate protects high-density lipoprotein against copper-dependent oxidation. Arch Biochem Biophys. 1999, 365: 143-149. 10.1006/abbi.1999.1150.
Sela S, Shurtz-Swirski R, Shapiro G, Nasser L, Hamzi M, Shasha SM, Kristal B: Oxidative stress during hemodialysis: effect of heparin. Kidney Int. 2001, 78: S159-S163. 10.1046/j.1523-1755.2001.07830.x.
Cremer M: Type II collagen-induced arthritis in rats. In Handbook of Animal Models for the Rheumatic Diseases. Edited by: Greenwald RA, Diamond HS. 1988, Boca Raton, FL: CRC Press, 1: 17-27.
Campo GM, Campo S, Ferlazzo AM, Vinci R, Calatroni A: Improved high-performance liquid chromatographic method to estimate aminosugars and its application to glycosaminoglycan determination in plasma and serum. J Chromatogr. 2001, 765: 151-160. 10.1016/S0378-4347(01)00427-3.
Larsson P, Kleinau S, Holmdahl R, Klareskog L: Homologous type II collagen-induced arthritis in rats. Characterization of the disease and demonstration of clinically distinct forms of arthritis in two strains of rats after immunization with the same collagen preparation. Arthritis Rheum. 1990, 33: 693-701.
Durie FH, Fava RA, Foy TM, Aruffo A, Ledbetter JA, Noelle RJ: Prevention of collagen-induced arthritis with an antibody to gp39, the ligand for CD40. Science. 1993, 261: 1328-1330.
Ellman GL: Tissue sulphydryl groups. Arch Biochem Biophys. 1959, 82: 70-77. 10.1016/0003-9861(59)90090-6.
Lefkowitz DL, Gelderman MP, Fuhrmann SR, Grahams Starnes JD, Lefkowitz SS, Bollen A, Moguilevsky N: Neutrophil myeloperoxidase-macrophage interactions perpetuate chronic inflammation associated with experimental arthritis. Clin Immun. 1999, 91: 145-155. 10.1006/clim.1999.4696.
Mullane KM, Kraemer R, Smith B: Myeloperoxidase activity as a quantitative assessment of neutrophil infiltration into ischaemic myocardium. J Pharmacol Methods. 1985, 14: 157-167. 10.1016/0160-5402(85)90029-4.
Bauerova K, Bezek A: Role of reactive oxygen and nitrogen species in etiopathogenesis of rheumatoid arthritis. Gen Physiol Biophys. 1999, 18: 15-20.
VanderBorght A, Geusens P, Raus J, Stinissen P: The autoimmune pathogenesis of rheumatoid arthritis: role of autoreactive T cells and new immunotherapies. Semin Arthritis Rheum. 2001, 31: 160-175. 10.1053/sarh.2001.27736.
Knight JA: Free radicals, antioxidants, and the immune system. Ann Clin Lab Sci. 2000, 30: 145-158.
Babior BM: Phagocytes and oxidative stress. Am J Med. 2000, 109: 33-44. 10.1016/S0002-9343(00)00481-2.
Haqqi TM, Anthony DD, Gupta S, Ahmad N, Lee MS, Kumars GK, Mukhtar H: Prevention of collagen-induced arthritis in mice by a polyphenolic fraction from green tea. Proc Natl Acad Sci USA. 1999, 96: 4524-4529. 10.1073/pnas.96.8.4524.
Corvo ML, Boerman OC, Oyen WJG, Van Bloois LV, Cruz MEM, Crommelin DJA, Storm G: Intravenous administration of superoxide dismutase entrapped in long circulating liposomes. II. In vivo fate in a rat model of adjuvant arthritis. Biochim Biophys Acta. 1999, 1419: 325-334. 10.1016/S0005-2736(99)00081-4.
Sakai A, Hirano T, Okazaki R, Okimoto N, Tanaka K, Nakamura T: Large-dose ascorbic acid administration suppresses the development of arthritis in adjuvant-infected rats. Arch Orthop Trauma Surg. 1999, 119: 121-126. 10.1007/s004020050374.
Darlington LG, Stone TW: Antioxidants and fatty acids in the amelioration of rheumatoid arthritis and related disorders. Br J Nutr. 2001, 85: 251-269.
Ostrakhovitch EA, Afanas'ev IB: Oxidative stress in rheumatoid arthritis leukocytes: suppression by rutin and other antioxidants and chelators. Biochem Pharmacol. 2001, 62: 743-746. 10.1016/S0006-2952(01)00707-9.
