
Abstract
Mastitis is still one of the three main diseases that affects the profitability of dairy farmers. Despite the implementation of the five-point mastitis control programme in the early 1970 s, the incidence in the UK has not reduced dramatically over the past 10 years. A review of the scientific literature indicates that there is a link between nutrition and mastitis in the dairy cow. The major impact of nutrition on udder health is via suppression of the immune system. Cows in negative energy balance are at a higher risk of ketosis and clinical ketosis is associated with a two-fold increase in the risk of clinical mastitis. Trace minerals and vitamins that can affect udder health are selenium and vitamin E, copper, zinc, and vitamin A and β-carotene.
Similar content being viewed by others

Introduction
Mastitis is still one of the three main diseases that affect the profitability of dairy farmers - lameness and fertility being the other two. Whilst what the farmer sees is clinical mastitis (treatment costs plus withheld milk) it is subclinical mastitis that can be responsible for the major costs (decreased milk production, cell count penalty payments).
Mastitis control measures were implemented in the early 1970 s following the development of the five-point control programme by the National Institute of Research in Dairying. These included:
-
Annual milking machine test;
-
Post-milking teat disinfection;
-
Dry cow therapy on all cows;
-
Correct treatment of clinical cases and record keeping; and,
-
Culling chronically infected cows.
A significant reduction in the incidence of clinical mastitis was seen initially, but the incidence in the UK has not reduced dramatically over the past 10 years - 47 cases per 100 cows per year based on estimates from historic farm records [3]. However, the major causes of mastitis are now Escherichia coli and Streptococcus uberis (Figure 1). The five-point control programme was effective against those pathogens that were spread from cow to cow (Staphylococcus aureus, Streptococcus agalactiae and Streptococcus dysgalactiae) but had little effect against the environmental pathogens (E. coli and S. uberis).

Consequently, there are two further points that need to be added to the five-point control programme:
-
Good teat preparation prior to milking to ensure teats are clean and dry; and,
-
Good housing and bedding management.
Data collected from two large commercial Holstein dairy herds in New York State [41] indicated that the majority of cases of clinical mastitis were detected during the first week of lactation, so it is imperative that the immune system is functioning properly at this time of high stress for the cow.
Defences of the mammary gland
The teat end is the first barrier against invading pathogens. The anatomical and physical characteristics of the teat canal (tightness of closure and keratin lining) inhibit penetration of udder pathogens. Approximately 40% of the keratin lining is removed at each milking [2] and, therefore, it requires constant regeneration. Consequently, it is important to ensure that there is closure of the teat canal post-milking.
After bacteria breach the teat end, they are taken up and destroyed by leukocytes and leukocyte chemotaxis is one of the major factors involved in migration of these cells towards the centre of inflammation. Several experimental-infection studies have shown a strong relationship between early leukocyte influx and outcome of infection [13, 18, 4].
Nutrition
Feeding should always be aimed at meeting the nutritional requirements of the dairy cow during the various stages of lactation cycle. During the transition period, the dairy cow can experience negative energy balance, which can have a major impact on the immune response.
It is well known that cows in negative energy balance are at a higher risk of ketosis. Clinical ketosis is associated with a two-fold increase in the risk of clinical mastitis [25] and the concentration of beta-hydroxybutyric acid (BHBA) showed a strong positive correlation to the severity of induced E. coli mastitis [19].
In a review of hyperketonemia and the impairment of udder defence, Suriyasathaporn et al. [38] postulated that cows experiencing negative energy balance show an impairment of udder defence mechanisms. Possible explanations for these effects reduced capacity for phagocytosis by polymorphonuclear neutrophils (PMN) and macrophages, and decreased generation of chemoattractant for blood leukocyte migration into the infected gland. Furthermore, using an in vitro chemotaxis assay it was found that PMN migration was faster for cells from cows with low serum BHB levels than for cells from cows with high BHB levels in their blood. If cells move slowly, they are out competed by bacteria, resulting in clinical mastitis [37] (Figure 2).

In a Canadian study, 28.6% of cows with pre-partum ketosis subsequently developed clinical mastitis compared with 8.7% of cows without pre-partum ketosis [20].
