
Abstract
Background
Fish entrainment through turbine intakes is one of the major issues for operators of hydropower facilities because it causes injury and/or mortality and adversely affects population abundance. Entrainment reduction strategies have been developed based on the behavior of downstream migrating fishes, particularly diadromous species. However, knowledge of the behavior of migratory fishes has very limited application for reducing the entrainment of resident fishes, including several species that represent important recreational and aboriginal fishery resources in reservoirs. In this study, we used fine-scale acoustic telemetry and state-space modeling to investigate behavioral attributes associated with entrainment risk of resident adult bull trout (Salvelinus confluentus) in a large hydropower reservoir in British Columbia, Canada.
Results
We found that adult bull trout resided longer in the vicinity of the powerhouse and moved closer to the turbine intakes in the fall and particularly in the winter. Bull trout were more likely to engage in exploratory behavior (characteristic of foraging or reduced activity) during periods when their body temperature was lower or higher than 6°C. We also detected diel changes in behavioral attributes, with bull trout distance to intakes and probability of exploratory behavior slightly increasing at night.
Conclusions
We hypothesize that the exploratory behavior in the forebay is associated with foraging for kokanee (nonanadromous form of Oncorhynchus nerka), which have been shown to congregate near the dams of hydropower reservoirs in the winter. Our study findings should be applicable to bull trout populations residing in other reservoirs and indicate that entrainment mitigation (for example, use of deterrent devices) should be focused on the fall and winter. This work also provides a framework for combining acoustic telemetry and state-space models to understand and categorize movement behavior of fish in reservoirs and, more generally, in any environment with fluctuating water levels.
Similar content being viewed by others

Background
One of the major challenges for hydropower operators is determining how to reduce the number of fish that are displaced from reservoirs to downstream waters through turbine intakes—a process termed entrainment [1, 2]. Entrainment can cause immediate fish mortality in a variety of ways (for example, strike, cavitation, pressure changes [3]) or delayed mortality due to injury and increased susceptibility to predation [1]. In some jurisdictions, there are regulations and guidelines that help reduce entrainment and the impacts that it can have on the abundance of fish populations. For example, resource agencies in the United States have set survival standards for salmonids migrating downstream through hydroelectric facilities in the Columbia River [4]. In Canada, the Fisheries Act prohibits any activity that causes serious harm to fish that are part of, or support, a fishery [5], and national guidelines for managing fish entrainment through turbines and other types of water intake structures are currently in development [6].
Currently, most of the efforts to quantify and reduce entrainment in hydropower facilities have focused on downstream migrating fishes, particularly diadromous species [7–9]. Detailed studies on the behavior of migrants as they enter the forebay (the area directly upstream of the dam) and approach the hydropower facility have informed the design and refinement of guidance systems to direct fish away from turbine intakes and into bypass structures [7, 10]. However, because migrants use the water flow as a cue to move past dams [11], knowledge of the movement behavior of downstream migrating fishes has very limited application for understanding and mitigating entrainment of resident fishes (that is, those that do not actively emigrate from reservoirs).
Resident fish can be accidentally entrained in hydropower facilities when using habitats near turbine intakes [11]. Indeed, many technical reports available through hydropower companies and regulatory agencies indicate variable levels of entrainment of resident fish [12–14]. Resident juvenile fish are particularly vulnerable to entrainment [11], but observed rates of entrainment appear to have little impact on populations due to the usual high abundance of juveniles [14]. Entrainment of resident adult fish has the potential for a greater impact on populations because even relatively low levels of entrainment of large, highly fecund females can reduce population growth and long-term viability, particularly in late-maturing species [15]. Important recreational and aboriginal fishery resources in many reservoirs of North America rely on such late-maturing species (for example, walleye, Sander vitreus; burbot, Lota lota; bull trout Salvelinus confluentus), making efforts to reduce entrainment of adult resident fish a crucial factor for future sustainability. Detailed knowledge of adult resident fish behavior near turbine intakes therefore can help regulators and hydropower operators to identify and implement approaches to reduce entrainment.
In this study, we investigated fine-scale behavior associated with entrainment risk of adult resident fish in a large hydropower reservoir (Kinbasket Reservoir, British Columbia, Canada, Figure 1). We focused on bull trout (Salvelinus confluentus), which is a char native to western Canada and the northwestern United States. Bull trout inhabit cold-water rivers, lakes and reservoirs and exhibit a range of life history forms (resident, fluvial, adfluvial and anadromous) [16]. The species remains active even in water temperatures below 2°C, possesses low thermal optima for growth and is highly sensitive to warm water temperature [17, 18]. As a result of climate change, overfishing and habitat loss, as well as fragmentation and degradation, a number of bull trout populations have been listed as threatened in the United States and designated as of special concern or threatened in three of five biogeographic populations in Canada [19, 20]. Reservoir populations of bull trout are further impacted by entrainment losses of adults, which occur mostly in the fall and winter and at annual rates ranging from 3.5% to 11.3% [21–23]. Seasonality in entrainment risk is most likely associated with temporal changes in physical (for example, turbine operations, water temperature) and biological (for example, prey distribution) factors affecting behavior and forebay use [11, 23].

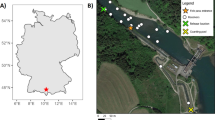
Study site and acoustic telemetry array. (a) Kinbasket Reservoir with inset showing the province of British Columbia, Canada. The arrow in the inset shows the location of the reservoir. (b) Forebay of Mica Dam showing the locations (surveyed once) of the acoustic telemetry receivers (black circles). The white crosses identify the fixed-position receivers with a beacon tag used to assess the performance of the telemetry positioning system. The black cross within the array denotes the location of the beacon tags whose detections were used to adjust the receivers’ clock drift. The black rectangle and polygon denote, respectively, the top of the powerhouse and part of the dam. The rectangle adjacent to the top of the powerhouse denotes the powerhouse wall. The small white rectangles on the wall denote the intakes, which are numbered 1 to 6 from right to left (only intakes 1 to 4 are operational). The dashed contour lines denote the water depth (in meters) below the surface at high pool. The reservoir is located at Universal Transverse Mercator (UTM) Zone 11.
