
Abstract
Canine babesiosis is a clinically significant emerging vector-borne disease caused by protozoan haemoparasites. This review article considers recent literature pertaining to the taxonomic classification of Babesia and Theileria species affecting dogs and the geographical distribution of these parasites. The diagnosis of canine babesiosis by traditional, molecular and serological methods is reviewed, together with recent advances in our understanding of the pathophysiology of piroplasmosis, and of the treatment and prevention of this disease.
Similar content being viewed by others

Introduction
Canine babesiosis is a common and clinically significant tick-borne haemoprotozoan disease with a worldwide distribution. Babesia species are often referred to as piroplasms, a collective term for phenotypically similar protozoan parasites that utilise mammalian erythrocytes in their life cycle. Piroplasms of domestic animals encompass two main genera, Babesia and Theileria, and have been subject of intense research interest and molecular-based re-classification during the last 10 years. In dogs, infection by these haemoparasites results in a wide range of clinical presentations; from subclinical disease to serious illness characterised by fever, pallor, jaundice, splenomegaly, weakness and collapse associated with intra- and extravascular haemolysis, hypoxic injury, systemic inflammation, thrombocytopenia and pigmenturia [1–3]. Although canine babesiosis is recognised as a tick-borne disease, transmitted by a variety of well-described Ixodid vectors around the world, Babesia gibsoni is an emerging disease with molecular evidence of clonal expansion due to non-vectored transmission by blood exchange during fighting and biting [4–6]. Recent research into canine babesiosis has focussed on determining the taxonomic status of well recognised and newly discovered canine piroplasms, developing improved diagnostic methods, investigating aspects of pathophysiology and searching for improved chemotherapeutic and immunoprophylactic protocols.
Taxonomy and molecular phylogeny
The classification of Babesia spp. places them in order Piroplasmida within the phylum Apicomplexa. Two morphologically distinct forms of the erythrocytic stage in the canine host were recognised in early studies that led to the naming of the larger form, measuring approximately 3–5 μm as B. canis, and the smaller (1–3 μm) as B. gibsoni. Despite painstaking observation of the parasites in blood films by many early researchers, further understanding of the taxonomic classification of these parasites was hampered for the best part of one hundred years by the fact that within these general size ranges, their morphological features did not permit further differentiation. The first suggestion that all B. canis isolates were not identical species came from the German protozoologist Eduard Reichenow who recognised differences in pathogenicity of "B. canis" isolates from France and North Africa (these were most likely parasites that are currently known as B. (canis) canis and B. (canis) vogeli, respectively) [7, 8]. Further clarification did not come until the late 1980s with the advent of molecular tools for phylogenetic studies; molecular genotyping of canine piroplasms has resulted currently in the identification of four large and at least four small parasites, but it is likely that new species will be added as further isolates are characterised.
"Babesia canis" was reclassified into three sub-species (B. canis canis, B. canis rossi and B. canis vogeli) on the basis of cross-immunity, serological testing, vector specificity and molecular phylogeny; these parasites are now considered to be separate species in their own right [9, 10] (Table 1). A fourth 'large' (as yet unnamed) Babesia sp. has been described recently in a number of dogs with clinical signs and haematological parameters consistent with babesiosis in North Carolina [11, 12]. With regard to small piroplasms, three genetically and clinically distinct species are currently recognised to cause disease in dogs; Babesia gibsoni, Babesia conradae (reported in dogs in the western United States and described in original reports as "B. gibsoni") [13, 14], and a Babesia microti-like piroplasm (named Theileria annae) [15, 16].
In addition, three Theileria species have been isolated in a small number of dogs' blood in Europe (Theileria (Babesia) equi and Theileria annulata) [17, 18] and from 82 dogs in South Africa (unnamed Theileria sp. related to an isolate obtained from antelope) [19] (Table 1). Until more information is available the competence of the dog as a host for these piroplasms is uncertain; the clinical correlation for these organisms is unknown, and neither intraerythrocytic nor extraerythrocytic stages have yet been visualised.