Few studies have investigated associations between clinical mastitis and feeding. However, a Swedish observational study included feeding factors among risk factors for veterinary treated clinical mastitis (VTCM) [1]. The study comprised 158 high yielding herds with low somatic cell count (SCC) and a high (27.3-67.5 cases/100-cow years) or low (0.0-10.8 cases/100-cow years) incidence of VTCM. Feeding factors associated with VTCM of first parity cows were amounts of concentrate given in the period around parturition and feeding related diseases (ketosis, laminitis, and udder-thigh dermatitis caused by udder oedema).
Trace minerals and vitamins
Trace minerals and vitamins that can influence udder health include selenium (Se) and vitamin E, copper, zinc, and Vitamin A and β-carotene.
Vitamin E and selenium
Vitamin E and selenium (Se) are integral components of the antioxidant defence of tissues and cells. Cattle consuming stored forages are likely to be low in vitamin E, unless supplemented, and vitamin E deficiencies are frequently observed in peripartum dairy cows. Pavalata et al. [26] found that about 50% of dairy cows sampled in the Czech Republic had marginal or deficient Se status. Other studies in Slovenia, Germany and Ireland have reported similar findings with the majority of dairy cows being marginal or deficient in Se [22, 12, 43].
Vitamin E is the most important lipid-soluble antioxidant and the biologically most active form is, α-tocopherol [28]. Vitamin E is an integral component of all lipid membranes and has a role in protecting lipid membranes from attack by high tissue concentrations of reactive oxygen species [29]. Selenium is an essential micronutrient present in tissues throughout the body and is important physiologically because it is an integral component of the enzyme glutathione peroxidase [8, 9].Glutathione peroxidase functions in the cytosol of the cell and vitamin E within lipid membranes.
Vitamin E and Se deficiency leads to impaired PMN activity. Dietary supplementation of cows with Se and Vitamin E results in a more rapid PMN influx into milk following intramammary bacterial challenge and increased intracellular killing of ingested bacteria by PMN [34], as well as lowering the frequency and shortening the duration of clinical mastitis [33]. Cows supplemented with 740 IU of vitamin E throughout the dry period had a 37% lower incidence of clinical mastitis during the next lactation compared to cows fed ad libitum haylage providing an estimated 320 IU vitamin E per cow per day. Injection of 0.1 mg Se/kg body weight 21 days before calving had no effect on incidence of clinical mastitis. However, cows supplemented with both Vitamin E and Se had shorter duration of clinical signs than cows supplemented with either micronutrient alone (44% and 46%, respectively) [33].
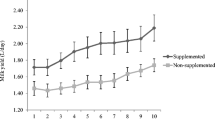
In a study using first lactation heifers [32] diets of heifers were either unsupplemented or supplemented with Se (0.3 ppm) and vitamin E (1,000 IU/d total intake) from 60 days prepartum and continuing throughout lactation. Supplemented heifers had significantly fewer quarters infected at calving, reduced prevalence of infection throughout lactation, fewer cases of clinical mastitis, infections of shorter duration, and lower milk SCC compared with unsupplemented heifers (Figure 3). Weiss et al. [39] found that high serum Se concentrations were associated with reduced rates of clinical mastitis and low bulk tank milk SCC (Figure 4). Erskine et al. [10] found that herds with the low SCCs (less than or equal to 150,000 cells/ml) had significantly higher mean (+/- SEM) blood Se-dependent glutathione-peroxidase (GSH-Px) activity (35.6 +/- 2.95 mU/mg of hemoglobin) than did the herds with the high SCCs (greater than or equal to 700,000 cells/ml) (20.2 +/- 2.38 mU/ mg of Hb). Whole blood concentrations of Se were also significantly higher in the herds with low SCC (0.133 +/- 0.010 mg/ml of blood) than in the herds with high SCC (0.074 +/- 0.007 mg/ml of blood).

The reduction in mastitis variables following dietary supplementation of selenium and Vitamin E in first lactation dairy cows [32]. Used by the permission of the Journal of Dairy Science 75, 1659-1665

Relationship between herd mean Se concentration in plasma and bulk tank somatic cell counts [39]. Used by the permission of the Journal of Dairy Science 73, 381.
Grasso et al. [11] found that cows fed Se-deficient diets, beginning at least 90 days before their first calving, had significantly (P < 0.05) lower blood Se concentration and blood GSH-Px activity than cows fed Se-supplemented diets (2 mg of Se/cow/d as sodium selenite). Milk neutrophils obtained from cows fed Se-supplemented diets killed a significantly (P < 0.05) higher percentage of ingested bacteria (using an in vitro assay) than did neutrophils from cows fed the Se-deficient diet.
