
Abstract
Splendid squid (Loligo formosana) is the species available in Thailand and used for local consumption and export. The squid ink is discarded as by-product which can be used as a source of active compounds. Therefore, the aim of this study was to investigate the antioxidant activity of squid ink after melanin removal. The antioxidative activities of melanin-free ink (MFI) from splendid squid were investigated using different in vitro assays. Antioxidative activity of MFI was also determined in different model systems. Thermal stability at 90°C was tested. MFI was subjected to ultrafiltration, and fractions with various molecular weight (MW) ranges were analyzed for antioxidative activities. MFI had DPPH, ABTS radical scavenging activities, and ferric reducing antioxidant power (FRAP) of 179.6 ± 2.1, 957.8 ± 89.3, and 171.2 ± 7.3 μmol TE/g protein, respectively. It also possessed metal chelating activity of 4.0 ± 1.2 μmol EE/g protein. MFI at 500 mg/L could prevent the oxidation of β-carotene-linoleic acid system, but its efficacy was lower than butylated hydroxyl anisole (BHA) at 200 mg/L (p < 0.05). The MFI (100 to 500 mg/L) showed the preventive effect on lipid peroxidation of lecithin liposome system in a dose-dependent manner. When mackerel mince was added with MFI at levels of 100 and 200 mg/kg, lipid oxidation was retarded during ice storage for 15 days as evidenced by the lower peroxide value and thiobarbituric acid-reactive substances, as compared with the control. However, its effectiveness was lower than 200 mg/kg BHA. When MFI was subjected to heat treatment at 90°C for up to 30 min, its DPPH and ABTS radical scavenging activities remained constant, but FRAP decreased within the first 5 min without subsequent changes. When the MFI was separated using ultrafiltration into different fractions (< 3 KDa, 3 to 10 KDa, and >10 KDa), the fractions with MW less than 3 KDa showed the highest antioxidative activities (p < 0.05). MFI, especially its fraction with MW < 3 KDa, from splendid squid could therefore serve as natural antioxidant to retard lipid oxidation in food products.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Lipid oxidation is one of the major causes for deterioration of many food products. It leads to changes in texture, flavor, odor, and quality of foods. Lipid oxidation also causes some health hazards in human beings such as cardiovascular disease, cancer, and neurological disorders as well as aging process (Gulcin 20112012). To prevent or slow down lipid oxidation, several antioxidants including synthetic and natural antioxidants have been widely used. However, synthetic antioxidants are suspected of being toxic upon long-term exposure and are banned in many countries (Madhavi and Salunkhe 1996). As a consequence, natural and safe antioxidants have gained increasing attention. Natural antioxidants from marine resources, especially from the by-products of seafood processing, can be another alternative antioxidant for food applications.
Squid and cuttlefish have become an important fishery product of Thailand as well as other Southeast Asian countries and are mainly exported worldwide (Hoque et al. 2010). During processing, the viscera along with the ink sac are generated as by-products with low market value and can create serious ecological problems and environmental pollution without appropriate management. These by-products can be a promising source of bioactive compounds. The squid ink has been proved to be an alternative medicine and has a wide range of therapeutic applications (McConnell et al. 1994). The growth performance, antioxidant functions, and immunity in growing broiler chickens were affected by squid ink (Liu et al. 2011). The antibacterial activity of the ink from Sepioteuthis lessoniana and Sepia pharaonis against biofilm bacteria was reported (Ramasamy and Murugan 2005). A protein extracted from cuttlefish (S. lessoniana) ink could inhibit the growth of Staphylococcus aureus(Mochizuki 1979). Tyrosinase present in squid ink is known to play a key role in the defense against microbes (Takai et al. 1992). Ink from Sepia officinalis (Lei et al. 2007) and Sepiella inermis (Rajaganapathy et al. 2000) showed antioxidative and antiretroviral activities, respectively. Due to the abundance of ink generated during squid processing, it can be used as the source of active compounds, particularly antioxidants. However, the ink has a black color, which may limit its application. The removal of melanin, a black pigment, prior to utilization should widen the application of ink. Therefore, the present study aimed to investigate antioxidative activity and properties of melanin-free ink from splendid squid (Loligo formosana), the most common squid used for processing in Thailand.