We applied state-space models to fine-scale acoustic telemetry data to estimate true positions, depth, body temperature and behavioral states of adult bull trout. The resulting estimates were used to investigate the relationship between adult bull trout behavior in the forebay of Kinbasket Reservoir and putative factors influencing their behavior. Behavior was characterized in terms of bull trout residence time and spatial distribution in the forebay, distance to the intakes and two behavioral states based on movement patterns: transiting (fast, directed movement) and exploratory (slow, undirected movement) [24, 25]. Specifically, our objectives were to investigate (1) temporal (diel and seasonal) patterns in bull trout behavior; (2) the association between bull trout behavior and physical (that is, turbine operations, reservoir water elevation) and biological (that is, body temperature) factors; and (3) the behavior of fish preceding their entrainment (should entrainment be observed). We synthesized our results to develop an overall characterization of adult bull trout behavior in relation to entrainment risk.
Results
A total of 85 bull trout were tagged, but only 25 individuals were detected in the forebay for a minimum of 30 minutes (see Methods). Twenty-two of these 25 individuals were detected in only one season, and the remaining three individuals were detected in two seasons. State-space model estimates were obtained for a similar number of individuals across seasons: six individuals in the summer, seven in the fall, seven in the winter and eight in the spring (see examples in Figure 2). However, the total number of positions varied markedly among seasons; they were highest in the winter (8,343, or 51.6% of the total number of 16,184 positions), followed by the fall (5,625, or 34.8%), summer (1,505, or 9.3%) and spring (711, or 4.4%). The median residence time of bull trout in the forebay was 6.3 hours (min–max = 0.6–114.9 hours) in the winter, 2.7 hours (min–max: 1.8–77.8 hours) in the fall, 1.3 hours (min–max: 0.7 - 16.9 hours) in the summer and 0.7 hours (min–max: 0.5–4.7 hours) in the spring.The estimated utilization distributions revealed a marked seasonal pattern of space use within the monitored forebay area. In the spring and summer, bull trout used areas away from the powerhouse more intensively, as indicated by the location of the 50% utilization distribution (Figure 3a and b). In the fall, their 50% utilization distribution extended to the powerhouse (Figure 3c) and was located immediately adjacent to it in the winter (Figure 3d).

Examples of raw position data and state-space model estimates. In (a) to (d), the solid gray lines denote the raw position data. The filled circles denote the state-space model estimates of true positions and associated probability of being in the exploratory state (P exp ). The black rectangle denotes the powerhouse of Mica Dam. The dashed line denotes the waterline at the time the position data were recorded. UTM (Universal Transverse Mercator) Zone 11.

Population-level estimates of utilization distribution for bull trout near the powerhouse of Mica Dam. Data for (a) spring, (b) summer, (c) fall and (d) winter. The black rectangle represents the powerhouse of Mica Dam. The black contour line overlaid on the utilization distribution (UD) denotes the area containing 50% of its volume, and the dashed line denotes the waterline at the maximum reservoir elevation observed in each season. UTM (Universal Transverse Mercator) Zone 11.
Model selection indicated strong support for the relationship between mean three-dimensional distance between fish locations and intakes (D int ) and season (Table 1). Bull trout were typically 57 to 99 m closer to the intakes in the winter than in any of the other seasons (Figure 4a and b). Visual inspection of the D int time series for each track and the fish trajectories indicated that none of the tagged bull trout were entrained (data not shown), with the shortest distance estimated between bull trout and intakes being 23 m and occurring in the winter.

Mean three-dimensional distance between bull trout locations and intakes by season and time of day. (a) and (c) show the raw data, and (b) and (d) show additive effect estimates from the top-ranked generalized additive mixed model. Estimates of additive effects by season in the second-ranked model are similar to those shown in (b). The solid line in (c) is a LOESS (local regression) smoother identifying the trend in the data. The dashed lines in (b) and (d) denote 95% confidence intervals. Degrees of freedom for the smoother shown in (d) are 2.0. D int , Mean three-dimensional distance between bull trout locations and intakes.
Model selection also supported the relationship between D int and time of day (Table 1). However, D int varied by only 20 m over the diel cycle, with slightly reduced D int values occurring between 13:00 and 24:00 (Figure 4c and d). The marginal and conditional R2 values for the D int models with ΔAIC c <2 ranged from 0.22 to 0.23 and 0.31 to 0.32, respectively. None of the models with ΔAIC c <2 included operational discharge or reservoir elevation (Table 1).
The estimated probability of being in the exploratory state (P exp ) progressively increased from spring to winter and was less variable during the winter than in any of the other seasons (Figure 5a). Model selection, however, did not support the relationship between season and P exp (Table 2). The top-ranked model included only body temperature as a predictor of P exp (Table 2). Bull trout were much less likely to be in the exploratory state when their body temperature was around 6°C, with P exp increasing rapidly when body temperature trended toward lower and higher values (Figure 5b and c). Model selection also supported the relationship between P exp and time of day (Table 2). Bull trout were less likely to be in the exploratory state around 10:00 and slightly more likely to be in the exploratory state between 14:00 and 04:00 (Figure 5d and e).

Probability of being in the exploratory state for bull trout by season, body temperature and time of day. (a), (b) and (d) show the raw data, and (c) and (e) show additive effect estimates on the logit scale. The additive effect estimate shown in (c) is from the top-ranked model and is similar to the one estimated by the second-ranked model. The additive effect estimate shown in (e) is from the second-ranked model, as the top-ranked model did not include time of day. The solid lines in (b) and (d) are LOESS (local regression) smoothers identifying the trend in the data. The dashed lines in (c) and (e) denote 95% confidence intervals. Degrees of freedom for the smoothers shown in (c) and (e) are 4.4 and 3.5, respectively. P exp , Probability of being in the exploratory state.
The marginal and conditional R2 values for the P exp models with ΔAIC c <2 ranged from only 0.04 to 0.05 and 0.15 to 0.16, respectively. None of the models with ΔAIC c <2 included operational discharge or season (Table 2). Although the model including body temperature and operational discharge had a ΔAIC c value of virtually 2, the effect of operational discharge was not supported, as its addition resulted in negligible improvement of the model’s log-likelihood compared to the model including only body temperature (Table 2).
Discussion
Our findings revealed marked seasonal changes in adult bull trout behavior in the forebay of Kinbasket Reservoir, which helps to explain the increased entrainment rates in the fall and winter observed previously [23]. Specifically, we observed increased residence time (see also [23]) and proximity to intakes in the fall and particularly in the winter, as well as slow, undirected movement in the forebay when body temperatures were low (mostly in the winter). Although turbine operations are maximized during the fall and winter at Mica Dam [26], we did not find any evidence that operational discharge directly influenced adult bull trout behavior in the forebay. This indicates that other factors related to seasons may be associated with the seasonality observed in bull trout behavior.