Malbe et al. [21] compared the effects of organic selenium (selenised yeast) and sodium selenite in 100 dairy cows where the feed was supplemented with either 0.2 ppm organic selenium or sodium selenite for eight weeks. Blood selenium (geometric mean) increased within this period from the back-ground level (about 5.6 mg/l) to 167 (Se-yeast) and to 91 mg/l (selenite). The bioavailability of yeast selenium was superior to selenite: the relative bioavailability (selenite = 1) of yeast selenium was 1.4 if blood GSH-Px, 1.9 if blood selenium, and 2.7 if milk selenium was used as the response criterion. Selenium-supplementation showed a positive effect on udder health. The percentage of quarters harbouring mastitis pathogens dropped from 22.9 to 13.0 in the Se-yeast group and from 18.4 to 7.4 in the selenite group during the supplementation period. The effect of selenium on mastitis was also reflected as a decrease in the output of milk somatic cells and N-acetyl-beta-D-glucosaminidase (NAGase).
In Norway, as in many European countries, content of Se in soil is low. The Se content in plants is low and cultivated roughage alone cannot supply the animals with recommended daily intake. Se is added as sodium selenite or organic selenium to all types of commercial concentrate. Se enrichment of concentrates does not ensure adequate Se intake of heifers and dry cows fed mainly roughage. Kommisrud et al. [17] carried out a survey of blood Se concentrations in 254 Norwegian Red heifers and dry cows to study possible associations with management, feeding, health and fertility. They found that the relative risk of mastitis treatment in herds with low Se concentration (<0.06 μg/g) was 1.3 to 1.4 times higher during the first 30 days of lactation than in herds with high concentrations of Se (>0.11 μg/g).
Moyo et al. [23] carried out a meta-analysis of 14 papers between 1984 and 2003 to estimate the magnitude and significance of the weighted summarised effect of vitamin E status on udder health. They found vitamin E supplementation was on average associated with a 14% reduction in the risk of intramammary infection (IMI), a reduction in milk SCC by a factor of 0.70 and a 30% decrease in the risk of occurrence of clinical mastitis (CM). These results support the hypothesis that vitamin E supplementation during the dry and early lactation periods is associated with lower IMI, SCC and CM. However, consideration of vitamin E alone would be folly as all the components of the antioxidant system appear to interact.
Copper
Scaletti et al. [30] carried out a study in first lactation heifers to assess a potential role for dietary copper (Cu) in enhancing resistance to E. coli mastitis. Twenty-three primigravid Holstein heifers were maintained on a basal (6.5 ppm copper; -Cu) diet or a diet supplemented (20 ppm) with copper sulphate (+Cu) beginning at 60 days prepartum until 42 days of lactation. They found that milk bacterial count (log10 cfu/ml) was lower at 12, 18, and 48 hours post challenge (day 34 of lactation) for the +Cu group and that the clinical severity score at 24 hours was lower for the +Cu cows. They concluded that copper supplementation reduced the severity of clinical signs during experimental E. coli mastitis but that the duration of mastitis was unaffected. In a separate study Scaletti et al. [31] attempted to determine the effect of copper source on the response to intramammary challenge with E. coli at day 32 of lactation. Twenty primigravid Holstein heifers were maintained on a basal diet (6.5 ppm Cu; CON) or diets supplemented (10 ppm) with copper sulphate (CUS) or Cu proteinate (Bioplex, Alltech Inc.; CUP) beginning 60 days prepartum until 49 days of lactation. Milk bacterial count (log10 cfu/ml) was lower (P < 0.08) for the CUP group at 24 hours compared to CON and CUS groups. Milk production and dry matter intake (DMI) were measured daily following challenge. Milk production and DMI were expressed as percentages of means for 7 days before challenge using the formula [(b/a) × 100 where a = mean value for the 7 days prior to challenge, and b = daily value post challenge] [14]. Dry matter intake following challenge was greater (P < 0.03) on day 4 post-challenge for CUP and CON groups compared to CUS cows and milk production was greater for CUP cows compared to CON (P < 0.12) and CUS (P < 0.05) cows.