As with many taxonomic debates, the classification of canine piroplasms has not been without disagreement among parasitologists, especially concerning nomenclature of the smaller parasites. Furthermore, data from molecular analysis of the mammalian host stages have far outstripped our understanding of the life-cycle features of these organisms, notably identification of their vectors and other important biological data. Under light microscopy the intraerythrocytic stage of Babesia spp. is indistinguishable from Theileria spp. (In cats Cytauxzoon is also similar in appearance, but to date there has been no report of this genus affecting canines.) Historically these two genera, Babesia and Theileria, are separated on the basis of certain life-cycle stages and transovarial passage within the tick vector [20], yet to date the only piroplasm of companion animals known to have an extra-erythrocytic (schizogony) stage to its life cycle is Cytauxzoon felis in the cat.
The controversy focuses on whether all small canine piroplasm species should be classified as Babesia spp. or whether Theileria spp. infect dogs as part of their natural life cycle. To inform the debate, molecular studies of canine piroplasms have predominantly utilised the small subunit ribosomal 18S gene locus to infer phylogenetic relationships, favoured for its conserved nature and predictable rate of mutation, but other rRNA loci, cytochrome b, and genes encoding the heat-shock proteins have been also utilised for this purpose [6, 18, 21].
As noted above, pre-erythrocytic parasite (schizont) stages in lymphocytes or macrophages, a defining feature of Theileria, have not been observed in dogs. However, the formation of distinct tetrad forms (the "Maltese cross"), considered as a feature of both Theileria spp. and Babesia microti, has been described in Babesia conradae [14, 22]. Babesia conradae appears to be most closely related to a group of piroplasms found in free-ranging ruminants (deer and sheep) and isolated from humans in the western United States – the 'western clade' [22]. This group is phylogenetically distinct from Babesia microti, a rodent piroplasm with a wide distribution throughout the Holarctic ecozone (see review [23]) in which schizogony in lymphocytes has been reported [24]. The piroplasm species reported in dogs from northern Spain forms into a clade with Babesia microti and was given the name Theileria annae [15], but is often referred to as the "Babesia microti-like" piroplasm or agent, or "Spanish isolate". Indisputably Theileria species (Theileria annulata and Theileria equi) have been detected in canine blood [17, 18] and the most recently reported canine piroplasm, in South Africa [19], is also phylogenetically aligned with recognised Theileria spp., yet as stated previously their significance is unclear at the present time.
Geographical distribution
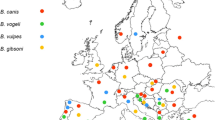
The distribution maps for canine vector-borne diseases are continually changing as new information about parasite infections and ranges comes to light. In general it may be useful to consider two levels of regional prevalence and incidence for specific diseases; those regions where the specific parasite is well established (i.e., endemic) and clinically recognised; and those regions where sporadic autochthonous infections or cases associated with travelling dogs have been reported.
Established endemic ranges
The established ranges for canine piroplasms are listed in Table 1; comprehensive and up-to-date canine vector-borne disease maps of Europe, and East and Southeast Asia are available [25]. As would be expected, the geographical distribution of these organisms is delineated largely by the ecological ranges of their vector ticks. Interestingly this generalisation seems not to apply to Babesia gibsoni, whose rapid recent global dispersion is now proposed to have been achieved predominantly by direct dog-to-dog transmission under unique ownership and management practices, without involvement of a vector (see later discussion) [4, 5].
Babesia vogeli is the most widespread canine piroplasm due to the cosmopolitan nature of its host, the brown dog tick Rhipicephalus sanguineus (Table 1). Babesia vogeli has a truly worldwide distribution throughout tropical and subtropical regions and extending into cooler latitudes where it potentially occurs alongside (and may be confused with) the other large Babesia spp. Babesia canis (sensu strictu) is transmitted by Dermacentor spp. and has been increasingly recognised throughout central Europe when it was once thought to be confined to France. Babesia rossi is the other African Babesia (with B. vogeli and B. gibsoni); originally recognised only in South Africa but has been reported recently in other regions of the African continent including Nigeria [26] and Sudan [27] where its vector ticks (Haemaphysalis spp.) are enzootic (Table 1).