The forebay of hydropower reservoirs may have high food density for adult bull trout during the late fall and winter. Indeed, kokanee (nonanadromous form of Oncorhynchus nerka), one of the main prey for overwintering bull trout [27], congregate near the dams of some hydropower reservoirs in the winter [28, 29]. In Revelstoke Reservoir (British Columbia, Canada), the density of kokanee near the dam increases particularly during periods of prolonged turbine operation in the winter, possibly due to advection [29]. Furthermore, kokanee usually occur in habitats with high light intensity during the winter [30, 31]. Relatively high light intensity may occur near the powerhouse of hydropower reservoirs in the late fall and winter due to the effect of turbine-induced flows in preventing or inhibiting formation of thick ice cover.
Our finding that P exp increased with decreasing body temperatures provides additional indirect evidence that bull trout may forage near the powerhouse in the late fall and winter. The exploratory behavior is defined by slow movements and frequent turns, which are characteristic movement patterns of animals conducting area-restricted search and foraging [24, 32]. Furthermore, the restricted movement of bull trout near the powerhouse is consistent with a sit-and-wait foraging strategy for capturing invertebrates and fish moving with the direction of turbine-induced flows—a behavior similar to that observed in stream-dwelling salmonids (for example, [33, 34]). Such a sit-and-wait behavior was observed in adult bull trout preying on sockeye salmon smolts migrating past counting fences in the Chilko River, British Columbia, Canada (N Furey, University of British Columbia, personal communication).
The relationship between P exp and body temperature in bull trout may also reflect the bell-shaped thermal dependency of aerobic scope for activity in fishes [35], though the exact relationship between aerobic scope and temperature has not yet been examined in bull trout. However, on the basis of the identified relationship between P exp and body temperatures, it is possible that aerobic scope to sustain fast movements (that is, relatively low P exp ) is highest around 6°C and decreases as body temperature trends toward 0°C or 12°C, leading to a reduction in swim speeds (that is, relatively high P exp ). Alternatively, the increased P exp toward 12°C may be related to a reduction in activity that would facilitate food assimilation and growth at warmer temperatures [36–38].
We also found that bull trout behavior varied slightly on a diel basis. Although diel patterns in activity should be expected to vary seasonally due to changes in the photoperiod [39], we could not evaluate interactions between time of day and season on D int and P exp (see Methods). However, because the majority of our data were recorded in the fall and winter, we believe that the uncovered diel patterns are more representative of bull trout activity in these seasons. The small variation (about 20 m) in D int over the day may be partly associated with bull trout diel vertical migration, which occurs throughout most of the year, including the fall and winter [40]. Bull trout diel vertical migration in winter has been attributed to foraging for kokanee [40], which also exhibit diel vertical migration in that season [31].
Our finding that P exp was higher over most of the night hours (corresponding to fall and winter) may indicate a reduction in bull trout activity when it is dark. However, as mentioned above, the exploratory state may also indicate foraging behavior. Interestingly, the increase in P exp at night coincides with the time that both kokanee and bull trout are located in shallow water [40, 41]. Indeed, diel activity of fishes is linked to the activity of their prey [39], and studies conducted on stream-dwelling bull trout during periods of cold water temperature (late fall to early spring) have shown that individuals emerge from cover at night and feed in the dark ([42, 43] and N Furey, University of British Columbia, personal communication).
Although we were able to detect relationships between bull trout D int and P exp and time of day, season and body temperature, a large proportion of the variability in our data (as measured by conditional R2) could not be explained by our predictor variables and individual random effects, particularly in the P exp model. The low ability of our models to explain the variability in the data is partly related to the stochastic component that underlies animal movements [44]. Furthermore, animal movement results from complex interactions between an individual’s internal state and environmental conditions [45], which can change at fine spatiotemporal scales. For example, turbine operations generate complex water flows over three-dimensional space in the forebay of hydropower reservoirs [46]. It is possible that bull trout moving in such an environment respond to fine spatiotemporal changes in flow properties (for example, velocity, turbulence, flow direction) that are not detected by relating behavior to total operational discharge. Indeed, the spatiotemporal variability in the characteristics of water and air flow helps to explain the trajectory of animals moving in fluids, though most work done to date has been focused on migrating animals [10, 47, 48].
We did not detect bull trout approaching or moving into the intakes in this study, though adult bull trout entrainment is known to occur through Mica Dam in the fall and winter [23]. The closest distance we detected bull trout from an intake was 23 m—a distance where water velocities are <0.2 m/s during turbine operations [49]. We do not consider these velocities challenging for adult bull trout, because even juvenile bull trout (11 to 19 cm) have mean critical swimming speeds >0.48 m/s and critical swim speed is positively correlated with body length [50]. Bull trout may have avoided approaching the flow field near the turbine intakes of Mica Dam, but we cannot exclude the possibility that our telemetry system was unable to position fish at distances <23 m from the intakes during turbine operations. Evidence for this was observed in our assessment of the telemetry system performance in positioning the receiver beacon tags (Additional file 1). During the fall and winter, the median efficiency of the telemetry system in positioning the receiver beacon tags located on the powerhouse wall was nearly 0%, with efficiency decreasing with total operational discharge. In contrast, median efficiency in the fall and winter was 16% and 73%, respectively, for the receiver beacon tag located about 275 m from the powerhouse and did not decrease with increases in operational discharge. The low efficiency in positioning the receiver beacon tags, and possibly fish, near the intakes during turbine operations may be related to reduced propagation and/or integrity of the acoustic signal in the accelerating flow field close to the intakes [51].
Conclusions
Our findings indicate that increased entrainment risk of adult bull trout in the fall and winter is related to a combination of maximization of turbine operations in these seasons with concomitant changes in behavioral attributes, such as increased residence and proximity of bull trout to the intakes (presumably for foraging on kokanee) and reduced movement (perhaps limiting escape responses to accelerating water flow) during periods of cold water temperatures. Therefore, it would be prudent to explore mitigation measures, such as operating deterrent devices (for example, strobe lights, sound, screens), to prevent bull trout from approaching and becoming entrained at hydropower intakes during the fall and winter. These approaches would likely benefit other resident fishes (for example, kokanee) at risk of and impacted by entrainment, but would be especially important for bull trout, given that the viability of their populations can be substantially reduced by losses of adults [16, 52]. The results of this study also show how acoustic telemetry and state-space models can be combined to understand and categorize fish behavior in reservoirs and, more generally, in other environments with fluctuating water levels.
