
Abstract
Background
Toxoplasma gondii is an opportunistic pathogenic protozoan parasite, which infects approximately one third of the human population worldwide, causing opportunistic zoonotic toxoplasmosis. The predilection of T. gondii for the central nervous system (CNS) causes behavioral disorders and fatal necrotizing encephalitis and thus constitutes a major threat especially to AIDS patients.
Methods
In the present study, we explored the proteomic profiles of brain tissues of the specific pathogen-free (SPF) Kunming mice at 7 d, 14 d and 21 d after infection with cysts of the Toxoplasma gondii Prugniaud (PRU) strain (Genotype II), by two-dimensional gel electrophoresis (2-DE) combined with MALDI-TOF/TOF tandem mass spectrometry (MS/MS).
Results
A total of 60 differentially expressed protein spots were selected. Fifty-six spots were successfully identified, which corresponded to 45 proteins of the mouse. Functional analysis using a Gene Ontology database showed that these proteins were mainly involved in metabolism, cell structure, signal transduction and immune responses, and will be beneficial for the understanding of molecular mechanisms of T. gondii pathogenesis.
Conclusions
This study identified some mouse brain proteins involved in the response with cyst-forming T. gondii PRU strain. These results provided an insight into the responsive relationship between T. gondii and the host brain tissues, which will shed light on our understanding of the mechanisms of pathogenesis in toxoplasmic encephalitis, and facilitate the discovery of new methods of diagnosis, prevention, control and treatment of toxoplasmic encephalopathy.
Similar content being viewed by others

Background
The obligate intracellular parasite Toxoplasma gondii is an important water- and food-borne protozoan and can infect humans as well as almost all warm-blooded animals including mammals and birds, and the infection usually persists throughout the life of the hosts[1–5]. T. gondii infects approximately 30% of the human population worldwide and 8% of population in China[6]. Humans can be infected by ingesting tissues cysts in uncooked meat or by ingesting food and water contaminated with oocysts from infected cat feces[1, 2, 6]. There are three infectious stages of T. gondii: tachyzoites (the rapidly multiplying form), bradyzoites (latent form found in tissue cysts) and sporozoites (in oocysts)[7]. Bradyzoites develop in cysts within host cells in a number of tissues, and they are more common in neural and muscular tissues.
T. gondii has long been known as an important opportunistic pathogen of immuno-compromised patients. Toxoplasmosis ranks high on the list of diseases that lead to death of the AIDS patients. Encephalitis caused by T. gondii is the most predominant manifestation of toxoplasmosis in immunosuppressed patients and is now recognized with great frequency in patients treated with immunosuppressive agents[8–11].
In the past decade, proteomic approaches have been extensively employed to study the interaction between pathogens and their hosts. The most frequently used technique in quantitative proteomics is two-dimensional electrophoresis (2-DE). In quantitative 2-DE, the appropriate experimental design plays an important role in the detection of significant and reliable protein expression differences[12]. Despite that there are some limitations of the technology, such as offering a limited dynamic range of separated proteins, it has been used to investigate host cell proteome changes after infection with T. gondii tachyzoites[13, 14], but little is currently known of the proteomic changes at differential time points in host brain tissues after infection with T. gondii cysts. In the present study, we applied 2-DE combined with mass spectrometry to study proteomic changes in mouse brain tissues infected with T. gondii cysts. The objective was to examine the proteomic modulation of host brain by cyst-forming T. gondii in vivo.
Methods
Ethics statement
The present study was approved by the Animal Ethics Committee of Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Approval No. LVRIAEC2010-008). The mice were handled in accordance with good animal practices required by the Animal Ethics Procedures and Guidelines of the People’s Republic of China.
Sample collection
Sixty Specific-Pathogen-Free (SPF)-grade female Kunming mice (including 30 mice for T. gondii infection and 30 mice as non-infected control), aged 6 to 8 weeks old, were purchased from Sun Yat-Sen University Laboratory Animal Center. All mice were handled in accordance with good animal practice according to the Animal Ethics Procedures and Guidelines of the People’s Republic of China. All mice were maintained under standard conventional conditions, with food and water ad libitum.