Methods
Chemicals
The chemicals including 2,2-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) diammonium salt (ABTS), 6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox), 2,2-diphenyl-1-picrylhydrazyl (DPPH), 2,4,6-tripyridyl-s-triazine (TPTZ), 1,1,3,3-tetramethoxypropane, and Tween 40 were purchased from Sigma-Aldrich (St. Louis, MO, USA). Linoleic acid, BHA, 2-thiobarbituric acid, and cumene hydroperoxide were procured from Fluka (Buchs, Switzerland). Methanol, ethanol, acetone, chloroform, hydrochloric acid, and ammonium thiocyanate were obtained from Lab-Scan (Bangkok, Thailand). All chemicals were of analytical grade.
Preparation of melanin-free ink
Fresh squids (24 h after capture) were purchased from a local market in Hat Yai, Songkhla, Thailand and transported in ice using a squid/ice ratio of 1:2 (w/w) to the Department of Food Technology, Prince of Songkla University, Hat Yai, within 30 min. Upon arrival, ink sac was separated from the squid by cutting the ink duct, and the ink was squeezed out. The squid ink was diluted tenfold using cold deionized water (4°C). Thereafter, it was subjected to centrifugation at 18,000×g for 30 min at 4°C using a refrigerated centrifuge (Allegra 25 R centrifuge, Beckman Coulter, Palo Alto, CA, USA). The supernatant obtained was referred to as 'melanin-free ink; MFI’. MFI was subjected to analyses.
Determination of protein content
Protein content was determined using the Lowry method (Lowry et al. 1951). Bovine serum albumin was used as a standard.
Determination of in vitro antioxidative activities
DPPH radical scavenging activity
DPPH radical scavenging activity was determined as per the method of Blois (1958) as modified by Binsan et al. (2008). The sample (1.5 mL) was added to 1.5 mL of 0.15 mM DPPH in 95% (v/v) ethanol was added to the sample. The mixture was mixed vigorously using a vortex mixer (model G-560E, Scientific Industries, Inc., Bohemia, NY, USA) and allowed to stand at room temperature in the dark for 30 min. The absorbance of the resulting solution was measured at 517 nm using a spectrophotometer (UV-1800, Shimadzu, Kyoto, Japan). The blank was prepared in the same manner, except that deionized water was used instead of the sample. A standard curve was prepared using Trolox in the range of 0 to 50 μM. The activity was expressed as micromole Trolox equivalents (TE)/g protein.
ABTS radical scavenging activity
ABTS radical scavenging activity was determined following the method of Blois (1958) as modified by Binsan et al. (2008). The stock solutions included 7.4 mM ABTS solution and 2.6 mM potassium persulphate solution. The working solution was prepared by mixing the two stock solutions in equal quantities and allowing them to react for 12 h at room temperature in the dark. The solution was then diluted by mixing 1 mL of ABTS solution with 50 mL of methanol in order to obtain an absorbance of 1.1 ± 0.02 units at 734 nm using a spectrophotometer. Fresh ABTS solution was prepared daily. A sample (150 μL) was mixed with 2,850 μL of ABTS solution and the mixture was left at room temperature for 2 h in the dark. The absorbance was then measured at 734 nm. A standard curve of Trolox ranging from 0 to 500 μM was prepared. The activity was expressed as micromole Trolox equivalents (TE)/g protein.
Ferric reducing antioxidant power
Ferric reducing antioxidant power (FRAP) was assayed according to the method of Benzie and Strain (1996). Stock solutions included 300 mM acetate buffer (pH 3.6), 10 mM TPTZ solution in 40 mM HCl, and 20 mM FeCl3.6H2O solution. A working solution was freshly prepared by mixing 25 mL of acetate buffer, 2.5 mL of TPTZ solution, and 2.5 mL of FeCl3·6H2O solution. The mixed solution was incubated at 37°C for 30 min and was referred to as FRAP solution. A sample (150 μL) was mixed with 2,850 μL of FRAP solution and was kept for 30 min in the dark. The ferrous tripyridyltriazine complex (colored product) was measured by reading the absorbance at 593 nm. The control was prepared in the same manner except that deionized water was used instead of the sample. The standard curve was prepared using Trolox ranging from 0 to 500 μM. The activity was expressed as micromole Trolox equivalents (TE) /g protein.
Chelating activity toward Fe2+
The chelating activity toward Fe2+ was measured by the method of Thiansilakul et al. (2007). The sample (4.7 mL) was mixed with 0.1 mL of 2 mM FeCl2 and 0.2 mL of 5 mM ferrozine. The reaction mixture was allowed to stand for 20 min at room temperature (26°C to 28°C). The absorbance was then read at 562 nm. The control was prepared in the same manner except that deionized water was used instead of the sample. A standard curve (0 to 50 μM EDTA) was prepared. The Fe2+ chelating activity was expressed as EDTA equivalents (μmol EDTA equivalents (EE)/g protein).