Methods
Study site
This study was conducted in Kinbasket Reservoir (52°8′ N, 118°28′ W), which is located in the Kootenay Mountain Region of British Columbia, Canada, and was formed by the impoundment of the Canoe and Columbia Rivers with the construction of the Mica Dam in 1973 (Figure 1a). The reservoir is large (43,200 ha), snowmelt-fed and oligotrophic, with steep, rocky shorelines, sand, rock and mud substrates. At its highest elevation (high pool, 755 m), the reservoir has a mean depth of 57 m and maximum depth of about 190 m [53]. Surface water temperatures in the reservoir range from 2°C to 15°C in early spring, with summer surface temperatures typically between 12°C and 18°C [54]. From midsummer to early fall, a linear thermal gradient is usually formed in the reservoir, with temperatures decreasing to 4°C at 60 m [55].
The powerhouse at Mica Dam currently consists of four Francis-type turbines, each with a rated maximum discharge of 283 m3/s and capacity of 465 MW [26]. The top of the turbine intakes is located at a depth of about 56 m during high pool. Turbine operation is markedly seasonal, with drawdown starting in late summer or early fall and lasting until early or midspring [26]. As a result of the spring freshet and drawdown, the water surface elevation of the reservoir (hereafter reservoir elevation) varies seasonally by as much as 47 m. The reservoir reaches its lowest elevation (low pool) in the early or midspring and its highest elevation (high pool) by late summer or early fall [26]. The lowest reservoir elevation during our study was about 722 m (top of turbine intakes at a depth of 23 m).
Capture and tagging
Eighty-five adult bull trout were captured by trolling throughout the Kinbasket Reservoir main pool (Figure 1a) in May 2011. Landed fish were anesthetized using clove oil (40 mg of clove oil/L emulsified in 95% ethanol at a 1:9 ratio), measured for total length (cm) and mass (g) and surgically tagged [56] with temperature- and depth-sensing acoustic transmitters (models MM-M-16-33-TP and MM-M-16-50-TP, size 16 × 64 to 81 mm; weight in air 27 to 33 g; fixed signal transmission rate 3, 4 or 5 s; temperature accuracy ±0.8°C; depth accuracy ±3.5 m; frequency 76 kHz; battery life 163 to 433 days; Lotek Wireless, Newmarket, ON, Canada). After surgery, fish were placed in a recovery box filled with ambient reservoir water and released at the capture site once they regained equilibrium (recovery typically took 10 to 15 minutes). The median total length and mass of the tagged bull trout were, respectively, 65.9 cm (min–max: 53.4–84.0 cm) and 2,560 g (min–max = 1,280–5,420 g).
Permits to capture fish were issued by the British Columbia Ministry of Environment (Permit No. CB-PG10- 61414). Tagging protocols were approved by the Carleton University Animal Care Committee.
Tracking and data processing
An array of seven autonomous acoustic telemetry receivers (model WHS 3150; Lotek Wireless) was used to track the movements of tagged bull trout in the immediate vicinity (approximately 300 m) of the powerhouse. The acoustic receivers employ code division multiple access technology, which enables simultaneous tracking of hundreds of tagged animals on a single frequency without code collision, and they operate efficiently under high ambient noise and multipath [57, 58]. Five of the receivers were suspended with aircraft cable attached to the log boom (n = 4) and a moored barrel (n = 1) located near the powerhouse (Figure 1b). The weight of the receivers (35 kg) and 15 kg of added weight kept the receivers vertically oriented at a fixed depth of 25 m. The other two receivers were deployed on the powerhouse wall approximately 15 m above turbine intakes 1 and 5 (Figure 1b). These receivers were deployed using custom-made carts that were connected with aircraft cable to electric winches located at the top of the powerhouse. The carts were fitted into grooves running down the powerhouse wall to keep the receivers stationary.
One beacon tag (burst rate of 30 s, referred to as a receiver beacon tag) was attached to each of three receivers, and two other beacon tags (burst rate of 10 or 20 s, referred to as nonreceiver beacon tags) were deployed at a fixed known location within the array (Figure 1b). The detections of the receiver beacon tags were used to assess the performance of the telemetry positioning system (Additional file 1), and the detections of the nonreceiver beacon tags were used to adjust the receivers’ clock drift during data processing.
The detection data were downloaded every 3 to 5 months between May 2011 and October 2012 and used to compute position estimates with ALPS software (Lotek Wireless). Key inputs used for computing fish positions were as follows: the fish and nonreceiver beacon tag detection data; the location of the receivers and nonreceiver beacon tags (surveyed once at the beginning of the study with a differential global positioning system device, model GeoXH handheld GPS; Trimble, Sunnyvale, CA, USA); the depth of the receivers, which was variable only for the ones located on the powerhouse wall, due to changes in reservoir elevation (monitored on-site by BC Hydro, British Columbia, Canada); and the speed of sound in water at a given water temperature. Water temperature was monitored on-site with thermal loggers (model TidbiT v2, accuracy ±0.2°C; Onset Computer Corporation, Bourne, MA, USA).
Data analyses
The telemetry positioning system could not resolve estimates for many of the detections and consequently generated tracks with position estimates at irregular time intervals. In addition, a number of estimated positions were obviously erroneous (for example, fish positioned on land or moving at >10 m/s; see examples in Figure 2). Furthermore, preliminary assessments of the system performance to position the receiver beacon tags revealed that position estimates had both systematic and random errors that increased with decreasing numbers of receivers included in the computation of a position estimate (Additional file 1). Indeed, detection efficiency variability associated with a variety of factors (for example, noise from boat traffic, turbines, rain) and positioning errors are common when using automated positioning systems based on acoustic telemetry [58, 59].
Rather than using ad hoc filters to remove erroneous position estimates, we used state-space models to estimate the true bull trout positions from the observed data. State-space models enable unobserved states (for example, true position, behavioral states) to be estimated from data observed with errors [60, 61]. These models have been used successfully to analyze movement data collected over large temporal (hours to days) and spatial (kilometers) scales from marine and terrestrial animals tracked with geolocation devices (for example, [62–64]). Despite the advantages of state-space models, they have rarely been used to deal with the complex error structure observed in fine-scale position data collected by acoustic telemetry (for example, [65]).
The bull trout movement data were analyzed using the first-difference correlated random walk model with switch between behavioral states (DCRWS) developed previously by Jonsen et al. [25]. The model enabled us to account for errors in the position estimates and missing data and to estimate the bull trout behavioral state associated with each position. Two behavioral states were estimated: transiting and exploratory. The transiting behavior is characterized by relatively fast moves and persistence in direction, and the exploratory behavior is characterized by relatively slow moves and frequent changes in direction [25].