Zinc
Zinc is required for keratin formation. Cows receiving supplemental zinc methionine had significantly (P < 0.03) more teat canal keratin [35].
Several studies have examined the effect of supplemental zinc methionine on SCC and in most cases supplementation statistically or numerically reduced SCC. In some studies SCCs were reduced by about 22% [16]. Organic forms of zinc appear to be better absorbed and retained by the animal. Popovic [27] replaced 33% of the supplemental inorganic zinc sulphate with organic zinc (zinc proteinate) from 45 days pre-calving until 100 days post-calving. Cows receiving the organic zinc had significantly lower SCC (62,670 vs 116,440 cells/ml at day 100 respectively).
Spain et al. [36] replaced 50% of the supplemental inorganic zinc with organic zinc (zinc proteinate) and found that cows had significantly fewer new intra-mammary infections.
Vitamin A and β-carotene
Low concentrations of plasma vitamin A (<80 μg/100 ml) and β-carotene (<200 μg/100 ml) were associated with severity of mastitis (based on California Mastitis Test scores) [5]. Dahlquist and Chew [6] carried out a study where cows were fed 53,000 IU of supplemental vitamin A/day, 173,000 IU/day or 53,000 IU/day plus 300 mg of supplemental β-carotene/day starting three weeks before drying-off and continuing through the dry period. Cows fed the vitamin A plus β-carotene treatment had fewer new infections during the early dry period (27% of previously uninfected quarters) than cows fed the other treatments (49% for low vitamin A and 50% for the high vitamin A treatments).
Chew and Johnston [6], carried out another study using the same treatments but supplementation commenced three weeks before calving and continued through 10 weeks of lactation. The vitamin A plus β-carotene treatment reduced mean SCC during weeks two to eight of lactation (85,000 cells/ml compared with 225,000 and 125,000/ml for low and high vitamin A treatments).
In another study, supplementation (50,000 IU/d of vitamin A, 170,000 IU/d of vitamin A or 50,000 IU/d of vitamin A plus 300 mg of β-carotene per cow) did not affect new infections during the dry period, new infections at calving, total new infections during the experiment, and cases of clinical mastitis [24]. A possible reason for the difference between this study and that of Dahlquist and Chew [7] is the β-carotene status of the control cows. At the start of the study by Dahlquist and Chew, mean plasma concentration of β-carotene was 2.5 mg/L whereas in the Oldham et al. study, the mean concentration was 10 mg/L. However, plasma concentrations of β-carotene in dairy cows should be >3 mg/L to optimise udder health [15].
Conclusion
Nutrition can influence the cow's resistance to mastitis. However, it does not influence the exposure of teat ends to pathogens. Mastitis control begins with implementation of the five-point control programme as well as providing a clean dry environment for cows. Ensuring that the cow has adequate energy, minerals and vitamins for optimal milk production is essential for the maintenance of udder health and immune status. However, it should be noted that if mastitis control practices are poor then these will outweigh any potential influence of energy balance, mineral or vitamin status. A holistic approach to mastitis control should be taken and nutritional management is but one part of the control programme.
References
Arvidson AK, Ekman T, Emanuelson U, et al: Feeding factors associated with clinical mastitis of first parity cows. Mastitis in dairy production Current knowledge and future solutions. Edited by: Hogeveen H. 2005, 629-634.
Bitman J, Wood DL, Bright SA, et al: Lipid composition of teat canal keratin collected before and after milking from Holstein and Jersey cows. J Dairy Sci. 1991, 74: 414-420. 10.3168/jds.S0022-0302(91)78184-8.
Bradley AJ, Leach KA, Breen JE, et al: National intervention study of mastitis control in dairy herds in England and Wales. Vet Rec. 2007, 160: 287-93. 10.1136/vr.160.9.287.
Burvenich C, Paape MJ, Hill AW, et al: Role of the neutrophil leucocyte in the local and systemic reactions during experimentally induced E. coli mastitis in cows immediately after calving. Vet Q. 1994, 16: 45-50.
Chew BP, Hollen LL, Hillers JK, et al: Relationship between vitamin A and β-carotene in blood plasma and milk and mastitis in Holsteins. J Dairy Sci. 1982, 65: 2111-2118. 10.3168/jds.S0022-0302(82)82469-7.