T. gondii (PRU) strain (Genotype II) was kindly provided by Prof Hai-Zhu Zhang (Department of Parasitology, Xinxiang Medical College, Henan, China) and were preserved in our laboratory. Tissue cysts of the PRU strain were obtained from the brains of Kunming mice infected with cysts according to Yan et al.[15]. Cysts were counted under an optical microscope. After counting, each of the 30 mice in the treatment group was inoculated intra-gastrically with 10 cysts of the PRU strain, while 30 mice in the non-infection control group were inoculated intra-gastrically with sterile physiological saline. After inoculation, the mice were observed daily for clinical symptoms. Based on a previous study[16] and our pilot study, at 7 d, 14 d and 21 d post infection, six mice from both treatment group and the control group were euthanized and the whole brain of each mouse was rapidly removed separately, washed to remove the blood, and immediately stored at −80°C until proteomic analysis, or directly processed for protein extraction. The mice that were successfully infected and produced T. gondii cysts were determined by observing tissue cysts in the brain under an optical microscope (for 14 d and 21 d).
Protein extraction
Proteins were prepared according to the previously published protocol[13]. In brief, mouse brain tissues of six mice from each group, respectively, were lysed in lysis buffer containing 7 M urea, 2% CHAPS {3-[(3-cholamidopropyl)-dimethylammonio]-1-propanesulfonate}, 2 M thiourea (Amersham), 20 mM Tris–HCl (pH 8.5, Amresco) and phenylmethylsulfonyl fluoride solution (Amresco). Then the sample was sonicated on ice (80 W, 12 s duration, 10 times, with 2 min intervals) and centrifuged at 12,000 × g for 20 min at 4°C. The supernatant was transferred to a new centrifuge tube with four times the volume of acetone added. The mixture was then precipitated overnight at −20°C and centrifuged on the next day with the same parameters. The precipitate was harvested and stored at −80°C until use. The samples were prepared in triplicates.
Isoelectric focusing electrophoresis
2-DE procedure was performed essentially according to a protocol published previously[13]. Briefly, before the isoelectric focusing electrophoresis, the precipitate was dissolved in rehydration buffer containing 7 M urea, 2% CHAPS, 2 M thiourea, followed by centrifugation at 12,000 × g for 20 min at 4°C. The protein concentration was determined by the Bradford method using a 2D Quant kit (Amresco) according to the manufacturer’s instructions. Using an equal mixture of the brain tissue protein of each mouse in the control group and treatment group respectively, proteins were initially separated using an Ettan IPGphor 3 Isoelectric Focusing system (GE Healthcare). Brain tissue proteins were focused to their isoelectric points on a 24 cm (pH 4–7) Immobiline DryStrip (GE Healthcare) with the following parameters: 300 V for 20 min, 700 V for 30 min, 1,500 V for 1.5 h, 9,000 V for 3 h and 9,000 V for 4 h. After the isoelectric focusing, the IPG strip was equilibrated for 15 min in equilibration buffer containing 2% sodium dodecyl sulfate (SDS), 50 mM Tris–HCl (pH 8.8), 6 M urea, 30% (vol/vol) glycerol, 0.002% bromophenol blue and 100 mM dithiothreitol (freshly added before use, Amresco), and followed by a second wash for 15 min with equilibration buffer containing 250 mM iodoacetamide (freshly added before use, Amersham). The IPG strip was then embedded in a precast gel and sealed into place using agarose sealing solution.
SDS-PAGE
After equilibration, the immobilized pH gradient strips were loaded onto 12.5% (w/v) homogeneous acrylamide gels (1 mm × 24 cm × 19 cm) and sealed with 1% (w/v) agarose. Proteins were separated by running the gels at 2 W/gel for 45 min and then at 18 W/gel at 10°C until the bromochlorophenol blue reached the end of gels. Finally, the gels were fixed in fixing solution (ethanol: glacial acetic acid: deionized water = 4:1:5) for 2 h, stained with Coomassie Brilliant Blue G-250 overnight and rinsed with deionized water.
