
Abstract
We have recently found that matrix metalloproteinases (MMPs) are targets for T-cell and B-cell reactivity in experimental arthritis. In the present article, we investigate whether modulation of MMP-specific T-cell responses could influence the course of adjuvant arthritis (AA). Lewis rats were treated nasally with MMP peptides prior to or after AA induction. Administration of the MMP-10 or the MMP-16 peptide prior to AA induction reduced the arthritic symptoms. In contrast, administration of the MMP-10 peptide after AA induction aggravated the arthritic symptoms. The present study shows the possible usefulness of MMP peptides for immunotherapy. However, a clear understanding of proper timing of peptide administration is crucial for the development of such therapies.
Similar content being viewed by others


Introduction
Rheumatoid arthritis (RA) is an autoimmune disease, in which autoreactive T cells are considered to play a central role in the autodestructive process [1, 2]. For the development of long-term acting, antigen-specific immunotherapies, considerable attention has been devoted to intervening in the autoimmune process via modulation of the autoaggressive T-cell response. Several studies in experimental autoimmune models have shown that it is possible to induce antigen-specific tolerance, leading to disease resistance [3–9]. This tolerance induction is dependent on several factors such as adjuvant use, antigen dose and the route of antigen administration (as reviewed in [10]). Nasal antigen administration appears to be an especially efficient way to induce mucosal tolerance in experimental arthritis models [9, 11–13].
We recently published a computer search profile to predict T-cell self-epitopes recognized in adjuvant arthritis (AA) [14]. This search profile led to the identification of matrix metalloproteinases (MMPs) as targets for T-cell recognition in experimental arthritis. Interestingly, active arthritis could be induced by immunization with synthetic peptides containing the predicted MMP-3, MMP-10 or MMP-16 T-cell epitope. Moreover, CD4+ T cells isolated from rats immunized with MMP-3 peptide, MMP-10 peptide or MMP-16 peptide could transfer arthritis into naïve Lewis rats.
As well as being a target for T cells, we have demonstrated that MMPs also become a target for B cells during the course of AA. This is shown by the appearance of MMP-3-specific IgM and IgG titers after AA induction (JHM van Bilsen et al., unpublished observations).
In the present study, we investigate the usefulness of the recently identified MMP epitopes to modulate MMP-specific T-cell responses, thereby interfering in the course of AA in Lewis rats.
Materials and methods
See Supplementary material for full details of the peptides, the animals and the arthritis model.
Nasal peptide treatment
Rats were treated nasally with 10 μl of 10 μg/μl peptide in PBS using a micropipette. In the pretreatment protocol, rats were treated four times on day –14, day –11, day –7 and day –4, preceding AA induction. In the treatment protocol, treatment was started after AA induction when more than 50% of the animals showed weight loss (i.e. at the onset of clinical arthritis). Animals were equally divided over different treatment groups based on their weight loss and the AA score. Nasal peptide administration was repeated four times with an interval of 3–4 days.
Delayed-type hypersensitivity assay
Peptides were dissolved in PBS (2 mg/ml) and 50 μl was injected in one ear. PBS was injected in the contra-lateral ear, and served as the control. The delayed-type hypersensitivity (DTH) reaction was determined by measuring the ear thickness 48 hours after injection with a pressure sensitive micrometer. Data are expressed as the mean difference in ear swelling between the peptide-treated and the PBS-treated ears (mm/100) ± standard error of the mean.
MHC class II–peptide binding assay
See Supplementary material for full details of the MHC class II–peptide binding assay.
Ex vivo proliferation assays
See Supplementary material for full details of the ex vivo proliferation assays.
Results
Natural occurrence of MMP peptide-specific T cells during AA
We have recently identified T-cell epitopes in MMP-3, MMP-10 and MMP-16 recognized during AA (JHM van Bilsen et al., unpublished observations). We monitored T-cell responses to these MMP epitopes during the course of AA in the popliteal lymph nodes (PLNs) to represent the local immune response in the arthritic joints. Although only very low proliferative responses were detected, they tend to rise during the course of the disease (see Supplementary material).