Chew BP, Johnston LA: Effects of supplemental vitamin A and β-carotene on mastitis in dairy cows. J Dairy Sci. 1985, 68 (1): 191-10.3168/jds.S0022-0302(85)81199-1.
Dahlquist SP, Chew BP: Effects of vitamin A and β-carotene on mastitis in dairy cows during the early dry period. J Dairy Sci. 1985, 68 (1): 191-
Diplock AT: The role of vitamin E and selenium in the prevention of oxygen-induced tissue damage. Selenium in Biology and Medicine. Edited by: Spallholz JE, Martin JL, Ganther HE. 1981, AVI Publishing, Hartford, CT, 303-
Erskine RJ: Nutrition and mastitis. The Veterinary Clinics of North America: Food Animal Practice. Edited by: Andersen KL, Saunders WB. 1993, Philadelphia, PA, 551-
Erskine RJ, Eberhart RJ, Hutchinson LJ, et al: Blood selenium concentrations and glutathione peroxidase activities in dairy herds with high and low somatic cell counts. J Am Vet Med Assoc. 1987, 190 (11): 1417-21.
Grasso PJ, Scholz RW, Erskine RJ, et al: Phagocytosis, bactericidal activity, and oxidative metabolism of milk neutrophils from dairy cows fed selenium-supplemented and selenium-deficient diets. Am J Vet Res. 1990, 51 (2): 269-74.
Gründer HD, Auer S: Selenium supply of cattle in Hessian herds and possible preventive measures. Tierärztliche Umschau. 1995, 50: 250-255.
Hill AW: Factors influencing the outcome of Escherichia coli mastitis in the dairy cow. Res Vet Sci. 1981, 31: 107-112.
Hogan JS, Weiss WP, Smith KL, et al: Effects of Escherichia coli J5 vaccine on mild clinical coliform mastitis. J Dairy Sci. 1995, 78: 285-290. 10.3168/jds.S0022-0302(95)76636-X.
Jukola E, Hakkarainen J, Saloniemi H, et al: Blood selenium, vitamin E, vitamin A, and β-carotene concentrations and udder health, fertility treatments and fertility. J Dairy Sci. 1996, 79: 838-845. 10.3168/jds.S0022-0302(96)76432-9.
Kincaid RL, Hodgson AS, Riley RE, et al: Supplemtation of diets for lactating with zinc oxide and zinc methionine. J Dairy Sci Suppl. 1984, 1: 103-
Kommisrud E, Østerås O, Vatn T: Blood Selenium Associated with Health and Fertility in Norwegian Dairy Herds. Acta vet scand. 2005, 46: 229-240. 10.1186/1751-0147-46-229.
Kremer WDJ, Noordhuizen-Stassen EN, Lohuis JACM: Host defence mechanisms and bovine coliform mastitis: A review. Vet Q. 1990, 12: 103-113.
Kremer WDJ, Burvenich C, Noordhuizen-Stassen EN, et al: Severity of experimental Escherichia coli mastitis in ketonemic and nonketonemic dairy cows. J Dairy Sci. 1993, 76: 3428-3436. 10.3168/jds.S0022-0302(93)77681-X.
Leslie KE, Duffield TF, Schukken YH, et al: The influence of negative energy balance on udder health. NMC Regional Meeting Proceedings. 2000, 25-33.
Malbe M, Klaassen M, Fang W, et al: Comparisons of selenite and selenium yeast feed supplements on Se-incorporation, mastitis and leucocyte function in Se-deficient dairy cows. Zentralbl Veterinarmed A. 1995, 42 (2): 111-21.
Mee JF, O'Farrell K, Rogers PAM: Trace element deficiency in Irish dairy herds. Irish Grassland and Animal Production Association Journal. 1993, 27: 29-40.
Moyo N, Nielen M, Kruitwagen C, et al: Vitamin E supplementation and udder health: A metaanalysis. Mastitis in dairy production: Current knowledge and future solutions. Edited by: Hogeveen H. 2005, 159-165.
Oldham ER, Eberhart RJ, Muller LD: Effects of supplemental vitamin A and β-carotene during the dry period and early lactation on udder health. J Dairy Sci. 1991, 74: 3775-3781. 10.3168/jds.S0022-0302(91)78569-X.
Oltenacu PA, Ekesbo I: Epidemiological study of clinical mastitis in dairy cattle. Vet Res. 1994, 25: 208-212.