Analysis of gels
Images of gels were obtained at 150 dpi (dots/in) using a scanner (Powerlook1100, UMAX) and analyzed using ImageMaster™ 2D Platinum 5.0 software (GE Healthcare). Spots were detected by 2DElite Automatic Spot Detection Program, which calculated spot volumes relative to the background and normalization. The volume percentage of each spot was determined by comparison of the spot volume to the total volume presented in the 2-DE gel. To select differentially expressed protein spots, quantitative analysis was performed using the Student’s t-test by the volume percentage of spots between brain tissues of the infected and uninfected groups on the triplicate gels. Two spots were considered significantly different if P < 0.05 and with 1.5 fold differences in volume. Spots meeting these criteria were then selected and subjected to in-gel tryptic digestion.
Protein enzymolysis
The differentially expressed protein spots were manually excised from the Coomassie Brilliant Blue-stained gels and put into a 96-well microplate. The gel pieces were washed twice with MilliQ water, destained with 50% methanol at 37°C for 30 min, and dehydrated in 100 μl of acetonitrile (ACN) at room temperature for 20 min. Next, the samples were swollen in 50 μl of 100 mM NH4HCO3, dehydrated for the second time and incubated in 1 μg/50 μl trypsin (Promega) at 4°C for 30 min. Then the samples were added with coverage solution (10% ACN, 50 mM NH4HCO3, MilliQ water) and incubated at 37°C for 16 h. After suction of the coverage solution, the peptide mixtures were extracted using 2.5% trifluoroacetic (TFA)/90% ACN at room temperature for 30 min and vacuum dried.
Identification of protein spots by MS
After vacuum drying, material was dissolved in 1.5 μl solution containing MilliQ water, 50% ACN and 0.1% TFA. Then, 0.8 μl of the mixture was loaded onto a target plate with 0.5 μl HCCA (5 mg/ml a-Cyano-4-hydroxycinnamic acid) matrix, dried at room temperature and analyzed using ABI 4800 matrix-assisted laser desorption ionization-time of flight/time of flight (MALDI-TOF/TOF) Proteomics Analyzer mass spectrometer (Applied Biosystems, USA). The UV laser was operated at a 200 Hz repetition rate with a wavelength of 355 nm and an accelerated voltage of 20 kV.
Database searching
The experimental MS data were matched to a corresponding virtual peptide mass database derived from GPS Explorer™ v 3.6, the Mascot and the International Protein Index (IPI) mouse protein database. Protein identification was carried out by peptide mass fingerprint (PMF) using the Mascot software (http://www.matrixscience.com). The search parameters used in PMF were as follows, database: IPI mouse (56871 sequences); species: mouse; enzyme: trypsin; fixed modifications: carbamidomethylation; variable modifications: oxidation (M). The function, gene name, and Gene Ontology category of each protein were determined using the Mascot v 2.1 software protein database search engine and the IPI mouse protein database.
Quantitative real-time PCR verification
Total RNA was extracted from the mouse brain tissues infected with T. gondii cysts using the Trizol reagent (Invitrogen). One microgram of total RNA was used to synthesize the first-strand cDNA, which was diluted 20 fold; 5 μl of the diluted cDNA was used as a template for real time PCR. SYBR Green-monitored real time PCR was performed on an ABI PRISM® 7500 Sequence Detection System (Applied Biosystems). The primer sequences used for real time PCR are listed in Table 1. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene for the tests.
Results
Comparative proteomic analysis by 2-DE
Global protein components of mouse brain tissues were separated with 2-DE in a 24 cm, pH 4–7 IPG strip. The 2-DE gels were processed by silver staining or Coomassie brilliant blue G-250 staining, then scanned using a UMAX scanner. Consistency of the method was confirmed by analyzing in gels in triplicate. The data were analyzed using ImageMaster™ 2D Platinum 5.0 software. The results showed that at least 2500 protein spots were detected in each gel. Spots with significant increase (or decrease) in their relative abundance were considered the differentially expressed proteins if P < 0.05 and two spots have 1.5 fold differences in volume. After bioinformatics analysis, 60 differentially expressed proteins were selected and identified using MALDI-TOF MS.