To gain more insight into these MMP-specific T-cell responses, we also performed DTH assays. As presented in Table 1, MMP-3444–458, MMP-10329–343 and MMP-16539–553 induced a significant DTH reaction at day 20 and day 27 after arthritis induction. Naïve rats did not show DTH responses to any of the tested peptides, and no DTH reaction was observed with the control ovalbumin peptide OVA323–339.
We next evaluated the MMP peptides for binding to the Lewis rat MHC class II molecule, RT1.BL, which was used as a prerequisite for the selection of the peptides [14]. Although all three peptides (MMP-3444–458, MMP-10329–343 and MMP-16539–553) bound to RT1.BL, as was expected from the search profile, no correlation was observed between their actual MHC binding affinity and the observed T-cell responses (see Supplementary material).
Altogether, these results indicate that the selected MMP epitopes are naturally processed and presented during arthritis, and that they are able to activate MMP-specific T cells.
Nasal peptide administration prior to AA induction
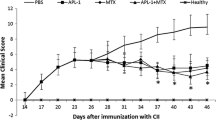
We then investigated whether we could modulate MMP-specific T-cell responses, thereby interfering in the development of AA. To this end, rats were treated nasally with MMP peptides or OVA323–339 (negative control) before AA induction. As shown in Figure 1, nasal administration of MMP-3444–458 had no influence on AA development as compared with treatment with OVA323–339. In contrast, nasal administration of MMP-10329–343 and MMP-16539–553 significantly reduced the clinical signs of AA. The clinical findings were consistent with the observed changes in bodyweight (see Supplementary material).

Modulation of adjuvant arthritis development after nasal administration of matrix metalloproteinase (MMP) peptides prior to adjuvant arthritis. The nasal treatment was repeated four times (arrows). Data shown are mean arthritis scores ± standard error of the mean of two experiments (n = 10 rats per group). *P < 0.05 (two-tailed Mann–Whitney test) compared with the ovalbumin peptide(OVA323–339)-treated group.
Nasal peptide administration after AA induction
Furthermore, we investigated whether it was possible to intervene in ongoing AA. We treated rats nasally with MMP peptides and OVA323–339 at the onset of clinical arthritis. As shown in Figure 2, treatment with MMP-3444–458 or MMP-16539–553 did not influence the course of the disease compared with OVA323–339-treated rats. In contrast, MMP-10329–343-treated rats showed an aggravation of AA. These findings were consistent with the higher weight loss in the MMP-10 peptide-treated group (see Supplementary material).

Modulation of adjuvant arthritis (AA) development after nasal treatment with matrix metalloproteinase (MMP) peptides after AA induction. The nasal treatment was started at day +11, after immunization with Mycobacterium tuberculosis, when more than 50% of the animals showed weight loss (i.e. at the onset of clinical arthritis). The nasal treatment was repeated four times (arrows). Data shown are mean arthritis scores ± standard error of the mean of five rats (MMP-10 and MMP-16 peptide groups) or 10 rats (MMP-3 peptide group) per group. OVA323–339, ovalbumin peptide.
Lymphocyte proliferation of MMP peptide-treated rats
To analyze whether the interference in AA after nasal administration of MMP-10329-343 or MMP-16539–553 was accompanied by tolerance induction, PLN cells of peptide-treated rats were tested for proliferative responses to the dominant mycobacterial 65 kDa heat-shock protein epitope 178–186 (hsp65178–186), recognized by arthritogenic T cells in AA [9].
Although very low proliferative responses were detected, MMP-10 peptide treatment before AA induction resulted in a decreased proliferation to mycobacterial hsp65178–186 as compared with the untreated group. In contrast, PLN cells from rats that received MMP-10 peptide treatment after AA induction showed an increased proliferative response to mycobacterial hsp65178–186 (Fig. 3).

Popliteal lymph node cell proliferative responses after nasal matrix metalloproteinase (MMP) peptide administration prior to (left) or after (right) adjuvant arthritis (AA) induction. Responses were measured 35 days after AA induction. The irrelevant peptides used were MBP72–85 or ovalbumin peptide, OVA323–339. Data are expressed as the mean stimulation index (SI) of three or four organs ± standard error of the mean. *P < 0.05.
Both these findings are consistent with the respectively lower and higher AA scores observed after MMP-10 peptide pretreatment or treatment after AA induction. In accordance with the marginal disease modulation after MMP-16539–553 therapy, no alteration of the proliferative mycobacterial hsp65178–186 response was observed.