Bull trout elevations (computed by subtracting fish depth from reservoir elevation) and body temperature data were also modeled using state-space models to account for missing data and errors in sensor readings. Modeling sensor data within the state-space framework enabled us to compare bull trout positions and elevation estimates with bathymetry and reservoir elevation data to create a dynamic, three-dimensional land mask, which informed the models of locations to which bull trout could not move. A Bayesian approach was used to fit the state-space models to the data. Detailed descriptions of the model structure, fitting, performance and parameter estimates are included in Additional file 2. Computer codes used to implement the models, as well as an example data set, are provided in Additional file 3.
Before fitting the models, the tracks were split when the time elapsed between two consecutive observations was greater than 5 minutes. We chose a threshold of 5 minutes to avoid unrealistic movement artifacts, such as looping tracks, which we observed when using time thresholds greater than that. These artifacts arise when the model-interpolated locations are insufficiently constrained by data and are a common outcome when no data are available to inform the DCRWS model with the movement of individuals over a long time interval relative to the model time step [64, 66]. From among the resulting tracks, only those that had a minimum duration of 30 minutes were used in the analyses. This filtering did not exclude detections potentially indicating that bull trout were being entrained; only seven excluded detections occurred at a depth <10 m from the top of the turbine intakes and in all cases fish were later detected near (<5 m) the water surface.
In total, the state-space models were fitted to 148 tracks from 25 individuals. The state-space models estimated true position, elevation, body temperature and behavioral state for bull trout at 60-second intervals. This was the smallest time step for which we could adequately fit the models within a reasonable time frame (about 15 days of computing). The proportion of behavioral states estimated as exploratory at each position was computed from 1,000 Markov chain Monte Carlo (MCMC) samples and interpreted as the probability of bull trout being in the exploratory state (P exp ). See Additional file 2 for details on MCMC sampling.
Population-level, season-specific utilization distributions (spring: April through June; summer: July through September; fall: October through December; winter: January through March) were estimated for bull trout using the state-space estimates of true positions. The kernel density estimation method [67] implemented in the R package “adehabitatHR” [68] was used to compute the utilization distributions. The utilization distributions were estimated using 150 positions randomly sampled (with replacement) from each bull trout observed in a season to avoid biases caused by some fish with numerous locations. The utilization distributions gave the probability of encountering a bull trout at a specific location in the forebay. The forebay area most intensively used by bull trout was defined as the area containing 50% of the utilization distribution volume.
Generalized additive mixed models were used to investigate variation in bull trout distance to turbine intakes (mean three-dimensional distance between fish locations and intakes; D int ) and P exp as a function of a number of predictor variables [69]. D int was modeled as a function of season, operational discharge (min-max: 0–1,168 m3/s), reservoir elevation (min-max: 726–754 m), and a smoother for time of day (min-max: 0.1 - 23.99 hours). P exp was modeled as a function of season, operational discharge, and smoothers for time of day and body temperature (min–max: 0.4°C–12.4°C). The analysis was based on logit-transformed P exp values [70].
For both the D int and P exp analyses, models were fitted with all possible combinations of the predictor variables. However, models with season as a predictor variable did not include operational discharge, reservoir elevation (D int ) or body temperature (P exp ), as these variables were collinear with season (variance inflation factor >5; [71]). The models were fitted only with main effects due to the occurrence of nonpositive definite variance–covariance matrices when interactions were included. The same issue occurred when the individual D int and P exp estimates were used in the analyses, but disappeared when the models were fitted to the median of the variables over 15-minute intervals. Fish identity was used as a random effect. Autocorrelation structures of order 1 were used to account for temporal correlations in the model residuals. A variance structure (implemented using the varIdent function in R package “nlme”; [72]) was used to account for the heteroscedasticity associated with seasons [69].
Model selection was conducted using the bias-corrected Akaike Information Criterion (AIC c ) [73], and models were considered to have strong support from the data when they differed in AIC c (ΔAIC c ) from the top-ranked model by <2 units [73]. The fit of the selected models was assessed based on marginal and conditional R2, which estimate the proportion of variability explained by fixed effects (marginal) and both fixed and random effects (conditional) [74]. Data exploration and model assessment were conducted using graphical approaches [69, 71]. Model fitting and selection were conducted in R 3.0.2 [75] using the packages “mgcv” [76] and “AICcmodavg” [77], respectively.
Abbreviations
- AIC c :
-
Bias-corrected Akaike Information Criterion
- ΔAIC c :
-
Difference in bias-corrected Akaike Information Criterion between a given model and the top ranked model
- AR1:
-
Autoregressive model of order 1
- btp:
-
Body temperature
- DCRWS:
-
First-difference correlated random walk model with switch between behavioral states
- D int :
-
Mean three-dimensional distance between fish locations and intakes
- dsg:
-
Operational discharge
- K:
-
Number of parameters in a model
- log(L):
-
Log-likelihood of a model
- P exp :
-
Probability of being in the exploratory state
- rel:
-
Reservoir elevation
- ssn:
-
Season
- tdy:
-
Time of day
- UTM:
-
Universal Transverse Mercator
- wAICc :
-
Weight bias-corrected Akaike Information Criterion.
References
Čada GF: The development of advanced hydroelectric turbines to improve fish passage survival. Fisheries 2001, 26: 14–23.
Smokorowski KE, Bergeron N, Boisclair D, Clarke K, Cooke S, Cunjak R, Dawson J, Eaton B, Hicks F, Katopodis C, Lapointe M, Legendre P, Power M, Randall R, Rasmussen J, Rose G, Saint-Hilaire A, Sellars B, Swanson G, Winfield N, Wysocki R, Zhu D: NSERC’s HydroNet: a national research network to promote sustainable hydropower and healthy aquatic ecosystems. Fisheries 2011, 36: 480–488. 10.1080/03632415.2011.616459
Dadswell MJ, Rulifson RA: Macrotidal estuaries: a region of collision between migratory marine animals and tidal power development. Biol J Linn Soc 1994, 51: 93–113. 10.1111/j.1095-8312.1994.tb00947.x
Skalski JR, Steig TW, Hemstrom SL: Assessing compliance with fish survival standards: a case study at Rock Island Dam, Washington. Environ Sci Policy 2012, 18: 45–51.
Fisheries Act. R.S.C. 1985. c. F-14 (last amended 25 November 2013) http://laws.justice.gc.ca/eng/acts/F-14/
Chen E, LeBlanc P Technical Report. In Report of the June 27–28, 2013 Workshop for the Development of National Guidance for Managing Impacts of Entrainment and Impingement of Fish at Medium and Large Intakes. Richmond Hill, ON, Canada: SENES Consultants; 2013.