Identification of differentially expressed proteins
Proteins with MW ranging from 17 to 170 kDa and pI between 4 and 7 were separated well. Differentially expressed proteins were picked from gels and identified using MALDI-TOF MS. The data are summarized in Table 2. Sixty significantly and consistently up- or down-regulated protein spots with 1.5 fold changes of volume intensity in the triplicate gels were digested by trypsin and analyzed by MALDI-TOF MS.
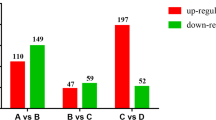
Fifty-six of 60 differentially expressed proteins were successfully identified (Figure 1), corresponding to 45 unique proteins. These proteins are listed in Table 2. Several different protein spots were identified as the same proteins by MS. Spots 4 and 18 were identified as the same protein. Spots 22 and 44 were identified as the same protein. Spots 31 and 36 were identified as different isoforms of the same protein. At 7 d post T. gondii infection, 9 spots (spots 1–9) were down-regulated and 6 were up-regulated (spots 10–15) in mouse brain tissue. At 14 d post infection, 17 spots were down-regulated (spots 16–24, 39–46) and 13 were up-regulated (spots 25–27, 47–56). At 21 d post infection, 13 protein spots (spots 28–32, 39–46) were down-regulated and 16 (spots 33–38, 47–56) were up-regulated. Eight protein spots (spots 39–46) and 10 protein spots (spots 47–56) were down- or up-regulated on both 14 d and 21 d, respectively (Figure 1).

Two-dimensional electrophoresis profiles of infected and non-infected mouse brain tissues with or without cyst-forming Toxoplasma gondii .
Analysis of differentially expressed proteins
According to the Uniprot Knowledgebase (Swiss-Prot/TrEMBL) and Gene Ontology database, the functions of successfully identified proteins were grouped based on biological processes, cellular component and molecular function. The results showed that the function of these proteins involved in cellular metabolism, structural molecule activity, immune responses, biological regulation, metabolic process, binding, catalytic activity, enzyme regulator activity, transporter activity and other functions. The differentially expressed proteins were mainly located in the cytoplasm, cell membranes and organelles (mitochondria and lysosomes), and some are secreted proteins (Figure 2).

Gene ontology (GO) categories of the identified proteins were obtained from the Uniprot Web site ( http://www.uniprot.org/ ), classified into cellular component, molecular function, and biological process according to the GO terms.
Quantitative real-time PCR verification of differentially expressed proteins
Four genes corresponding to the protein spots designated calreticulin, Rho GDP-dissociation inhibitor 1, endoplasmin and stomatin-like protein 2 were chosen for quantitative real-time PCR analysis to quantify their transcript levels. The real time PCR results were consistent with those of the 2-DE studies, and suggested that these proteins identified as differentially expressed were regulated at transcriptional level.
Discussion
The present study compared the proteomic profiles of brain tissues at 7 d, 14 d and 21 d after infection with cysts of T. gondii PRU strain by 2-DE analysis. A total of 60 differentially expressed protein spots were selected and identified by MALDI-TOF MS. Of these, 56 protein spots were successfully identified, which represented 45 different proteins. Four protein spots were not successfully identified, which may be due to the low concentrations of the proteins, which therefore failed to produce high quality mass spectrometric data.
GO analysis revealed that most of the differentially expressed proteins are involved in metabolism, cell structure, signal transduction and immune responses. Here, we focused on the discussion of the functions of several main differentially expressed proteins, and the relationships between these proteins and T. gondii infection.