Discussion
In the present study, we explored the possibility of using recently identified MMP T-cell epitopes for immunotherapy in AA. First, we monitored T-cell responses to the MMP epitopes during the course of AA. In general, low proliferative responses to these epitopes were detected, which were accompanied by specific DTH reactions. We have previously shown that the proliferative response to mycobacterial 65 kDa heat-shock protein (hsp65) 178–186, which is the dominant epitope recognized by arthritogenic T cells in AA [9], is also very low when tested in a polyclonal lymph node cell population [9] (Fig. 3).
To analyze whether the low MMP-specific proliferative responses are due to, for example, low precursor frequency or tolerance, it would be necessary to isolate and further characterize the MMP-specific T cells. We are currently developing a specific T-cell capture assay based on liposomal-bound MHC–peptide complexes to isolate such cells [15]. Interestingly, although the MMP epitopes greatly differed in MHC class II RT1.BL binding affinity, no differences in DTH reaction and/or proliferation were observed, indicating that these epitopes become a target for T-cell recognition irrespective of their MHC binding affinity. It was previously suggested that immunotherapy is most successful with high-affinity MHC binders [16]. However, the present study shows that the strongest immunomodulatory peptide, MMP-10329–343, was a weak MHC class II RT1.BL binder.
The upregulation of MMP-3 and its pathogenic role in arthritis has been shown in numerous reports, while only a few reports describe the presence of MMP-10 and MMP-16 in the synovium of RA patients [17, 18]. Although MMP-10 and MMP-16 have been suggested to be involved in connective tissue/bone remodeling around prostheses [19, 20], their role in arthritis is less clear. Surprisingly, peptides derived from MMP-10 and MMP-16, but not from MMP-3, can alter the course of AA after nasal administration. The observed opposite effect of nasal therapy using MMP-10 peptide furthermore illustrates that we seem to target the proper cell population to interfere in arthritis, but that the desired disease inhibitory effect strongly depends on the timing of T-cell modulation.
Other studies have also shown that mucosal therapy might in some cases induce or even exacerbate T helper 1 cell autoimmunity [21–23]. Moreover, the critical aspect of proper timing has also been described for cytokine therapies as shown by both disease inhibition and exacerbation after in vivo administration of IFN-γ, IL-2 or IL-12 in experimental arthritis models [24–26].
In the present study, we stated that the improvement of arthritis symptoms after MMP-10 peptide pretreatment coincided with a decreased proliferative response to the critical T-cell epitope for the induction of AA, mycobacterial hsp65178–186. Although the proliferative response to this epitope is difficult to measure in a polyclonal lymph node cell population, our results suggest that MMP-10 peptide pretreatment inhibits the response to the arthritogenic epitope via bystander suppression. Immunotherapy using agents that induce T-cell-mediated bystander suppression makes it unnecessary to identify the self-antigens involved in the initiation of the arthritis process, but makes it possible to exploit spreading epitopes or other self-antigens that become available during the arthritis process for disease intervention.
Conclusion
The present study shows the possible usefulness of MMP peptide immunotherapy in arthritis, based on modulation of the T-cell response. However, a clear understanding of proper timing of MMP peptide-based immunotherapy and the consequences for T-cell modulation will be crucial for the development of such therapies.
Supplementary material
Supplementary materials and methods
Peptides
Guinea pig myelin basic protein (MBP)72–85 (QKSQRSQDENPV), Mycobacterium tuberculosis hsp65178–186 (SNTFGLQLE) and chicken OVA323–339 (ISQAVHAAHAEINEAGR) peptides were obtained from Isogen Bioscience (Maarn, The Netherlands). Rat MMP-3444–458(FLYFFSGSSQLEFDP), rat MMP-10329–343 (SAFWPSLPSGLDAAY) and human MMP-16539–553 (VKEGHSPPDDVDIVI)were synthesized via automated simultaneous multiple peptide synthesis [S1]. Peptides were obtained as C-terminal amides after cleavage with 90–95% trifluoroacetic acid/scavenger cocktails. Peptides were analyzed by reversed-phase high-performance liquid chromatography, and checked via electrospray ionization mass spectrometry on an ion-trap mass spectrometer (LCQ; Thermoquest, Breda, The Netherlands). Peptide MBP72–85 was biotinylated during synthesis and used as the marker peptide in the RT1.BL MHC–peptide binding assays.