Scruton DA, McKinley RS, Kouwen N, Eddy W, Booth RK: Use of telemetry and hydraulic modeling to evaluate and improve fish guidance efficiency at a louver and bypass system for downstream-migrating Atlantic salmon ( Salmo salar ) smolts and kelts. Hydrobiologia 2002, 483: 83–94. 10.1023/A:1021350722359
Brown L, Haro A, Castro-Santos T: Three-dimensional movement of silver-phase American eels in the forebay of a small hydroelectric facility. In Eels at the Edge: Science, Status, and Conservation Concerns. Edited by: Casselman JM, Cairns DK. Bethesda, MD: American Fisheries Society; 2009:277–291.
Keefer ML, Taylor GA, Garletts DF, Helms CK, Gauthier GA, Pierce TM, Caudill CC: High-head dams affect downstream fish passage timing and survival in the Middle Fork Willamette River. River Res Appl 2013, 29: 483–492. 10.1002/rra.1613
Goodwin RA, Nestler JM, Anderson JJ, Weber LJ, Loucks DP: Forecasting 3-D fish movement behavior using a Eulerian–Lagrangian–agent method (ELAM). Ecol Modell 2006, 192: 197–223. 10.1016/j.ecolmodel.2005.08.004
Coutant CC, Whitney RR: Fish behavior in relation to passage through hydropower turbines: a review. Trans Am Fish Soc 2000, 129: 351–380. 10.1577/1548-8659(2000)129<0351:FBIRTP>2.0.CO;2
Federal Energy Regulatory Commission (FERC) (Technical Report, Paper DPR-10). In Preliminary Assessment of Fish Entrainment at Hydropower Projects: A Report on Studies and Protective Measures. Washington, DC: FERC, Office of Hydropower Licensing; 1995. (accessed 15 July 2014) http://www.ferc.gov/industries/hydropower/gen-info/guidelines/preliminary-assessment-fish-entrainment-vol-1.pdf
Skaar D, DeShazer J, Garrow L, Ostrowski T, Thomburg B Technical Report. In Quantification of Libby Reservoir Levels Needed to Maintain or Enhance Reservoir Fisheries: Investigations of Fish Entrainment through Libby Dam, 1990–1994. Kalispell: Montana Department of Fish, Wildlife and Parks; 1996.
Barnthouse LW: Impacts of entrainment and impingement on fish populations: a review of the scientific evidence. Environ Sci Policy 2013, 31: 149–156.
Morris WF, Doak DF: Quantitative Conservation Biology: Theory and Practice of Population Viability Analysis. Sunderland, MA: Sinauer Associates; 2002.
Dunham J, Baxter C, Fausch K, Fredenberg W, Kitano S, Koizumi I, Morita K, Nakamura T, Rieman B, Savvaitova K, Stanford J, Taylor E, Yamamoto S: Evolution, ecology, and conservation of dolly varden, white-spotted char, and bull trout. Fisheries 2008, 33: 537–550. 10.1577/1548-8446-33.11.537
Selong JH, McMahon TE, Zale AV, Barrows FT: Effect of temperature on growth and survival of bull trout, with application of an improved method for determining thermal tolerance in fishes. Trans Am Fish Soc 2001, 130: 1026–1037. 10.1577/1548-8659(2001)130<1026:EOTOGA>2.0.CO;2
Gutowsky LFG, Harrison PM, Landsman SJ, Power M, Cooke SJ: Injury and immediate mortality associated with recreational troll capture of bull trout ( Salvelinus confluentus ) in a reservoir in the Kootenay-Rocky Mountain region of British Columbia. Fish Res 2011, 109: 379–383. 10.1016/j.fishres.2011.02.022
United States Fish and Wildlife Service (USFWS): Endangered and threatened wildlife and plants: determination of threatened status for bull trout in the coterminous United States. Final Rule. 50 CFR Part 17. Subpart D—Threatened Wildlife. § 17.44. Special Rules—Fishes. Fed Reg 1999, 64: 58910–58933.
Committee on the Status of Endangered Wildlife in Canada (COSEWIC): Canadian Wildlife Species at Risk. Gastineau, QC, Canada: COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada; 2013. (accessed 15 July 2014) http://www.cosewic.gc.ca/eng/sct0/rpt/csar_e_2013.pdf
Flatter B Technical Report. In Life history and population status of migratory bull trout (Salvelinus confluentus) in Arrowrock Reservoir, Idaho. Boise, Idaho: Department of Fish and Game; 1998.
Salow T, Hostettler L Technical Report. In Movement and Mortality Patterns of Adult Adfluvial Bull Trout (Salvelinus confluentus) in the Boise River Basin Idaho. US Department of the Interior, Bureau of Reclamation; 2004. (accessed 15 July 2014) http://www.usbr.gov/pn/snakeriver/esa/bulltrout/reports/2004-Arrowrockvalveradiotelemetry.pdf
Martins EG, Gutowsky LFG, Harrison PM, Patterson DA, Power M, Zhu DZ, Leake A, Cooke SJ: Forebay use and entrainment rates of resident adult fish in a large hydropower reservoir. Aquat Biol 2013, 19: 253–263. 10.3354/ab00536
Pedersen MW, Patterson TA, Thygesen UH, Madsen H: Estimating animal behavior and residency from movement data. Oikos 2011, 120: 1281–1290. 10.1111/j.1600-0706.2011.19044.x
Jonsen ID, Mills-Flemming J, Myers RA: Robust state-space modeling of animal movement data. Ecology 2005, 86: 2874–2880. 10.1890/04-1852
Hydro BC Technical Report. In Mica Dam: Entrainment Risk Screening. Burnaby, BC, Canada: BC Hydro; 2006.
Beauchamp DA, Van Tassell JJ: Modeling seasonal trophic interactions of adfluvial bull trout in Lake Billy Chinook, Oregon. Trans Am Fish Soc 2001, 130: 204–216. 10.1577/1548-8659(2001)130<0204:MSTIOA>2.0.CO;2
Maiolie M, Elam S (Project 198709900, BPA Report DOE/BP-35167–10). In Kokanee Entrainment Losses at Dworshak Reservoir: Dworshak Dam Impacts Assessment and Fisheries Investigation Project, 1996 Annual Report. Portland, OR: Bonneville Power Administration; 1998. (accessed 15 July 2014) https://pisces.bpa.gov/release/documents/documentviewer.aspx?doc=35167–10;
Dawson J, Parkinson E Technical Report. In Revelstoke Reservoir Kokanee Behavior and Entrainment Rate Assessment. Seattle, WA: Biosonics; 2013.