Calatroni A, Donnelly PV, Di Ferrante N: The glycosaminoglycans of human plasma. J Clin Invest. 1969, 48: 332-343.
Staprans I, Felts JM: Isolation and characterization of glycosaminoglycans in human plasma. J Clin Invest. 1985, 76: 1984-1991.
Ferlazzo AM, Campo S, Vinci R, Ferrlazzo A, Calatroni A: Concentration and composition of serum and plasma glycosaminoglycans in domestic animal species. Comp Biochem Physiol. 1997, 118B: 935-942.
Calatroni A, Vinci R, Ferlazzo AM: Characteristics of the interactions between acid glycosaminoglycans and proteins in normal human plasma as revealed by the behaviour of the protein-polysaccharide complexes in ultrafiltration and chromatographic procedures. Clin Chim Acta. 1992, 206: 167-180. 10.1016/0009-8981(92)90086-6.
Friman C, Nordstrom D, Eronen I: Plasma glycosaminoglycans in systemic lupus erythematosus. J Rheumatol. 1987, 14: 1132-1134.
Laurent TC, Laurent UBG, Fraser JRE: Serum hyaluronan as a disease marker. Ann Med. 1996, 28: 241-253.
Radhakrishnamurthy B, Tracy RE, Dalferes ER, Berenson GS: Proteoglycans in human coronary arteriosclerotic lesions. Exp Mol Pathol. 1998, 65: 1-8. 10.1006/exmp.1998.2222.
Gambaro G, van der Woude FJ: Glycosaminoglycans: use in treatment of diabetic nephropathy. J Am Soc Nephrol. 2000, 11: 359-368. 10.1159/000017267.
Engstrom-Laurent A, Hallgren R: Circulating hyaluronate in rheumatoid arthritis: relationship to inflammatory activity and the effect of corticosteroid therapy. Ann Rheum Dis. 1985, 44: 83-88.
Roughley PJ: Articular cartilage and changes in arthritis: non-collagenous proteins and proteoglycans in the extracellular matrix of cartilage. Arthritis Res. 2001, 3: 342-347. 10.1186/ar326.
Bjork J, Kleinau S, Tengblad A, Smedegard G: Elevated levels of serum hyaluronate and correlation with disease activity in experimental models of arthritis. Arthritis Rheum. 1989, 32: 306-311.
Van der Kraan PM, de Lange J, Vitters EL, van Beuningen HM, van Osch GJ, van Lent PL, van den Berg WB: Analysis of changes in proteoglycan content in murine articular cartilage using image analysis. Osteoarthritis Cartilage. 1994, 2: 207-214.
Thonar EJ, Lenz ME, Klintworth GK, Caterson B, Pachman LM, Glickman P, Katz R, Huff J, Kuettner KE: Quantification of keratan sulfate in blood as a marker of cartilage catabolism. Arthritis Rheum. 1985, 28: 1367-1376.
Ruoslahti E, Yamaguchi Y: Proteoglycans as modulators of growth factor activities. Cell. 1991, 64: 867-869. 10.1016/0092-8674(91)90308-L.
Vestweber D, Blanks J: Mechanisms that regulate the function of the selectins and their ligands. Physiol Rev. 1999, 79: 181-213.
Haslinger B, Mandl-Weber S, Sellmayer A, Sitter T: Hyaluronan fragments induce the synthesis of MCP-1 and IL-8 in cultured human peritoneal mesothelial cells. Cell Tissue Res. 2001, 305: 79-86. 10.1007/s004410100409.
Sconocchia G, Campagnano L, Adorno D, Iacona A, Cococcetta NY, Boffo V, Amadori S, Casciani CU: CD44 ligation on peripheral blood polymorphonuclear cells induces interleukin-6 production. Blood. 2001, 97: 3621-3627. 10.1182/blood.V97.11.3621.
Syrokou A, Tzanakakis G, Tsegenidis T, Hjerpe A, Karamanos NK: Effects of glycosaminoglycans on proliferation of epithelial and fibroblast human malignant mesothelioma cells: a structure-function relationship. Cell Prolif. 1999, 32: 85-99. 10.1046/j.1365-2184.1999.32230085.x.
Foschi D, Castoldi L, Radaelli E, Abelli P, Calderini G, Rastrelli A, Mariscotti C, Marazzi M, Trabucchi E: Hyaluronic acid prevents oxygen free-radical damage to granulation tissue: a study in rats. Int J Tissue React. 1990, 12: 333-339.