Animals
Male inbred Lewis rats were obtained from Maastricht University (Maastricht, The Netherlands). Rats were 6–8 weeks old at the start of each experiment. This study was approved by the Dutch Committee of Animal Experiments.
Induction and clinical evaluation of AA
AA was induced by intradermal injection at the base of the tail with 0.1 ml of 5 mg/ml heat-killed M. tuberculosis H37RA in incomplete Freund's adjuvant (DIFCO Laboratories, Detroit, MI, USA). Rats were examined for clinical signs of arthritis in a blinded set-up. The severity of arthritis was scored by grading each paw from 0 to 4 based on erythema, swelling and deformity of the joints, resulting in a maximum score of 16.
MHC class II–peptide binding assay
Lewis rat MHC class II molecules, RT1.BL, were purified from the Z1a T cell clone through affinity chromatography using the OX6 monoclonal antibody, as described previously [S2]. Affinity-purified RT1.BL MHC molecules were solubilized in 1% n-β-octyl glucopyranoside (Sigma-Aldrich Chemie BV, Zwijndrecht, The Netherlands). The MHC class II–peptide binding studies were performed using the competitive binding assay as described previously [S2]. Briefly, 3 M RT1.BL was incubated with 400 nM biotinylated marker peptide and a concentration range (0–256 M) of unlabeled competitor peptide at room temperature for 40 hours at pH5 in the presence of a protease inhibitor mix.
The MHC–peptide mixtures were analyzed via nonreducing SDS-PAGE, followed by western blotting (Hybond-ECL; Amersham, Pharmacia Biotech Benelux, Roosendaal, The Netherlands). Biotinylated peptides were visualized on hyperfilm (Amersham, Pharmacia Biotech Benelux, Roosendaal, The Netherlands), through enhanced chemiluminescence (Western Blot ECL kit; Amersham, Pharma-cia Biotech Benelux). Spots on the films were quantified by Molecular Analyst Software version 1.5 (Bio-Rad Laboratories B.V., Veenendaal, The Netherlands). The relative MHC binding affinity (IC50 value) is expressed as the concentration range of competitor peptide (M) resulting in 50% inhibition of the MHC binding of the marker peptide.
Ex vivo proliferation assays
At different time points after AA induction, PLNs were isolated and single cell suspensions were cultured in 96-well flat-bottomed microtiter plates (2 ×105 cells/well in triplicate) in the absence or presence of peptide (1 μg/ml or 10 μg/ml). After 3 days of culturing, proliferation was determined by overnight incorporation of [3H]thymidine (0.4 μCi/well; specific activity, 1 Ci/mmol; Amersham Pharmacia Biotech Benelux). Proliferation was expressed as the stimulation index (counts per minute of cells cultured with peptide divided by the counts per minute of cells cultured in medium only).
Statistics
Statistical evaluation was performed using GraphPad Prism version 3.0 (GraphPad Software, San Diego, CA, USA). To evaluate the effect of each peptide in the DTH reaction, the paired, two-tailed Student's t test was performed on the mean difference in ear swelling between the peptide-treated ear and the PBS-treated ear. The two-tailed Mann–Whitney U test was performed to compare the mean differences in DTH reactions between naïve and arthritic rats. For the comparison of arthritis scores and proliferative responses between the different groups, a two-tailed Mann–Whitney U test was used. P < 0.05 was considered statistically significant.
Supplementary results
Natural occurrence of MMP peptide-specific T cells during AA
The PLN responses were analyzed in lymphocyte proliferation assays at day 0 (naïve animals), day 10 (before clinical onset of disease), day 14 (onset of disease), day 21 (overt disease) and day 35 (no or limited clinical disease) after disease induction with M. tuberculosis/incomplete Freund's adjuvant. Supplementary Figure 1 shows the proliferative responses to the MMP peptides in a representative experiment. As can be seen, only very low proliferative responses were detectable, which tend to rise during the course of the disease.