The small canine piroplasm with the greatest geographical distribution is undoubtedly B. gibsoni. The type-species is referred to as the "Asia strain", reflecting its original molecular identification from a number of different southern, eastern and southeastern Asian countries. However, in the last 10 years B. gibsoni infections have been reported in many countries outside Asia, in American Pit Bull Terrier-type dogs predominantly, and there is now convincing evidence that these cases have arisen due to biting and fighting between infected and non-infected dogs [4, 28]. Gene sequence conservation has been noted in the ITS and 18S gene of B. gibsoni isolates originating in many disparate regions of the world [6, 22, 29]. This observation has led to the intriguing hypothesis that the lack of genetic diversity in B. gibsoni is a result of the absence of sexual reproduction (sporogony) in the tick; in other words a clonal expansion of a single strain is occurring within a susceptible host population. Therefore the dogs themselves are the reservoir for B. gibsoni, specific fighting dogs in particular, and not an endemic species of tick. Due to the worldwide popularity of this and similar breeds, it is speculated that Babesia gibsoni will be reported eventually from all countries where (usually illegal) dog fighting is practiced.
The three most recently characterised small piroplasm species of dogs occur, as previously mentioned, in the western United States, specifically California (Babesia conradae), southern Africa (Theileria sp.) and the Iberian peninsular (Theileria annae), where the latter is thought to be transmitted by Ixodes hexagonus. The vectors for Babesia conradae and the South African Theileria sp. are currently unknown and apart from a single case report of Theileria annae in the USA [28], these piroplasms have not been reported outside these fairly localised geographical boundaries.
Autochthonous infections and other single infection reports
Autochthonous infection refers to discovery of an infection that has originated in the place where it is found, usually in the context of a new, or unexpected finding in a location where the infection is generally not considered to occur. In the case of babesiosis, autochthonous cases are most likely to arise when dogs have come into contact with infected vectors which may themselves have been introduced by hosts returning from vector-endemic areas, or by extension of the vector range due to ecological changes. There have been a number of reports of autochthonous babesiosis and piroplasmosis in northern Europe (where vector-borne diseases have been considered unusual or exotic) and such cases have raised concern among the local veterinary and pet-owning fraternities. The importance of collecting travel history as an integral step during a veterinary consultation has been emphasised, together with concern about the risks of disease spread associated with deregulated pet travel, particularly in mainland Europe and the UK. The history of being a hunting dog or extensive rural exposure is a risk factor for canine babesiosis in Europe [30].
Diagnosis of babesiosis
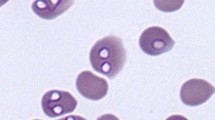
Microscopy remains the simplest and most accessible diagnostic test for most veterinarians and during acute infections microscopy is reasonably sensitive for detecting intraerythrocytic parasites in Giemsa or Wright's stained blood smears. Differentiation between large and small piroplasms is also relatively simple. Moreover, microscopy is still the only viable option available to veterinarians in many parts of the developing world where babesiosis is endemic. With large Babesia species at least, sampling from capillary beds (ear tip, toe nail) or examination of cells from beneath the buffy coat of a haematocrit tube may improve the probability of finding parasites [31, 32].
The diagnosis of piroplasmosis in chronically infected and carrier dogs however remains a significant challenge due to very low, often intermittent parasitaemias. Failure to detect Babesia/Theileria parasites in animals with hemolytic anaemia or thrombocytopenia has led to an incorrect diagnosis in documented cases, often when the clinical suspicion of babesiosis was also low. Given the possibility of direct horizontal transmission of Babesia gibsoni, veterinary clinicians should always ascertain whether the patient has been bitten by any other dog in the preceding 4–8 weeks, irrespective of its breed [4, 28].