Arai H, Kashiwagi S, Nagasaka Y, Uchida K, Hoshii Y, Nakamura K: Oxidative modification of apolipoprotein E in human very-low-density lipoprotein and its inhibition by glycosaminoglycans. Arch Biochem Biophys. 1999, 367: 1-8. 10.1006/abbi.1999.1222.
Albertini R, Passi A, Abuja PM, De Luca G: The effect of glycosaminoglycans and proteoglycans on lipid peroxidation. Int J Mol Med. 2000, 6: 129-136.
Graf J, Neusel E, Schneider E, Niethard FU: Intra-articular treatment with hyaluronic acid in osteoarthritis of the knee joint: a controlled clinical trial versus mucopolysaccharide polysulfuric acid ester. Clin Exp Rheumatol. 1993, 11: 367-372.
Breborowicz A, Wieczorowska K, Martis L, Oreopoulos DG: Glycosaminoglycan chondroitin sulphate prevents loss of ultrafiltration during peritoneal dialysis in rats. Nephron. 1994, 67: 346-350.
Shankland WE: The effects of glucosamine and chondroitin sulfate on osteoarthritis of the TMJ: a preliminary report of 50 patients. Cranio. 1998, 16: 230-235.
Leffler CT, Philippi AF, Leffler SG, Mosure JC, Kim PD: Glucosamine, chondroitin, and manganese ascorbate for degenerative joint disease of the knee or low back: a randomized, double-blind, placebo-controlled pilot study. Mil Med. 1999, 164: 85-91.
Beren J, Hill SL, Diener-West M, Rose NR: Effect of preloading oral glucosamine HCl/chondroitin sulfate/manganese ascorbate combination on experimental arthritis in rats. Exp Biol Med. 2001, 226: 144-151.
Jira W, Spiteller G, Richter A: Increased levels of lipid oxidation products in low density lipoproteins of patients suffering from rheumatoid arthritis. Chem Phys Lipids. 1997, 87: 81-89. 10.1016/S0009-3084(97)00030-3.
Wills ED: Evaluation of lipid peroxidation in lipid and biological membranes. In Biochemical Toxicology; a Practical Approach. Edited by: Snell K, Mullock B. 1987, Oxford: IRL Press, 127-152.
Kizilntuc A, Cogalgil S, Cerrahoglu L: Carnitine and antioxidants levels in patients with rheumatoid arthritis. Scand J Rheumatol. 1998, 27: 441-445. 10.1080/030097498442271.
Hassan MQ, Hadi RA, Al-Rawi ZS, Padron VA, Stohs SJ: The glutathione defense system in the pathogenesis of rheumatoid arthritis. J Appl Toxicol. 2001, 21: 69-73. 10.1002/jat.736.
Feldmann M, Brennan FM, Foxwell BM, Maini RN: The role of TNF-alpha and IL-1 in rheumatoid arthritis. Curr Dir Autoimmun. 2001, 3: 188-199.
Volpi N, Tarugi P: Influence of chondroitin sulfate charge density, sulfate group position, and molecular mass on Cu++-mediated oxidation of human low-density lipoproteins: effect of normal human plasma-derived chondroitin sulfate. J Biochem (Tokyo). 1999, 125: 297-304.
Palmieri L, Conte A, Giovannini L, Lualdi P, Ronca G: Metabolic fate of exogenous chondroitin sulfate in the experimental animal. Arzneimittelforschung. 1990, 40: 319-323.
Gustafson S, Bjorkman T: Circulating hyaluronan, chondroitin sulphate and dextran sulphate bind to a liver receptor that does not recognize heparin. Glycoconj J. 1997, 14: 561-568. 10.1023/A:1018528308132.
Omata T, Itokazu Y, Inoue N, Segawa Y: Effects of chondroitin sulfate-C on articular cartilage destruction in murine collagen-induced arthritis. Arzneimittelforschung. 2000, 50: 148-153.
Acknowledgements
We gratefully acknowledge the expert technical assistance of Enea Letterio. This study was supported by a grant from PRA (Research Athenaeum Project) of MURST, Italy.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
None declared.
Rights and permissions
About this article
Cite this article
Campo, G.M., Avenoso, A., Campo, S. et al. Efficacy of treatment with glycosaminoglycans on experimental collagen-induced arthritis in rats. Arthritis Res Ther 5, R122 (2003). https://doi.org/10.1186/ar748
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/ar748