Monitoring of proliferative responses to matrix metalloproteinase (MMP) peptides during adjuvant arthritis in popliteal lymph node cells (PLNC) of Lewis rats. All organs were tested separately. Data are expressed as the mean of the stimulation index (SI) of three to four rats ± standard error of the mean. At day 0, PLNC and inguinal lymph node cells of n = 4 rats were pooled and subsequently tested.
MHC binding affinity of MMP peptides
We previously selected the MMP peptides based on their putative binding to Lewis rat MHC class II RT1.BL[14]. In the present study, we determined the binding affinity of the MMP peptides for RT1.BL in a competitive peptide–MHC binding assay.
The relative binding affinities of the MMP epitopes are depicted in Supplementary Figure 2. The MMP epitopes could be ranked into three broadly defined categories:MMP-3244–258 is a good RT1.BL binder (IC50 ≤ 8 μM), MMP-16539–553 is an intermediate RT1.BL binder (IC50 = 64–128 μM), and MMP-10329–343 is a poor RT1.BL binder (IC50 = 128–256 μM).

Binding affinity of matrix metalloproteinase (MMP) peptides to purified RT1.BL. The binding affinity of the competitor peptides (MMP peptides) was tested in a competitive MHC class II peptide binding assay in a dose range of 0–256 μM. * IC50, concentration of competitor peptide (μM) resulting in 50% inhibition of the binding of marker peptide to RT1.BL, as evaluated by Molecular Analyst Software (see Supplementary Materials and methods).
Nasal peptide administration prior to AA induction
Nasal administration of MMP-10329–343 and MMP-16539–553significantly reduced the severity of AA. Weight loss (a sensitive objective measure of physical well-being) was analyzed as an addition parameter (Supplementary Figure 3). The mean body weight demonstrated that MMP-3444–458-treated rats showed the same weight loss as the OVA323–339-treated group, which is consistent with the comparable AA severity. In contrast, MMP-10329–343-treated rats showed a clear reduction in weight loss as compared with the OVA323–339-treated group, consistent with the lower AA scores. MMP-16539-553-treated rats showed no difference in weight loss as compared with the OVA323–339 control group.

Mean body weight of rats treated nasally with matrix metalloproteinase (MMP) peptide prior to adjuvant arthritis (AA) induction. The mean body weight is depicted as the percentage of the body weight at the time of AA induction. Data shown are the percentage of the body weight of two experiments (n = 10 rats per group) ± standard error of the mean. OVA323–339, ovalbumin peptide.
Nasal peptide administration after AA induction
MMP-10 peptide treatment after AA induction aggravated the arthritic symptoms of the Lewis rats. These findings were consistent with the lower mean body weight in the MMP-10 peptide-treated group compared with the OVA323–339 control group (Supplementary Figure 4). MMP-3 peptide-treated rats and MMP-16 peptide-treated rats showed no differences in body weight and clinical scores as compared with the OVA323–339-treated group.

Mean body weight of rats treated nasally with matrix metalloproteinase (MMP) peptides after induction of adjuvant arthritis (AA). The mean body weight is depicted as the percentage of the body weight at the time of AA induction. Data shown are percentage body weight ± standard error of the mean of two experiments with five rats (MMP-10 and MMP-16 peptide groups) or 10 rats (MMP-3 peptide group) per group. OVA323–339, ovalbumin peptide.
Abbreviations
- AA:
-
= adjuvant arthritis
- DTH:
-
= delayed-type hypersensitivity
- IC50:
-
= relative MHC binding affinity
- IFN:
-
= interferon
- IL:
-
= interleukin
- hsp65:
-
= 65 kDa heat-shock protein
- MHC:
-
= major histocompatibility complex
- MMP:
-
= matrix metalloproteinase
- OVA323–339:
-
= ovalbumin peptide
- PBS:
-
= phosphate-buffered saline
- PLN:
-
= popliteal lymph node
- RA:
-
= rheumatoid arthritis
- RT1.BL:
-
= Lewis rat MHC class II molecule.
References
Breedveld FC: New insights in the pathogenesis of rheumatoid arthritis. J Rheumatol. 1998, 53 (suppl): S3-S7.