Molecular diagnosis – conventional PCR and other methods
Although PCR has greatly increased the sensitivity and specificity of parasite detection and is well suited to epidemiological and phylogenetic studies, access to molecular techniques for routine clinical diagnosis of babesiosis is still restricted to relatively few laboratories worldwide. Ribosomal RNA genes 18S, 5.8S, 28S and the internal transcribed spacer (ITS) sequences have been used for conventional PCR, but some researchers have chosen other loci such as the p18/BgTRAP [33]. Since parasite morphology is a poor guide to speciation, modifications of the PCR have been utilised to rapidly differentiate between piroplasm species; PCR-RFLP and nested PCR have been reported to differentiate B. vogeli and B. gibsoni in Australia [34], and between the large babesial species [9] and B. gibsoni in endemic regions [35]. In a clinicopathological study of large Babesia spp. infections in dogs in Italy using PCR-RFLP, Babesia canis was detected in 34/164 and Babesia vogeli in 11/164 dogs. Although a distinct geographical difference in the incidence of these infections is noted [30], many of the dogs with B. canis infection had recently returned from hunting trips in Eastern Europe. Further refinement in primer design was reported recently to clearly separate amplicons of 342 bp, 546 bp, and 746 bp target fragments of B. canis rossi, B. canis vogeli, B. canis canis, respectively [36]. Loop-mediated isothermal amplification (LAMP) was found to have advantages of speed and specificity for detecting B. gibsoni infections in Japan [37] and reverse-line blot (RLB) hybridisation was applied in epidemiological studies of arthropod-borne haemopathogens of dogs and cats in Trinidad [38] and dogs in Africa [19]. Real-time PCR enables quantification of pathogen levels within blood and tissue samples; the amount of final PCR product can be used to deduce the starting number of target molecules and infer parasite levels within a host. Quantitative PCR (qPCR) was used in an experimental infection of 3 dogs with B. gibsoni that concluded that the methodology might be adapted to the determination of vaccine or chemotherapeutic efficacy, or the elucidation of immunological responses [39]. Furthermore, a number of these PCR methods have been applied to filter-paper technologies such as FTA cards (Whatman Bioscience) and IsoCode Stix (Schleicher and Scheull) for ease of transport of samples to distant laboratories and for epidemiological and other diagnostic studies [34, 40]. Long-term storage and archival qualities of this methodology have not been investigated [40].
Whereas the detection limit of light microscopy is approximately 0.001% parasitaemia, PCR is capable of detecting parasite loads in the region of 50 organisms/ml [35] and 9 parasites/μl [39]. Yet despite its extraordinary sensitivity, PCR will clearly not detect target DNA when there are no organisms within the sample. "False negative" results may occur in chronic babesiosis and it is very important to recognise this limitation when screening potential carriers and other asymptomatic dogs such as blood donors. The ability of PCR to detect infected dogs in such situations, with and without treatment, has been investigated [41, 42]. In one study [42] clinical parameters, haematology, serologic titer (by IFAT) and the presence of Babesia DNA was monitored on a daily basis after experimental infection. All dogs (n = 3) made a full clinical recovery, as judged by normal clinical signs, absence of splenic enlargement, a normal haemogram and absence of piroplasms on microscopic examination by 30–50 days after peak parasitaemia. During this period of clinical normality, babesial DNA was inconsistently detected. This suggests a very low, fluctuating parasitaemia in these dogs, possibly analogous to chronic, asymptomatic natural infection.
The ability of PCR to detect parasite DNA in chronically infected animals can be improved by testing on more than one occasion, but the use of serology as an alternative, complementary diagnostic test is advisable in these situations [42, 43].
Serological testing – the IFAT and ELISA
Immunofluorescent antibody testing (IFAT) has been the most widely supported serological diagnostic test for canine babesiosis for the last 30 years [44, 45]. However, poor specificity due to cross-reactions between Babesia spp. and with other apicomplexan parasites, operator subjectivity and its inadequacy for large-scale screening have all been limiting factors [46]. Recent research by several groups in Japan directed towards finding specific immunodominant Babesia gibsoni antigens for use in recombinant protein enzyme-linked immunosorbent assays (ELISA) – and potential vaccine candidates in the future – has identified numerous promising compounds [46–52]. Thrombospondin-related adhesive proteins (TRAPs) comprise a group of highly conserved functional proteins identified in apicomplexan parasites, mooted to be associated with merozoite motility and invasion, and capable of inducing a host antibody response [49]. An ELISA using recombinant BgTRAP was reported to be more sensitive than other ELISAs using recombinant antigens rBgP50, rBgSA1, and rBgP32 [43], and Konishi et al. (2008) utilised a BgTRAP ELISA to test 1,206 randomly selected non-fighting breed dogs in Japan to test for exposure to B. gibsoni [53]. These authors reported higher infection rates in western Japan and concluded that having excluded fighting dogs, a history of tick (Haemaphysalis longicornis) exposure was a significant risk factor for anti-babesial antibody detection, thus demonstrating two distinct epidemiological patterns of B. gibsoni infection in that country.