Serine protease inhibitor (SERPIN) A3k is persistently up-regulated in mouse brain tissues 7, 14 and 21 days after infection with Toxoplasma cysts. Many members of the serine protease inhibitor superfamily play an important role in the physiological and pathological process, and can be regarded as protease inhibitors which are involved in the coagulation reaction, fiber dissolution, angiogenesis, complement activation, immune and inflammatory reaction[17, 18]. It was suggested that SERPINs could inhibit host cell apoptosis[19]. In addition, SERPIN may inhibit replication and decrease T. gondii viability[20]. Therefore, the persistence of up-regulated SERPIN A3k may play an important role in preventing the death of infected mouse brain cells as well as limiting the growth of T. gondii in the brain tissue cysts.
Protein disulfide isomerase (PDI) is down-regulated 14 days after infection with T. gondii cysts. PDI family members can function as molecular chaperones and as disulfide oxidoreductase/isomerases, which means that they can make, break, or rearrange disulfide bonds[21]. The PDI family’s main function is to catalyze the oxidative folding of nascent polypeptide chains in the endoplasmic reticulum, yet they also play an important role in the ER-associated protein degradation pathway (ERAD), protein transport, calcium homeostasis, antigen presentation and virus invasion[22, 23]. PDI may play an important role in immune and inflammatory responses and one of the intracellular effector molecules involved in anti-inflammatory reactions[24].
Endoplasmin, also known as heat shock protein 90B1, is a molecular chaperone protein. In the present study, it showed sustained up-regulation in brain tissues 14 and 21 days after infection with T. gondii. In Toll like receptor and integrin secretory pathways, it plays a crucial role in folding protein, so was thought to be one of the basic immune chaperone proteins in the regulation of innate and adaptive immunity[25]. Calreticulin (CRT) is persistently up-regulated in brain tissues 14 and 21 days after infection with Toxoplasma cysts. Calreticulin is a Ca2+ binding protein that has been implicated in many diverse functions inside and outside of the endoplasmic reticulum (ER), including the regulation of intracellular Ca2+ homeostasis and Ca2+-dependent pathways, chaperone activity, steroid-mediated gene expression, cell adhesion and the interactions of CRT with immunoglobulin G and immunoglobulin Y[26–28]. CRT could be used as a biomarker in lung cancer prediction and diagnosis[29]. CRT also plays an important role in autoimmunity. Enolase 1 and CRT are important proteins in regulating the differentiation and functions of mouse mast cells[30]. CRT has important implications involved in the genesis, development and prognosis of many diseases.
Lamin B1 is down-regulated in mouse brain tissues 7 days after infection with T. gondii cysts. Lamins are the important cytoskeletal proteins in the nucleus, and can be divided into two types A and B. Lamin B1 is necessary for growth, development and nuclear membrane integrity in mice[31–33]. Cytochrome b5 is also down-regulated in mouse brain tissues 7 days after infection with T. gondii cysts. It is a small microsomal protein which serves as an electron transfer component in a number of oxidative reactions in biological tissues, including the anabolic metabolism of fats and steroids, as well as the catabolism of xenobiotics and compounds of endogenous metabolism[34].
Prohibitin (PHB) is down-regulated in mouse brain tissues 14 days after infection with T. gondii cysts. PHB has a variety of cell biological functions, including the regulation of cell proliferation, apoptosis, development, transcription, mitochondrial protein folding and as a cell surface receptor[35–37]. In the present study, α-tubulin of mouse brain tissues is up-regulated after infection with T. gondii cysts at 21 days post infection, but β-tubulin is down-regulated at 14 days post infection. α- and β-tubulins are the major components of microtubules of the eukaryotic cytoskeleton. Microtubules constitute a major portion of cytoplasmic proteins in nerve cells. They have been implicated to play a central role in axonal transport, neurotransmitter release, neurite outgrowth and synaptogenesis. An increase in the tubulin microheterogeneity was demonstrated during brain maturation. Tubulin comprises a large percentage of the total protein in brain[38–40]. Therefore, abnormal expression of tubulin could have an effect on the normal growth and development of host brain, even causing behavioral disorders of the host.