Antioxidative effect of MFI in different model systems
β-Carotene-linoleate model system
The antioxidative activity of MFI in β-carotene linoleic acid emulsion model system was determined as described by Binsan et al. (2008). β-Carotene (10 mg) was dissolved in 10 mL of chloroform. Thereafter, the solution (0.2 mL) was added to 20 mg of linoleic acid and 200 mg of Tween 40. Chloroform was then removed by purging with nitrogen. Fifty milliliters of oxygenated deionized water was added to β-carotene emulsion and mixed well. MFI (500 μL) was then mixed with 4.5 mL of oxygenated β-carotene emulsion to obtain the final concentrations of 100, 200, and 500 mg/L. The oxidation of β-carotene emulsion was monitored spectrophotometrically at 470 nm after 0, 10, 20, 30 40, 60, 90, and 120 min of incubation at 50°C in the dark. BHA at 200 mg/L level was also used. The control was prepared by using distilled water instead of MFI in the assay system.
Lecithin liposome system
The antioxidative activity of MFI in lecithin liposome system was determined according to the method of Frankel et al. (1997). Lecithin was suspended in deionized water at a concentration of 8 mg/mL. The mixture was stirred with glass rod, followed by sonicating for 30 min using a sonicating bath (Elmasonic S 30 H, Elma, Germany). The sample (3 mL) was mixed with lecithin liposome (15 mL) to obtain the final concentrations of 100, 200, and 500 mg/L. The liposome suspension was then sonicated for 2 min. To initiate the assay, 20 μL of 0.15 M cupric acetate were added. The mixture was shaken at 120 rpm using a shaker (WNB 14 and SV 1422, Memmert, Germany) at 37°C in the dark for 0, 6, 12, 24, 36, and 48 h. The liposome oxidation was monitored by determining thiobarbituric acid-reactive substances (TBARS). TBARS values were calculated from the standard curve (0 to 3 mg/L malonaldehyde, MDA) and expressed as milligrams of MDA per milliliters of liposome.
Fish mince model system
Preparation of fish mince
Fish mince was prepared according to the method of Kamil et al. (2002) with a slight modification. Mackerel (Rastrelliger kanagurta) with an average weight of 100 to 150 g, off-loaded 24 h after capture, was purchased from the local market in Hat Yai, Thailand. The fish were kept in ice during the transportation. Upon arrival, the fish were washed and dressed, and the meat was separated manually. The fish mince obtained was divided into four portions (100 g each). One portion, without the addition of MFI, was used as the control and 10 mL of deionized water was added instead. Two portions were mixed with 10 mL of MFI to obtain the final concentrations of 100 and 200 mg/kg mince. Another portion was added with BHA to obtain the final concentration of 200 mg/kg mince. The mince was then thoroughly mixed in order to ensure the homogeneous distribution of MFI and BHA in the mince. Different mince samples were placed in polyethylene bag and kept in ice using a mince/ice ratio of 1:2 (w/w). Molten ice was removed every day and the same quantity of ice was replaced. After the designated storage time (0, 3, 6, 9, 12, and 15 days), the samples were taken for analyses of TBARS and peroxide value (PV).
Determination of peroxide value
The peroxide value (PV) was determined according to the method of Richards and Hultin (2000) with a slight modification. The ground sample (1 g) was homogenized at a speed of 13,500 rpm for 2 min in 11 mL of chloroform/methanol (2:1, v/v). The homogenate was then filtered using a Whatman no. 1 filter paper (GE Healthcare UK Limited, Buckinghamshire, UK). Two milliliters of 0.5% NaCl was then added to 7 mL of the filtrate. The mixture was vortexed at a moderate speed for 30 s and then centrifuged at 3,000×g for 3 min at 4°C using a refrigerated centrifuge to separate the sample into two phases. The lower phase (3 mL) was carefully pipetted out, and 2 mL of cold chloroform/methanol (2:1) mixture was added. Then 25 μL of 30% (w/v) ammonium thiocyanate and 25 μL of 20 mM iron(II) chloride were added to the mixture. The reaction mixture was allowed to stand for 20 min at room temperature prior to reading the absorbance at 500 nm. The blank sample was prepared in the same manner except that deionized water was used instead of ferrous chloride. A standard curve was prepared using cumene hydroperoxide at concentrations ranging from 0.5 to 2 mg/L. PV was expressed as milligrams of cumene hydroperoxide per kilogram sample.