Steinhart GB, Wurtsbaugh WA: Winter ecology of kokanee: implications for salmon management. Trans Am Fish Soc 2003, 132: 1076–1088. 10.1577/T02-135
Steinhart GB, Wurtsbaugh WA: Under-ice diel vertical migrations of Oncorhynchus nerka and their zooplankton prey. Can J Fish Aquat Sci 1999, 56: 152–161. 10.1139/f99-214
Benhamou S, Bovet P: How animals use their environment: a new look at kinesis. Anim Behav 1989, 38: 375–383. 10.1016/S0003-3472(89)80030-2
Bachman RA: Foraging behavior of free-ranging wild and hatchery brown trout in a stream. Trans Am Fish Soc 1984, 113: 1–32. 10.1577/1548-8659(1984)113<1:FBOFWA>2.0.CO;2
Piccolo JJ, Hughes NF, Bryant MD: Water velocity influences prey detection and capture by drift-feeding juvenile coho salmon ( Oncorhynchus kisutch ) and steelhead ( Oncorhynchus mykiss irideus ). Can J Fish Aquat Sci 2008, 65: 266–275. 10.1139/f07-172
Fry FEJ: Effects of the environment on animal activity. Publ Ontario Fish Res Lab 1947, 68: 1–52.
Larsson S, Berglund I: The effect of temperature on the energetic growth efficiency of Arctic charr ( Salvelinus alpinus L.) from four Swedish populations. J Therm Biol 2005, 30: 29–36. 10.1016/j.jtherbio.2004.06.001
Armstrong JB, Schindler DE, Ruff CP, Brooks GT, Bentley KE, Torgersen CE: Diel horizontal migration in streams: juvenile fish exploit spatial heterogeneity in thermal and trophic resources. Ecology 2013, 94: 2066–2075. 10.1890/12-1200.1
Mesa MG, Weiland LK, Christiansen HE, Sauter ST, Beauchamp DA: Development and evaluation of a bioenergetics model for bull trout. Trans Am Fish Soc 2013, 142: 41–49. 10.1080/00028487.2012.720628
Helfman GS: Fish behaviour by day, night and twilight. In Behavior of Teleost Fishes. 2nd edition. Edited by: Pitcher TJ. London: Chapman & Hall; 1993:479–512.
Gutowsky LFG, Harrison PM, Martins EG, Leake A, Patterson DA, Power M, Cooke SJ: Diel vertical migration hypotheses explain size-dependent behaviour in a freshwater piscivore. Anim Behav 2013, 86: 365–373. 10.1016/j.anbehav.2013.05.027
Levy DA: Acoustic analysis of diel vertical migration behavior of Mysis relicta and kokanee ( Oncorhynchus nerka ) within Okanagan Lake, British Columbia. Can J Fish Aquat Sci 1991, 48: 67–72. 10.1139/f91-010
Thurow RF: Habitat utilization and diel behavior of juvenile bull trout ( Salvelinus confluentus ) at the onset of winter. Ecol Freshw Fish 1997, 6: 1–7. 10.1111/j.1600-0633.1997.tb00136.x
Jakober MJ, McMahon TE, Thurow RF: Diel habitat partitioning by bull charr and cutthroat trout during fall and winter in Rocky Mountain streams. Environ Biol Fishes 2000, 59: 79–89. 10.1023/A:1007699610247
Turchin P: Quantitative Analysis of Movement: Measuring and Modeling Population Redistribution in Animals and Plants. Sunderland, MA: Sinauer Associates; 1998.
Nathan R, Getz WM, Revilla E, Holyoak M, Kadmon R, Saltz D, Smouse PE: A movement ecology paradigm for unifying organismal movement research. Proc Natl Acad Sci USA 2008, 105: 19052–19059. 10.1073/pnas.0800375105
Shammaa Y, Zhu DZ, Rajaratnam N: Flow upstream of orifices and sluice gates. J Hydraul Eng 2005, 131: 127–133. 10.1061/(ASCE)0733-9429(2005)131:2(127)
McElroy B, DeLonay A, Jacobson R: Optimum swimming pathways of fish spawning migrations in rivers. Ecology 2012, 93: 29–34. 10.1890/11-1082.1
Safi K, Kranstauber B, Weinzierl R, Griffin L, Rees EC, Cabot D, Cruz S, Proaño C, Takekawa JY, Newman SH, Waldenström J, Bengtsson D, Kays R, Wikelski M, Bohrer G: Flying with the wind: scale dependency of speed and direction measurements in modelling wind support in avian flight. Mov Ecol 2013, 1: 4. 10.1186/2051-3933-1-4
Langford MT, Robertson CB, Zhu DZ Technical Report. In Mica Dam Fish Entrainment Hydraulics: Computational Fluid Dynamic Modeling and Field Study. Edmonton, AB, Canada: University of Alberta; 2012.
Mesa MG, Weiland LK, Zydlewski GB: Critical swimming speeds of wild bull trout. Northwest Sci 2004, 78: 59–65.
Pincock DG, Johnston SV: Acoustic telemetry overview. In Telemetry Techniques: A User Guide for Fisheries Research. Edited by: Adams NS, Beeman JW, Eiler JH. Bethesda, MD: American Fisheries Society; 2012:305–337.
Post JR, Mushens C, Paul A, Sullivan M: Assessment of alternative harvest regulations for sustaining recreational fisheries: model development and application to bull trout. North Am J Fish Manag 2003, 23: 22–34. 10.1577/1548-8675(2003)023<0022:AOAHRF>2.0.CO;2
Sebastian D, Scholten G, Addison D, Labelle M, Green D (Stock Management Unit Report No 1). In Results of the 1991–1993 Hydroacoustic Surveys at Mica and Revelstoke Reservoirs. Victoria, BC, Canada: British Columbia Ministry of Environment; 1995.