Apolipoprotein E (ApoE) is persistently up-regulated in brain tissues at 14 and 21 days after infection with Toxoplasma cysts. ApoE is one of components of plasma lipoprotein. It regulates the metabolism of plasma lipoprotein by binding to lipoprotein receptors[41]. Abnormal expression of the protein causes the disorder of the metabolism of plasma lipoprotein, and is closely related to atherosclerosis, hypercholesterolemia and hypertriglyceridemia[42].
Conclusions
The present study revealed changes in the proteomic profiles of mouse brain tissues after infection with cyst-forming T. gondii PRU strain (Genotype II). 45 mouse brain proteins differentially expressed between infected and non-infected mice were identified, these proteins were mainly involved in metabolism, cell structure, signal transduction and immune responses. Further exploration of these proteomic data will contribute to understanding the pathogenesis of toxoplasmic encephalitis, and facilitates the discovery of new methods of diagnosis, prevention, control and treatment of Toxoplasma encephalopathy.
References
Dubey JP: Toxoplasmosis of Animals and Humans. 2010, Boca Raton, Florida: CRC Press, 313-second
Dubey JP, Jones JL: Toxoplasma gondii infection in humans and animals in the United States. Int J Parasitol. 2008, 38: 1257-1278. 10.1016/j.ijpara.2008.03.007.
Nardoni S, Angelici MC, Mugnaini L, Mancianti F: Prevalence of Toxoplasma gondii infection in Myocastor coypus in a protected Italian wetland. Parasit Vectors. 2011, 4: 240-10.1186/1756-3305-4-240.
Chen J, Xu MJ, Zhou DH, Song HQ, Wang CR, Zhu XQ: Canine and feline parasitic zoonoses in China. Parasit Vectors. 2012, 5: 152-10.1186/1756-3305-5-152.
Tian YM, Dai FY, Huang SY, Deng ZH, Duan G, Zhou DH, Yang JF, Weng YB, Zhu XQ, Zou FC: First report of Toxoplasma gondii seroprevalence in peafowls in Yunnan Province, Southwestern China. Parasit Vectors. 2012, 5: 205-10.1186/1756-3305-5-205.
Zhou P, Chen Z, Li HL, Zheng H, He S, Lin RQ, Zhu XQ: Toxoplasma gondii infection in humans in China. Parasit Vectors. 2011, 4: 165-10.1186/1756-3305-4-165.
Dubey JP: History of the discovery of the life cycle of Toxoplasma gondii. Int J Parasitol. 2009, 39: 877-882. 10.1016/j.ijpara.2009.01.005.
da Silva RC, Langoni H: Toxoplasma gondii: host-parasite interaction and behavior manipulation. Parasitol Res. 2009, 105: 893-898. 10.1007/s00436-009-1526-6.
Suzuki Y: Host resistance in the brain against Toxoplasma gondii. J Infect Dis. 2002, 185 (Suppl 1): S58-65.
Afonso C, Paixão VB, Costa RM: Chronic Toxoplasma infection modifies the structure and the risk of host behavior. PLoS One. 2012, 7: e32489-10.1371/journal.pone.0032489.
Carruthers VB, Suzuki Y: Effects of Toxoplasma gondii infection on the brain. Schizophr Bull. 2007, 33: 745-751. 10.1093/schbul/sbm008.
Tchankouo-Nguetcheu S, Bourguet E, Lenormand P, Rousselle JC, Namane A, Choumet V: Infection by chikungunya virus modulates the expression of several proteins in Aedes aegypti salivary glands. Parasit Vectors. 2012, 5: 264-10.1186/1756-3305-5-264.
Zhou DH, Yuan ZG, Zhao FR, Li HL, Zhou Y, Lin RQ, Zou FC, Song HQ, Xu MJ, Zhu XQ: Modulation of mouse macrophage proteome induced by Toxoplasma gondii tachyzoites in vivo. Parasitol Res. 2011, 109: 1637-1646. 10.1007/s00436-011-2435-z.