Determination of TBARS
The TBARS values of the sample were determined as described by Buege and Aust (1978). A sample (0.5 g) was mixed with 2.5 mL of TBA solution containing 0.375% thiobarbituric acid, 15% trichloroacetic acid, and 0.25 N HCl. The mixture was heated in boiling water for 10 min to develop a pink color, cooled with running tap water, sonicated for 30 min, followed by centrifugation at 5,000×g at 25°C for 10 min. The absorbance of the supernatant was measured at 532 nm. Standard curve was prepared using 1,1,3,3-tetramethoxypropane (malonaldehyde, MAD) at the concentrations ranging from 0 to 10 ppm, and TBARS were expressed as milligram of MAD equivalents per kilogram sample.
Thermal stability of MFI
Two milliliters of MFI solution (2.8 mg protein/mL) was transferred to a screw-capped test tube. The tube was capped tightly and placed in a water bath (90°C) for 0, 5, 10, 15, 20, 25, and 30 min. After heating for designated times, the treated samples were immediately cooled in iced water. The samples were analyzed for DPPH, ABTS radical scavenging activities, and FRAP as previously described. The remaining activities were expressed, relative to that of untreated sample.
Ultrafiltration of MFI
The ultrafiltration of MFI was carried out using a stirred ultrafiltration cell (model 8050, Amicon Bioseparations, Millipore Corporation, Bedford, MA, USA). Initially, the filtration of MFI was done using the membrane with molecular weight (MW) cut-off of 10 KDa. Then the permeate was filtered through the membrane with MW cut-off of 3 KDa. During filtration, the pressure was maintained at 60 psi with continuous stirring at 150 rpm. The fractions obtained were subjected to determination of antioxidative activities as described above.
Statistical analysis
All experiments were run in triplicate using three different lots of samples. Data were subjected to analysis of variance (ANOVA). Comparison of means was carried out by Duncan’s multiple range tests (Steel and Torrie 1980). Statistical analysis was performed using the Statistical Package for Social Science (SPSS 17.0 for Windows, SPSS Inc., Chicago, IL, USA).
Results and discussion
Antioxidative activities of MFI
MFI showed antioxidative activities when tested using several assays as shown in Table 1. MFI was effective in scavenging DPPH and ABTS radicals, in which DPPH and ABTS radical scavenging activities of 179.6 ± 2.1 and 957.8 ± 89.3 μmol TE/g protein were observed, respectively. DPPH and ABTS radical scavenging activities are based on the ability of antioxidants to donate a hydrogen atom or an electron to stabilize radicals by converting them to the non-radical species (Binsan et al. 2008; Chandrasekara and Shahidi 2011). DPPH is a radical having an odd electron and reacts with hydrogen donated from antioxidant. In case of ABTS radical scavenging activity, the effectiveness depends on the molecular weight, the number of aromatic rings, and the nature of hydroxyl groups’ substitution than the specific functional groups (Hagerman et al. 1998). Squid ink was reported to contain l-dopa and dopamine at concentrations of 1.15 and 0.19 mM, respectively (Lucero et al. 1994). The hydroxyl groups of those compounds more likely donated hydrogen atom to the tested radicals. The FRAP of MFI was 171.1 ± 07.3 μmol TE/g protein and had metal chelating activity of 4.0 ± 1.2 μmol EE/g protein. FRAP is generally used to measure the capacity of a substance in reducing TPTZ-Fe(III) complex to TPTZ-Fe(II) complex (Benzie and Strain 1996; Binsan et al. 2008). The result indicated that MFI was able to act as reducing agent which provided electron for stabilization. Additionally, some compounds in MFI could chelate prooxidative metals, thereby lowering or retarding the initiation of lipid oxidation process. The capacity of antioxidant for chelating metals is strongly dependent on the number of hydroxylic groups in ortho-position (Maqsood and Benjakul 2010). Squid ink was reported to function as antioxidant in hyperlipidemia rats and broil chicken (Lei et al. 2007; Liu et al. 2011).