A second major limitation of serological tests is their inability to differentiate acute from chronic infections, and interpretation of a positive titre is somewhat problematic for clinicians working in regions that are endemic for babesiosis. Nevertheless, for B. gibsoni in the USA and Australia, where cases are sporadic, the IFAT is a useful tool for detection of infected dogs, especially if combined with PCR [31, 38].
Pathophysiology
The severity of babesiosis in dogs and cats ranges from subclinical infection, the development of mild anaemia to widespread organ failure and death. The critical determinant of this variable pathogenesis is the piroplasm species, yet other factors such as the age and immune status of the host and concurrent infections or illness also play a role. All species (Table 1) may cause pyrexia, anorexia, splenomegaly, anaemia and thrombocytopenia. Direct parasite-induced red-cell damage, increased osmotic fragility of infected cells, oxidative and secondary immune-mediated injury of the erythrocyte membrane result in a combination of intravascular and extravascular haemolysis.
The clinical features of babesiosis have been reviewed elsewhere [1–3]. In broad terms it is generally agreed that the least pathogenic of the well-recognised canine piroplasm species is Babesia vogeli, at least in adult dogs, and the most virulent is Babesia rossi in Africa [1, 30, 31]. With Babesia rossi infections a large proportion of dogs develop complications, some of which (hepatopathy, immune-mediated haemolysis) typically extend hospital stay but do not affect mortality if treated appropriately, while others (haemoconcentration, neurological signs, acute renal failure and pulmonary oedema) require early, aggressive and intensive therapy and carry a poor prognosis [32]. In contrast, Babesia vogeli is often subclinical (except in puppies less than 3–4 months old, in which infection may be fatal); it is occasionally observed in blood films of dogs with other primary disease or receiving medical treatment (e.g., immunosuppression or chemotherapy) or surgery (notably splenectomy), and in these potentially immunocompromised individuals the appearance of the parasite may be inconsequential to the outcome (and may not even warrant treatment).
The pathogenicity of Babesia canis, Babesia gibsoni, Theileria annae and Babesia conradae is moderate to severe in dogs, but it should be stressed again that a wide range of clinical signs of varying severity can be observed in individuals. Babesia conradae is considered to be more pathogenic than B. gibsoni, resulting in higher parasitaemias and more severe anaemia [14]. In Spain, Theileria annae infection is associated with severe haemolysis and azotaemia [16]. The unnamed Babesia sp. from North Carolina has been associated with non-specific illness (lethargy and anorexia), pigmenturia and mild fever, predominantly in splenectomised dogs [54]. Laboratory findings have included mild anaemia and severe thrombocytopenia, similar to Babesia vogeli infections [54].
The potential for renal involvement in dogs with babesiosis has received attention in recent years, with hypoxaemia, haemoglobinuric nephropathy and glomerulonephritis all considered possible mechanisms and supported by histological studies [55]. A clinicopathological study of Theileria annae infection in Northwest Spain reported 36% dogs (n = 58) were azotaemic at the time of diagnosis and that these dogs had a 10-fold higher risk of death from piroplasmosis than those that were not azotaemic [56]. Unfortunately neither urine-specific gravity nor urine osmolality was reported in these dogs, but elevated urine protein: creatinine ratios, hypoalbuminaemia and hypercholesterolaemia in affected dogs led the authors to suggest that glomerular injury was occurring and that renal failure was most likely a feature of Theileria annae infection [56]. In contrast, study of azotaemia associated with B. rossi infection concluded that urea and creatinine were unreliable indicators of renal damage or acute renal failure in acute babesiosis [57, 58]. Mortality of B. rossi was significantly associated with high cortisol and high ACTH concentrations and with low T4 and fT4 concentrations in a recent study investigating endocrine markers of disease [59].
The clinical consequences of chronic babesial infection are unclear and while most dogs appear to tolerate this state of premunity with few ill effects, theoretically they remain at risk of developing immune-mediated complications and recrudescence of clinical disease (and parasitaemia) if immunocompromised at a later time. Chronic infection may be inconsequential in some dogs and may be even beneficial for hosts living in endemic regions by protecting them from further disease [60].