Bray K Study Report CLBMON-3. In Columbia River Project Water Use Plan: Kinbasket and Revelstoke Reservoirs Ecological Productivity Monitoring: Progress Report Year 3 (2010). Burnaby, BC, Canada: BC Hydro, Environment; 2011. (accessed 15 July 2014) http://www.bchydro.com/content/dam/hydro/medialib/internet/documents/planning_regulatory/wup/southern_interior/2012q1/clbmon-3_yr3_2012–01–01.pdf
Robertson CB: Forebay thermal dynamics at hydropower facilities on the Columbia River system. University of Alberta, Department of Civil and Environmental Engineering; 2012. MSc thesis
Wagner GF, Cooke SJ, Brown RS, Deters KA: Surgical implantation techniques for electronic tags in fish. Rev Fish Biol Fish 2011, 21: 71–81. 10.1007/s11160-010-9191-5
Niezgoda G, Benfield M, Sisak M, Anson P: Tracking acoustic transmitters by code division multiple access (CDMA)-based telemetry. Hydrobiologia 2002, 483: 275–286. 10.1023/A:1021368720967
Cooke SJ, Niezgoda GH, Hanson KC, Suski CD, Phelan FJS, Tinline R, Philipp DP: Use of CDMA acoustic telemetry to document 3-D positions of fish: relevance to the design and monitoring of aquatic protected areas. Mar Technol Soc J 2005, 39: 31–41. 10.4031/002533205787521659
Kessel ST, Cooke SJ, Heupel MR, Hussey NE, Simpfendorfer CA, Vagle S, Fisk AT: A review of detection range testing in aquatic passive acoustic telemetry studies. Rev Fish Biol Fish 2014, 24: 199–218. 10.1007/s11160-013-9328-4
Patterson TA, Thomas L, Wilcox C, Ovaskainen O, Matthiopoulous J: State-space models of individual animal movement. Trends Ecol Evol 2008, 23: 87–94. 10.1016/j.tree.2007.10.009
Jonsen ID, Basson M, Bestley S, Bravington MV, Patterson TA, Pedersen MW, Thomson R, Thygesen UH, Wotherspoon SJ: State-space models for bio-loggers: a methodological road map. Deep Sea Res Part II 2013, 88–89: 34–46.
Morales JM, Haydon DT, Frair J, Holsinger KE, Fryxell JM: Extracting more out of relocation data: building movement models as mixtures of random walks. Ecology 2004, 85: 2436–2445. 10.1890/03-0269
Jonsen ID, Myers RA, James MC: Robust hierarchical state-space models reveal diel variation in travel rates of migrating leatherback turtles. J Anim Ecol 2006, 75: 1046–1057. 10.1111/j.1365-2656.2006.01129.x
Block BA, Jonsen ID, Jorgensen SJ, Winship AJ, Shaffer SA, Bograd SJ, Hazen EL, Foley DG, Breed GA, Harrison A-L, Ganong JE, Swithenbank A, Castleton M, Dewar H, Mate BR, Shilinger GL, Schaefer KM, Benson SR, Weise MJ, Henry RW, Costa DP: Tracking apex marine predator movements in a dynamic ocean. Nature 2011, 475: 86–90. 10.1038/nature10082
Baktoft H, Aarestrup K, Berg S, Boel M, Jacobsen L, Jepsen N, Koed A, Svendsen JC, Skov C: Seasonal and diel effects on the activity of northern pike studied by high-resolution positional telemetry. Ecol Freshw Fish 2012, 21: 386–394. 10.1111/j.1600-0633.2012.00558.x
Bailey H, Shillinger G, Palacios D, Bograd S, Spotila J, Paladino F, Block B: Identifying and comparing phases of movement by leatherback turtles using state-space models. J Exp Mar Bio Ecol 2008, 356: 128–135. 10.1016/j.jembe.2007.12.020
Worton BJ: Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70: 164–168. 10.2307/1938423
Calenge C: The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecol Modell 2006, 197: 516–519. 10.1016/j.ecolmodel.2006.03.017
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM: Mixed Effects Models and Extensions in Ecology with R. New York: Springer; 2009.
Warton DI, Hui FKC: The arcsine is asinine: the analysis of proportions in ecology. Ecology 2011, 92: 3–10. 10.1890/10-0340.1
Zuur AF, Ieno EN, Elphick CS: A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 2010, 1: 3–14. 10.1111/j.2041-210X.2009.00001.x
Pinheiro J, Bates D, DebRoy S, Sarkar D: R Development Core Team: nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1–104. (accessed 15 July 2014) [http://cran.r-project.org/web/packages/nlme/index.html]
Burnham KP, Anderson DR: Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach. 2nd edition. New York: Springer; 2002.
Nakagawa S, Schielzeth H: A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol Evol 2013, 4: 133–142. 10.1111/j.2041-210x.2012.00261.x
R Development Core Team: R: A Language and Environment for Statistical Computing. (accessed 15 July 2014) http://www.r-project.org/
Wood S: Generalized Additive Models: An Introduction with R. Boca Raton, FL: Chapman & Hall/CRC; 2006.
Mazerolle MJ: AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R package version 1.33 (accessed 15 July 2014) [http://CRAN.R-project.org/package=AICcmodavg]
Acknowledgements
This paper is dedicated to the late Paul Higgins, a talented fisheries professional and friend who was a strong supporter of this work before his premature passing. We thank Jamie Tippe, Juan Molina, Jayme Hills, Taylor Nettles, Lisa Donaldson, Glenn Crossin, Jeff Nitychoruk, Jason Thiem and Sean Landsman for helping in the field; Nathan Furey for testing the codes provided in Additional file 3; and two anonymous reviewers for providing helpful comments to improve the manuscript. This research was funded by the Natural Sciences and Engineering Research Council of Canada (NSERC HydroNet and a Collaborative Research and Development Grant; discovery grants to MP and SJC; and a NSERC OTN Research Network grant to SJC, JEMF and IDJ), BC Hydro and the Centre for Expertise on Hydropower Impacts of Fish and Fish Habitat of Fisheries and Oceans Canada. SJC is supported by the Canada Research Chairs Program. Some of the telemetry infrastructure was provided by the Canada Foundation for Innovation and the Ontario Ministry of Research and Innovation. We also thank Karen Smokorowski, Daniel Boisclair and Shannon O’Connor for supporting the project.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
EGM conducted fieldwork, data management and analyses and drafted the manuscript. LFGG and PMH conducted fieldwork. JEMF and IDJ helped to develop the extended version of the DCRWS state-space model. DZZ, AL, DAP, MP and SJC conceived and designed the study. All authors contributed to the drafting of the manuscript and read and approved the final manuscript.
Electronic supplementary material
40317_2014_33_MOESM2_ESM.docx
Additional file 2: Description of state-space model structure, fitting, performance and parameter estimates. (DOCX 4 MB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Martins, E.G., Gutowsky, L.F.G., Harrison, P.M. et al. Behavioral attributes of turbine entrainment risk for adult resident fish revealed by acoustic telemetry and state-space modeling. Anim Biotelemetry 2, 13 (2014). https://doi.org/10.1186/2050-3385-2-13
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/2050-3385-2-13