Nelson MM, Jones AR, Carmen JC, Sinai AP, Burchmore R, Wastling JM: Modulation of the host cell proteome by the intracellular apicomplexan parasite Toxoplasma gondii. Infect Immun. 2008, 76: 828-844. 10.1128/IAI.01115-07.
Yan HK, Yuan ZG, Song HQ, Petersen E, Zhou Y, Ren D, Zhou DH, Li HX, Lin RQ, Yang GL, Zhu XQ: Vaccination with a DNA vaccine coding for perforin-like protein 1 and MIC6 induces significant protective immunity against Toxoplasma gondii. Clin Vaccine Immunol. 2012, 19: 684-689. 10.1128/CVI.05578-11.
Djurković-Djaković O, Djokić V, Vujanić M, Zivković T, Bobić B, Nikolić A, Slavić K, Klun I, Ivović V: Kinetics of parasite burdens in blood and tissues during murine toxoplasmosis. Exp Parasitol. 2012, 131: 372-376. 10.1016/j.exppara.2012.05.006.
Silverman GA, Bird PI, Carrell RW, Church FC, Coughlin PB, Gettins PG, Irving JA, Lomas DA, Luke CJ, Moyer RW, Pemberton PA, Remold-O’Donnell E, Salvesen GS, Travis J, Whisstock JC: The serpins are an expanding superfamily of structurally similar but functionally diverse proteins. Evolution, mechanism of inhibition, novel functions, and a revised nomenclature. J Biol Chem. 2001, 276: 33293-33296. 10.1074/jbc.R100016200.
Molehin AJ, Gobert GN, McManus DP: Serine protease inhibitors of parasitic helminths. Parasitology. 2012, 139: 681-695. 10.1017/S0031182011002435.
Ahn HJ, Kim JY, Ryu K, Nam HW: STAT6 activation by Toxoplasma gondii infection induces the expression of Th2 C-C chemokine ligands and B clade serine protease inhibitors in macrophage. Parasitol Res. 2009, 105: 1445-1453. 10.1007/s00436-009-1577-8.
Shaw MK, Roos DS, Tilney LG: Cysteine and serine protease inhibitors block intracellular development and disrupt the secretory pathway of Toxoplasma gondii. Microb Infect. 2002, 4: 119-132. 10.1016/S1286-4579(01)01520-9.
Benham AM: The protein disulfide isomerase family: key players in health and disease. Antioxid Redox Signal. 2012, 16: 781-789. 10.1089/ars.2011.4439.
Wilkinson B, Gilbert HF: Protein disulfide isomerase. Biochim Biophys Acta. 2004, 1699: 35-44.
Stolf BS, Smyrnias I, Lopes LR, Vendramin A, Goto H, Laurindo FR, Shah AM, Santos CX: Protein disulfide isomerase and host-pathogen interaction. Sci World J. 2011, 11: 1749-1761.
Higuchi T, Watanabe Y, Waga I: Protein disulfide isomerase suppresses the transcriptional activity of NF-kappaB. Biochem Biophys Res Commun. 2004, 318: 46-52. 10.1016/j.bbrc.2004.04.002.
Morales C, Wu S, Yang Y, Hao B, Li Z: Drosophila glycoprotein 93 is an ortholog of mammalian heat shock protein gp96 (grp94, HSP90b1, HSP4) and retains disulfide bond-independent chaperone function for TLRs and integrins. J Immunol. 2009, 183: 5121-5128. 10.4049/jimmunol.0900811.
Wang WA, Groenendyk J, Michalak M: Calreticulin signaling in health and disease. Int J Biochem Cell Biol. 2012, 44: 842-846. 10.1016/j.biocel.2012.02.009.
Gelebart P, Opas M, Michalak M: Calreticulin, a Ca2 + −binding chaperone of the endoplasmic reticulum. Int J Biochem Cell Biol. 2005, 37: 260-266. 10.1016/j.biocel.2004.02.030.