Antioxidative effect of MFI in different model systems
β-Carotene-linoleate model system
MFI showed preventive effect toward oxidation of β-carotene-linoleate model system in a dose-dependent manner (Figure 1a). MFI at the level of 500 mg/L showed the highest activity, followed by 200 and 100 mg/L, respectively. The decrease in A470 indicates the oxidation of β-carotene in the system caused by free radicals from oxidation of linoleic acid (Chandrasekara and Shahidi 2010). When oxidation of linoleic acid occurs, the free radicals formed are able to attack highly unsaturated β-carotene molecules. As a result, chromophore of β-carotene was lost, as indicated by the decrease in orange color (Binsan et al. 2008). In the presence of MFI, β-carotene bleaching was retarded, mainly due to the chain-breaking inhibition of lipid peroxidation by neutralizing linoleic free radical formed. When comparing antioxidative effect of MFI with BHA (200 mg/L), a higher antioxidative of activity in the system was found for BHA. The ability to prevent the bleaching of β-carotene was more likely governed by their amphiphilic properties of amino acid compositions of peptides in MFI. When antioxidative compounds were oriented at linoleic acid/water interface, the antioxidative effect could be maximized (Binsan et al. 2008). Thus, MFI could prevent oxidation of lipids in the emulsion system.

Antioxidative effects of MFI. Antioxidative effects of MFI from splendid squid in β-carotene-linoleate model system (a) and lecithin liposome model system (b). Bars represent the standard deviation (n = 3).
Lecithin liposome model system
The ability of MFI to retard oxidation in the lecithin liposome system at various concentrations is depicted in Figure 1b. The formation of secondary lipid oxidation products in the lecithin liposome was evaluated by TBARS. MFI effectively retarded the oxidation of lecithin liposome system during incubation of 48 h. However, its preventive effect was less than that of BHA (200 mg/L) as indicated by lower TBARS values of system containing BHA. It was noted that the control sample (without MFI or BHA) had the sharp increase in TBARS after 24 h of incubation, suggesting that the oxidation took place to a higher extent. Liposomes are appropriate lipid models to evaluate antioxidative activity in lipid food or lipoprotein particles containing phospholipids (Frankel et al. 1997). The sample added with MFI at a level of 500 mg/L showed the lower TBARS value, compared to those containing MFI at levels of 200 and 100 mg/L. The result suggested that antioxidant activity of MFI in lecithin liposome system was in the dose-dependent manner. Furthermore, hydrophilic domains of peptides or proteins in MFI more likely migrated and localized at head portion of lecithin in liposome, thereby inhibiting oxidation of liposome via radical scavenging mechanism (Table 1). Due to its metal chelating ability, MFI could chelate Cu2+, a prooxidant in the system. As a result, the oxidation of liposome could be impeded with addition of MFI.
Fish model system
The formation of PV in the mackerel mince treated without and with MFI during ice storage is shown in Figure 2a. The PV increased up to day 6 of storage in all samples (p < 0.05), except for the control sample, in which PV still increased until the end of storage (day 15). The sample added with MFI at the level of 100 mg/kg had a slight decrease in PV at day 9, but PV increased thereafter. It was noted that similar PV values were observed between samples added with 100 and 200 mg/kg MFI and 200 mg/kg BHA at day 15 of storage (p > 0.05). The PV values of mince treated with MFI or BHA were lower than that of control. This indicated that MFI acted as an antioxidant and lowered the formation of primary lipid oxidation products.

Effect of MFI on lipid oxidation in mackerel mince during iced storage. Peroxide value (a) and thiobarbituric acid-reactive substances (TBARS) values (b). Bars represent the standard deviation (n = 3).
Changes in TBARS of mackerel mince without and with MFI at the levels of 100 and 200 mg/kg during ice storage are shown in Figure 2b. TBARS values remained unchanged up to 3 days of storage (p > 0.05). The marked increases in TBARS were observed in all samples up to day 12, except for the sample added with 200 mg/kg BHA, which had lower increase in TBARS. There was a slight decrease at day 15. When comparing TBARS values among the samples, the control had higher TBARS value than other samples throughout 15 days of iced storage (p < 0.05). The TBARS values of mince treated with MFI at the level of 200 mg/kg were less than that treated with 100 mg/kg (p < 0.05). Nevertheless, the TBARS values of mince added with MFI at the levels of 100 and 200 mg/kg were higher than those of sample added with BHA at a level of 200 mg/kg. At day 15 of storage, the decreases in TBARS values were obtained. This was plausibly due to the loss of volatile lipid oxidation products. Since MFI exhibited radical scavenging activity and capability of metal chelation (Table 1), it was able to retard the oxidation in fish mince. Mackerel meat contained high fat content (4.54 ± 0.28 g/100 g) (Osman et al. 2001). It was reported that fish meat was rich in polyunsaturated fatty acid (PUFA) (Chantachum et al. 2000). PUFAs are susceptible to oxidation which is associated with rancidity. Additionally, mackerel meat contained a high amount of heme. The heme protein or other iron-containing proteins are denatured with coincidental release of iron as the storage time increased (Benjakul and Bauer 2001). Degradation of protein during extended storage was also associated with the increase in non-heme iron. Denaturation and degradation might favor destruction of heme, thereby enhancing the release of iron. Chaijan et al. (2005) reported that the non-heme iron content increased in mackerel mince during the iced storage. This non-heme iron could act as prooxidant in fish flesh or mince (Benjakul and Bauer 2001). Owing to the chelating ability of MFI (Table 1), it could scavenge Fe2+ or Fe3+ in the mince. As a result, the lipid oxidation could be retarded. Thus, MFI retarded the lipid oxidation in the mince as revealed by the lower TBARS values than that of control.