Pavalata L, Illek J, Pechova A, et al: Selenium status of cattle in the Czech Republic. Acta Veterinaria Brno. 2002, 71: 3-8. 10.2754/avb200271010003.
Popovic Z: Performance and udder health status of dairy cows influenced by organically bound zinc and chromium. PhD thesis. 2004, University of Belgrade
Putman ME, Comben N: Vitamin E. Vet Rec. 1987, 121: 541-
Rice DA, Kennedy S: Assessment of vitamin E, selenium and polyunsaturated fatty acid interactions in the aetiology of disease in the bovine. Proc Nutr Soc. 1988, 47: 177-184. 10.1079/PNS19880028.
Scaletti RW, Trammell DS, Smith BA, et al: Role of Dietary Copper in Enhancing Resistance to Escherichia coli Mastitis. J Dairy Sci. 2003, 86: 1240-1249. 10.3168/jds.S0022-0302(03)73708-4.
Scaletti RW, Hamiliton CH, Harmon RJ: Effect of copper source on resistance to coliform mastitis. J Dairy Sci. 2002, 85: 375-
Smith KL, Conrad HR, Amiet BA, et al: Incidence of environmental mastitis as influenced by vitamin E and selenium. Kiel. Milchwirtsch. Forschungsber. 1985, 37: 482-
Smith KL, Harrison JH, Hancock DD, et al: Effect of vitamin E and selenium supplementation on incidence of clinical mastitis and duration of clinical symptoms. J Dairy Sci. 1984, 67: 1293-10.3168/jds.S0022-0302(84)81436-8.
Smith KL, Hogan JS, Weiss WP: Dietary Vitamin E and Selenium Affect Mastitis and Milk Quality. J Anim Sci. 1997, 75: 1659-1665.
Spain JN, Jones CA, Rapp C, et al: The effect of complexed zinc on keratin synthesis in the teat canal and the establishment and severity of experimentally induced E. coli mastitis in dairy cows. Mastitis in dairy production: Current knowledge and future solutions. Edited by: Hogeveen H. 2005, 948-
Spain JN, Stevens BJ, Hardin DK, et al: Effects of Bioplex zinc or zinc oxide on mastitis incidence in lactating dairy cows. J Dairy Sci. 1993, 76: 265-
Suriyasathaporn W, Daemen AJ, Noordhuizen-Stassen EN, et al: Beta-hydroxybutyrate levels in peripheral blood and ketone bodies supplemented in culture media affect the in vitro chemotaxis of bovine leukocytes. Vet Immunol Immunopathol. 1999, 68 (2-4): 177-86. 10.1016/S0165-2427(99)00017-3.
Suriyasathaporn W, Heuer C, Noordhuizen-Stassen EN, et al: Hyperketonemia and the impairment of udder defense: a review. Vet Res. 2000, 31 (4): 397-412. 10.1051/vetres:2000128.
Weiss WP, Hogan JS, Smith KL, et al: Relationships among selenium, vitamin E, and mammary gland health in commercial dairy herds. J Dairy Sci. 1990, 73: 381-10.3168/jds.S0022-0302(90)78684-5.
Wilesmith JW, Francis PG, Wilson CD: Incidence of clinical mastitis in a cohort of British dairy herds. Vet Rec. 1986, 118: 199-204. 10.1136/vr.118.8.199.
Wilson DJ, Gonzalez RN, Hertl J, et al: Effect of clinical mastitis on the lactation curve: a mixed model estimation using daily milk weights. J Dairy Sci. 2004, 87: 2073-2084. 10.3168/jds.S0022-0302(04)70025-9.
Zadoks RN: The role of nutrition in udder health. 2006, High Plains Dairy Conference, Albuquerque, New Mexico, 115-127.
Zust J, Hrovatin B, Simundic B: Assessment of selenium and vitamin E deficiencies in dairy herds and clinical disease in calves. Vet Rec. 1996, 139: 391-394. 10.1136/vr.139.16.391.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
O'Rourke, D. Nutrition and udder health in dairy cows: a review. Ir Vet J 62 (Suppl 4), S15 (2009). https://doi.org/10.1186/2046-0481-62-S4-S15
Published:
DOI: https://doi.org/10.1186/2046-0481-62-S4-S15