Panayi GS, Lanchbury S, Kingsley GH: The importance of the T cell in initiating and maintaining the chronic synovitis of rheumatoid arthritis. Arthritis Rheum. 1992, 35: 729-735.
Barchan D, Souroujon MC, Im SH, Antozzi C, Fuchs S: Antigen-specific modulation of experimental myasthenia gravis: nasal tolerization with recombinant fragments of the human acetylcholine receptor alpha-subunit. Proc Natl Acad Sci USA. 1999, 96: 8086-8091. 10.1073/pnas.96.14.8086.
Karachunski PI, Ostlie NS, Okita DK, Conti-Fine BM: Prevention of experimental myasthenia gravis by nasal administration of synthetic acetylcholine receptor T epitope sequences. J Clin Invest. 1997, 100: 3027-3035.
Tian J, Atkinson MA, Clare-Salzler M, Herschenfeld A, Forsthuber T, Lehmann PV, Kaufman DL: Nasal administration of glutamate decarboxylase (GAD65) peptides induces Th2 responses and prevents murine insulin-dependent diabetes. J Exp Med. 1996, 183: 1561-1567.
Dick AD, Cheng YF, Liversidge J, Forrester JV: Intranasal administration of retinal antigens suppresses retinal antigen-induced experimental autoimmune uveoretinitis. Immunology. 1994, 82: 625-631.
Metzler B, Wraith DC: Inhibition of experimental autoimmune encephalomyelitis by inhalation but not oral administration of the encephalitogenic peptide: influence of MHC binding affinity. Int Immunol. 1993, 5: 1159-1165.
Laliotou B, Liversidge J, Forrester JV, Dick AD: Interphotoreceptor retinoid binding protein is a potent tolerogen in Lewis rat: suppression of experimental autoimmune uveoretinitis is retinal antigen specific. Br J Ophthalmol. 1997, 81: 61-67.
Prakken BJ, van der Zee R, Anderton SM, van Kooten PJ, Kuis W, van Eden W: Peptide-induced nasal tolerance for a mycobac-terial heat shock protein 60 T cell epitope in rats suppresses both adjuvant arthritis and nonmicrobially induced experimental arthritis. Proc Natl Acad Sci USA. 1997, 94: 3284-3289. 10.1073/pnas.94.7.3284.
Wauben MH: Immunological mechanisms involved in experimental peptide immunotherapy of T-cell-mediated diseases. Crit Rev Immunol. 2000, 20: 451-469.
Myers LK, Seyer JM, Stuart JM, Kang AH: Suppression of murine collagen-induced arthritis by nasal administration of collagen. Immunology. 1997, 90: 161-164. 10.1046/j.1365-2567.1997.00191.x.
Joosten LA, Coenen-de Roo CJ, Helsen MM, Lubberts E, Boots AM, van den Berg WB, Miltenburg AM: Induction of tolerance with intranasal administration of human cartilage gp-39 in DBA/1 mice: amelioration of clinical, histologic, and radiologic signs of type II collagen-induced arthritis. Arthritis Rheum. 2000, 43: 645-655. 10.1002/1529-0131(200003)43:3<645::AID-ANR22>3.0.CO;2-O.
Staines NA, Harper N, Ward FJ, Malmström V, Holmdahl R, Bansal S: Mucosal tolerance and suppression of collagen-induced arthritis (CIA) induced by nasal inhalation of synthetic peptide 184–198 of bovine type II collagen (CII) expressing a dominant T cell epitope. Clin Exp Immunol. 1996, 103: 368-375.
van Bilsen JHM, Wagenaar-Hilbers JPA, Boot EPJ, van Eden W, Wauben MHM: Searching for the cartilage-associated mimicry epitope in adjuvant arthritis. Autoimmunity. 2002,
Prakken B, Wauben M, Genini D, Samodal R, Barnett J, Mendivil A, Leoni L, Albani S: Artificial antigen-presenting cells as tool to exploit the immune 'synapse'. Nat Med. 2000, 6: 406-410.
Anderton S, Burkhart C, Metzler B, Wraith D: Mechanisms of central and peripheral T-cell tolerance: lessons from experimental models of multiple sclerosis. Immunol Rev. 1999, 169: 123-137.