Thermal stability of MFI
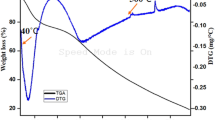
MFI was subjected to heat treatment at 90°C for 30 min, and the remaining DPPH, ABTS radical scavenging activities and FRAP were monitored (Figure 3). DPPH and ABTS radical scavenging activities remained unchanged after heating up to 30 min (p > 0.05). FRAP was decreased when heated for 5 min (p < 0.05). Thereafter, no changes in FRAP were observed (p > 0.05). Some peptides or compounds with FRAP might undergo some aggregation after being heated, while the heat-stable counterparts were resistant to alteration. Zayas (1997) reported that smaller size peptides were more stable to aggregation at high temperatures. The smaller peptides or compounds in MFI could survive during heating and they more likely contributed to prevent lipid oxidation in thermally processed foods.

Thermal stability of MFI from splendid squid at 90°C. Bars represent the standard deviation (n = 3).
Antioxidant activities of fractions with different MWs
DPPH radical scavenging activity of fractions obtained from ultrafiltration with various MW ranges is shown in Figure 4a. The higher activity was observed as MW decreased. The <3 KDa fraction had higher activity than those with MW of 3–10 KDa and >10 KDa, respectively. The <3 KDa fraction had five-fold higher activity than MFI. It was noted that MFI showed similar DPPH radical scavenging activity to that with MW of >10 KDa fraction (p > 0.05). ABTS radical scavenging activity (Figure 4b) had the similar result to that of DPPH radical scavenging activity but ABTS radical scavenging activity of >10 KDa fraction was lower than that of MFI (p < 0.05). The < 3 KDa fraction showed two-fold higher activity than MFI (p < 0.05). FRAP of the fractions decreased as MW of fractions increased (Figure 4c). Nevertheless, FRAP of >10 KDa fraction was higher than that of MFI (p < 0.05). FRAP of < 3 KDa fraction was seven-fold higher than that of MFI. Metal chelating activity of different fractions varied with MW (p < 0.05) (Figure 4d). It was found that chelating activity of >10 KDa fraction was similar to that of MFI (p > 0.05). In general, peptides or proteins with lower MW showed higher antioxidative activity (Bernardini et al. 2011). Liu et al. (2010) reported that <3 KDa fraction from porcine plasma protein hydrolysate exhibited the highest DPPH radical scavenging activity and reducing power. The low MW fraction (< 1 KDa) from the protease N hydrolysate of royal jelly proteins had the highest antioxidative activity (Guo et al. 2009). Park et al. (2001) also noted that <5 KDa hydrolysate from egg yolk protein had the highest antioxidative activity. Wang et al. (2007) reported that the antioxidant activity of wheat gluten hydrolysate UF fraction (<5 KDa) was higher than its original hydrolysate. The <1 KDa fraction of conger eel muscle protein hydrolysates obtained from ultrafiltration exhibited the highest inhibition activity of linoleic acid peroxidation (Ranathunga et al. 2006). Therefore, MW distribution was an important factor governing antioxidative activity of MFI from splendid squid.

DPPH, ABTS, FRAP, and chelating activity of MFI ultrafiltration fractions. DPPH (a), ABTS (b), FRAP (c), and chelating activity toward Fe2+ (d) of ultrafiltration fractions from MFI of splendid squid with different MWs. Bars represent the standard deviation (n = 3). Different letters on the bars for each assay indicate significant differences (p < 0.05).
Conclusions
Squid ink, which is discarded as by-product, could be used as the good source for natural antioxidant after melanin removal. MFI with high thermal stability possessed radical scavenging and metal chelating activities. After ultrafiltration, fraction with lower MW had the greater antioxidative activity. The small MW fraction could be further used as a potential antioxidant.