Møllegaard KM, Duus K, Træholt SD, Thaysen-Andersen M, Liu Y, Palma AS, Feizi T, Hansen PR, Højrup P, Houen G: The interactions of calreticulin with immunoglobulin G and immunoglobulin Y. Biochim Biophys Acta. 1814, 2011: 889-899.
Liu R, Gong J, Chen J, Li Q, Song C, Zhang J, Li Y, Liu Z, Dong Y, Chen L, Jin B: Calreticulin as a potential diagnostic biomarker for lung cancer. Cancer Immunol Immunother. 2012, 61: 855-864. 10.1007/s00262-011-1146-8.
Ryu SY, Hong GU, Kim DY, Ro JY: Enolase 1 and calreticulin regulate the differentiation and function of mouse mast cells. Cell Signal. 2012, 24: 60-70. 10.1016/j.cellsig.2011.07.011.
Shimi T, Butin-Israeli V, Adam SA, Hamanaka RB, Goldman AE, Lucas CA, Shumaker DK, Kosak ST, Chandel NS, Goldman RD: The role of nuclear lamin B1 in cell proliferation and senescence. Genes Dev. 2011, 25: 2579-2593. 10.1101/gad.179515.111.
Vergnes L, Péterfy M, Bergo MO, Young SG, Reue K: Lamin B1 is required for mouse development and nuclear integrity. Proc Natl Acad Sci USA. 2004, 101: 10428-10433. 10.1073/pnas.0401424101.
Martin C, Chen S, Maya-Mendoza A, Lovric J, Sims PF, Jackson DA: Lamin B1 maintains the functional plasticity of nucleoli. J Cell Sci. 2009, 122: 1551-1562. 10.1242/jcs.046284.
Schenkman JB, Jansson I: The many roles of cytochrome b5. Pharmacol Ther. 2003, 97: 139-152. 10.1016/S0163-7258(02)00327-3.
Van Aken O, Whelan J, Van Breusegem F: Prohibitins: mitochondrial partners in development and stress response. Trends Plant Sci. 2010, 15: 275-282. 10.1016/j.tplants.2010.02.002.
Theiss AL, Sitaraman SV: The role and therapeutic potential of prohibitin in disease. Biochim Biophys Acta. 1813, 2011: 1137-1143.
Mishra S, Ande SR, Nyomba BL: The role of prohibitin in cell signaling. FEBS J. 2010, 277: 3937-3946. 10.1111/j.1742-4658.2010.07809.x.
George HJ, Misra L, Field DJ, Lee JC: Polymorphism of brain tubulin. Biochemistry. 1981, 20: 2402-2409. 10.1021/bi00512a006.
Eipper BA: Properties of rat brain tubulin. J Biol Chem. 1974, 249: 1407-1416.
Gozes I, Littauer UZ: Tubulin microheterogeneity increases with rat brain maturation. Nature. 1978, 276: 411-413. 10.1038/276411a0.
Hauser PS, Narayanaswami V, Ryan RO: Apolipoprotein E: from lipid transport to neurobiology. Prog Lipid Res. 2011, 50: 62-74. 10.1016/j.plipres.2010.09.001.
Gregg RE, Zech LA, Schaefer EJ, Stark D, Wilson D, Brewer HB: Abnormal in vivo metabolism of apolipoprotein E4 in humans. J Clin Invest. 1986, 78: 815-821. 10.1172/JCI112645.
Acknowledgments
Project support was provided by the National Natural Science Foundation of China (Grant Nos. 31230073, 31172316 and 31228022). We thank Guangzhou FitGene Biotechnology Co., Ltd., China, for technical assistance.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
XQZ and DHZ conceived and designed the study, and critically revised the manuscript. DHZ, FRZ and SYH performed the experiments, analyzed the data and drafted the manuscript. MJX, HQS and CS helped in study design, study implementation and manuscript revision. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Zhou, DH., Zhao, FR., Huang, SY. et al. Changes in the proteomic profiles of mouse brain after infection with cyst-forming Toxoplasma gondii. Parasites Vectors 6, 96 (2013). https://doi.org/10.1186/1756-3305-6-96
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1756-3305-6-96