Konttinen YT, Ainola M, Valleala H, Ma J, Ida H, Mandelin J, Kinne RW, Santavirta S, Sorsa T, Lopez-Otin C, Takagi M: Analysis of 16 different matrix metalloproteinases (MMP-1 to MMP-20) in the synovial membrane: different profiles in trauma and rheumatoid arthritis. Ann Rheum Dis. 1999, 58: 691-697.
Pap T, Shigeyama Y, Kuchen S, Fernihough JK, Simmen B, Gay RE, Billingham M, Gay S: Differential expression pattern of membrane-type matrix metalloproteinases in rheumatoid arthritis. Arthritis Rheum. 2000, 43: 1226-1232. 10.1002/1529-0131(200006)43:6<1226::AID-ANR5>3.0.CO;2-4.
Takei I, Takagi M, Santavirta S, Ida H, Ishii M, Ogino T, Ainola M, Konttinen YT: Messenger ribonucleic acid expression of 16 matrix metalloproteinases in bone-implant interface tissues of loose artificial hip joints. J Biomed Mater Res. 2000, 52: 613-620. 10.1002/1097-4636(20001215)52:4<613::AID-JBM5>3.3.CO;2-#.
Ishiguro N, Ito T, Kurokouchi K, Iwahori Y, Nagaya I, Hasegawa Y, Iwata H: mRNA expression of matrix metalloproteinases and tissue inhibitors of metalloproteinase in interface tissue around implants in loosening total hip arthroplasty. J Biomed Mater Res. 1996, 32: 611-617. 10.1002/(SICI)1097-4636(199612)32:4<611::AID-JBM14>3.0.CO;2-G.
Blanas E, Carbone FR, Allison J, Miller JFAP, Heath WR: Induction of autoimmune diabetes by oral administration of autoantigen. Science. 1996, 274: 1707-1709. 10.1126/science.274.5293.1707.
Lens JW, van den Berg WB, van de Putte LB, van den Bersselaar L: Flare-up of antigen-induced arthritis in mice after challenge with oral antigen. Clin Exp Immunol. 1984, 58: 364-371.
Weinberg AD, Whitham R, Swain SL, Morrison WJ, Wyrick G, Hoy C, Vandenbark AA, Offner H: Transforming growth factor-b enhances the in vivo effector function and memory phenotype of antigen-specific T helper cells in experimental autoimmune encephalomyelitis. J Immunol. 1992, 148: 2109-2117.
Jacob CO, Holoshitz J, van der Meide P, Strober S, McDevitt HO: Heterogeneous effects of IFN-gamma in adjuvant arthritis. J Immunol. 1989, 142: 1500-1505.
Joosten LA, Lubberts E, Helsen MM, van den Berg WB: Dual role of IL-12 in early and late stages of murine collagen type II arthritis. J Immunol. 1997, 159: 4094-4102.
Thornton S, Boivin GP, Kim KN, Finkelman FD, Hirsch R: Heterogeneous effects of IL-2 on collagen-induced arthritis. J Immunol. 2000, 165: 1557-1563.
van der Zee R, Anderton SM, Buskens CAF, Alonso de Velasco E, van Eden W: Heat shock protein T-cell epitopes as immunogenic carriers in subunit vaccines. In Peptides 1994. Proceedings of the Twenty-Third European Peptide Symposium, September 4–10, Braga, Portugal. Edited by: Maia HLS. 1994, Leiden, The Netherlands: ESCOM, 841-842.
Joosten I, Wauben MHM, Holewijn MC, Reske K, Pedersen LO, Roosenboom CF, Hensen EJ, van Eden W, Buus S: Direct binding of autoimmune disease related T cell epitopes to purified Lewis rat MHC class II molecules. Int Immunol. 1994, 6: 751-759.
Acknowledgements
The authors thank MC Grosfeld for the peptide–MHC binding assays. The research by Dr MHM Wauben was made possible by a fellowship of the Royal Netherlands Academy of Arts and Sciences.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
van Bilsen, J.H., Wagenaar-Hilbers, J.P., van der Cammen, M.J. et al. Successful immunotherapy with matrix metalloproteinase-derived peptides in adjuvant arthritis depends on the timing of peptide administration. Arthritis Res Ther 4, R2 (2002). https://doi.org/10.1186/ar421
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/ar421