Abbreviations
- ABTS:
-
2,2-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid)
- BHA:
-
Butylated hydroxyl anisole
- DPPH:
-
2,2-diphenyl-1-picrylhydrazyl
- FRAP:
-
Ferric reducing antioxidant power
- KDa:
-
Kilo Dalton
- MFI:
-
Melanin-free ink
- PUFA:
-
Polyunsaturated fatty acids
- PV:
-
Peroxide value
- SPSS:
-
Statistical Package for Social Science
- TBARS:
-
Thiobarbituric acid-reactive substances.
References
Benjakul S, Bauer F: Biochemical and physicochemical changes in catfish ( Silurus glanis Linne) muscle as influenced by different freeze-thaw cycles. Food Chem 2001, 72: 207–217. 10.1016/S0308-8146(00)00222-3
Benzie IFF, Strain JJ: The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal Biochem 1996, 239: 70–76. 10.1006/abio.1996.0292
Bernardini RD, Harnedy P, Bolton D, Kerry J, O’Neill E, Mullen AM, Hayes M: Antioxidant and antimicrobial peptidic hydrolysates from muscle protein sources and by-products. Food Chem 2011, 124: 1296–1307. 10.1016/j.foodchem.2010.07.004
Binsan W, Benjakul S, Visessanguan W, Roytrakul S, Tanaka M, Kishimura H: Antioxidative activity of Mungoong, an extract paste, from the cephalothorax of white shrimp ( Litopenaeus vannamei ). Food Chem 2008, 106: 185–193. 10.1016/j.foodchem.2007.05.065
Blois MS: Antioxidant determination by the use of stable free radicals. Nature 1958, 181: 1199–1200. 10.1038/1811199a0
Buege JA, Aust SD: Microsomal lipid peroxidation. Methods Enzymol 1978, 52: 302–304.
Chaijan M, Benjakul S, Visessanguan W, Faustman C: Changes of pigments and color in sardine ( Sardinella gibbosa ) and mackerel ( Rastrelliger kanagurta ) muscle during iced storage. Food Chem 2005, 93: 607–617. 10.1016/j.foodchem.2004.10.035
Chandrasekara A, Shahidi F: Content of insoluble bound phenolics in millets and their contribution to antioxidant capacity. J Agric Food Chem 2010, 58: 6706–6714. 10.1021/jf100868b
Chandrasekara A, Shahidi F: Inhibitory activities of soluble and bound millet seed phenolics on free radicals and reactive oxygen species. J Agric Food Chem 2011, 59: 428–436. 10.1021/jf103896z
Chantachum S, Benjakul S, Sriwirat N: Separation and quality of fish oil from precooked and non-precooked tuna heads. Food Chem 2000, 69: 289–294. 10.1016/S0308-8146(99)00266-6
Frankel EN, Huang SW, Aeschbach R: Antioxidant activity of green tea in different lipid systems. J Am Oil Chem Soc 1997, 74: 1309–1315. 10.1007/s11746-997-0062-8
Gulcin I: Antioxidant activity of eugenol: a structure-activity relationship study. J Med Food 2011, 14: 975–985. 10.1089/jmf.2010.0197
Gulcin I: Antioxidant activity of food constituents: an overview. Arch Toxicol 2012, 86: 345–391. 10.1007/s00204-011-0774-2
Guo H, Kouzuma Y, Yonekura M: Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem 2009, 113: 238–245. 10.1016/j.foodchem.2008.06.081
Hagerman AE, Riedl KM, Jones GA, Sovik KN, Ritchard NT, Hartzfeld PW, Riechel TL: High molecular weight plant polyphenolics (tannins) as biological antioxidants. J Agric Food Chem 1998, 46: 1887–1892. 10.1021/jf970975b
Hoque MS, Benjakul S, Prodpran T: Effect of heat treatment of film forming solution on the properties of film from cuttlefish ( Sepia pharaonis ) skin gelatin. J Food Eng 2010, 96: 66–73. 10.1016/j.jfoodeng.2009.06.046
Kamil JYVA, Jeon YJ, Shahidi F: Antioxidant activity of chitosan of different viscosity in cooked comminuted flesh of herring ( Clupea jarengus ). Food Chem 2002, 79: 69–77. 10.1016/S0308-8146(02)00180-2
Lei M, Wang JF, Pang L, Wang YM, Chen SG, Xue CH: Effects of Sepia on the metabolization of blood lipid and antioxidant ability in hyperlipidemia rats. Chin J Mar Drugs 2007, 3: 30–33.
Liu H, Luo P, Chen S, Shang J: Effects of squid ink on growth performance, antioxidant functions and immunity in growing broiler chickens. Asian-Aust J Anim Sci 2011, 24: 1752–1756. 10.5713/ajas.2011.11128
Liu Q, Kong B, Xiong YL, Xia X: Antioxidant activity and functional properties of porcine plasma protein hydrolysate as influenced by the degree of hydrolysis. Food Chem 2010, 118: 403–410. 10.1016/j.foodchem.2009.05.013
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ: Protein measurement with the folin phenol reagent. J Biol Chem 1951, 193: 265–275.
Lucero MT, Farrington H, Gilly WF: Qualification of L -dopa and dopamine in squid ink: implications for chemoreception. Biol Bull 1994, 187: 55–63. 10.2307/1542165
Madhavi DL, Salunkhe DK: Toxicological aspects of food antioxidants. In Food antioxidants. Edited by: Madhavi DL, Deshpande SS, Salunkhe DK. New York: Marcel Dekker; 1996:267–359.
Maqsood S, Benjakul S: Comparative studies of four different phenolic compounds on in vitro antioxidative activity and the preventive effect on lipid oxidation of fish oil emulsion and fish mince. Food Chem 2010, 119: 123–132. 10.1016/j.foodchem.2009.06.004
McConnell O, Longley RE, Koehn FE, Gullo VP: The discovery of natural products with therapeutic potential. Edited by: Gullo VP. Boston: Butterworth-Heinemann; 1994:109–174.
Mochizuki A: An antiseptic effect of cuttlefish ink. Bull Jpn Soc Sci Fish 1979, 45: 1401–1403. 10.2331/suisan.45.1401
Osman H, Suriah AR, Law EC: Fatty acid composition and cholesterol content of selected marine fish in Malaysian waters. Food Chem 2001, 73: 55–60. 10.1016/S0308-8146(00)00277-6
Park PJ, Jung WK, Nam KS, Shahidi F, Kim SK: Purification and characterization of antioxidative peptides from protein hydrolysate of lecithin-free egg yolk. J Am Oil Chem Soc 2001, 78: 651–656. 10.1007/s11746-001-0321-0
Rajaganapathy J, Thyagarajan SP, Edward JK: Study on the cephalopod’s ink for anti-retroviral activity. Indian J Exp Biol 2000, 38: 519–520.
Ranathunga S, Rajapakse N, Kim SK: Purification and characterization of antioxidative peptide derived from muscle of conger eel ( Conger myriaster ). Eur Food Res Technol 2006, 222: 310–315. 10.1007/s00217-005-0079-x
Richards MP, Hultin HO: Effect of pH on lipid oxidation using trout hemolysate as a catalyst: a possible role for deoxyhemoglobin. J Agric Food Chem 2000, 48: 3141–3147. 10.1021/jf991059w
Ramasamy MS, Murugan A: Potential antimicrobial activity of marine molluscs from Tuticorin, southeast coast of India against 40 biofilm bacteria. J Shellfish Res 2005, 24: 243–251.
Steel RGD, Torrie JH: Principle and procedure of statistics: a biometrical approach. 2nd edition. New York: McGraw-Hill; 1980.
Takai M, Kawai Y, Inoue N, Shinano H: Comparative studies on microbiological and chemical characteristics of Ika-shiokara akazukuri and Ika-shiokara kurozukuri. Bull Jpn Soc Sci Fish 1992, 58: 2373–2378. 10.2331/suisan.58.2373
Thiansilakul Y, Benjakul S, Shahidi F: Antioxidative activity of protein hydrolysate from round scad muscle using Alcalase. J Food Biochem 2007, 31: 266–287. 10.1111/j.1745-4514.2007.00111.x
Wang J, Zhao M, Zhao Q, Jiang Y: Antioxidant properties of papain hydrolysates of wheat gluten in different oxidation systems. Food Chem 2007, 101: 1658–1663. 10.1016/j.foodchem.2006.04.024
Zayas JF: Functionality of proteins in food. Berlin: Springer; 1997.
Acknowledgment
The authors would like to express their sincere thanks to Graduate School of Prince of Songkla University and the TRF Senior Research Scholar program for the financial support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
SB formulated the hypothesis and designed the studies. NKV performed experiments and analyses. NKV, SB and ABE wrote the paper. All authors have read and approved the final version of the manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Vate, N.K., Benjakul, S. Antioxidative activity of melanin-free ink from splendid squid (Loligo formosana). Int Aquat Res 5, 9 (2013). https://doi.org/10.1186/2008-6970-5-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/2008-6970-5-9