Recent advances in the treatment and prevention of babesiosis
Despite a plethora of anecdotal reports and uncontrolled experimental data, there is a paucity of scientifically robust evidence regarding the efficacy of drugs that have been used to treat canine babesiosis over the last 100 years. Early treatment studies were hampered by the need to rely on clinical signs and blood films to determine parasite clearance. More recently the limitations have been the cost of establishing controlled infections and the relatively small numbers of experimental dogs permitted by animal ethics committees. In order to determinate the efficacy of an anti-piroplasm drug with reasonable confidence, necropsy-obtained tissues from all organs need to be tested by validated PCR at the end of the experiment; clearance of the parasite DNA from peripheral blood alone is insufficient evidence of cure.
Imidocarb dipropionate and diminazine aceturate are widely used anti-piroplasm drugs, but other compounds that have been used for many years and have enjoyed varying degrees of success in managing the clinical signs of piroplasmosis include quinuronium sulphate, trypan blue, pentamidine, phenamidine and parvaquone. National registration authorities have restricted access to some of these drugs in certain countries, and some, notably the diamidine derivative diminazine, are associated with a high rate of toxic side effects. At best these drugs result in amelioration of clinical signs; rarely do they achieve true sterilisation of the infection.
The successful treatment of the small piroplasm infections, notably B. gibsoni, has been especially challenging. Clindamycin at 25 mg/kg q12 h PO induced morphological changes and reduced the parasitaemia in an experimental infection compared with untreated controls, but did not eliminate the parasite, and piroplasms were still observed in low numbers 108 days after infection [61]. Recently, the apparent cure of B. gibsoni was reported in 3 out of 4 experimental dogs that had not responded to repeated diminazine treatment, with a combination of clindamycin, metronidazole (15 mg/kg q12 h PO) and doxycycline (5 mg/kg q12 h PO) [62]. Successful treatment in these dogs was determined by a combination of normal clinical signs and the absence of B. gibsoni DNA in peripheral blood. More recently, a combination of azithromycin (10 mg/kg q24 h PO) and atovaquone (13.3 mg/kg q8 h PO) for 10 days has been used for treating B. gibsoni and appears to combine reasonable clinical efficacy with great safety [41]; the expense of atovaquone limits widespread acceptance of this therapy where it is most needed, in Asia. Cheaper formulations of atovaquone with proguanil cause an unacceptably high incidence of gastrointestinal side effects in dogs. Unfortunately this drug combination also does not result in a cure in some dogs, and rapid development of resistance to atovaquone caused by mutation of the cytochrome b gene has been reported [42, 63].
In vitro studies of numerous rainforest plant extracts for their antibabesial properties have been published recently [64–66], but to date no clinical trials have been reported with these compounds. A Haemaphysalis longicornis tick-derived peptide reduced parasitaemias of B. microti in BALB/c mice and directly kills B. gibsoni parasites in ticks [67].
Prevention of babesiosis, as with any tick-transmitted disease, is best achieved by removing the possibility of exposure to the vector. This is rarely achievable in endemic areas despite attentive ectoparasite control.
Several drugs have been investigated for their prophylactic potential against babesiosis, yet none has been consistently reliable in this regard. Experimental studies suggested that a single dose of imidocarb dipropionate (6 mg/kg) protects dogs from B. canis challenge for up to 8 weeks [68] and that doxycycline at 5 mg/kg/day ameliorates the severity of disease when challenged with virulent B. canis [69].
In vitro culture-derived soluble parasite antigens (SPA) have been used to protect dogs in European against B. canis (sensu strictu) challenge in a commercially available vaccine since the 1980s, and it was reported in one study that the incidence of babesiosis decreased from 16% to near zero in populations of vaccinated dogs living in endemic regions over the three-year period of study (reviewed in [70]). However, variable efficacy of the SPA vaccine during homologous challenge has been attributed to strain variation [71] and it was clear from early studies that B. canis-derived SPA did not protect dogs from heterologous challenge with B. rossi [72], which led to the development of a vaccine containing a mixture of SPA from both European B. canis and South African B. rossi that offered greater protection against heterologous challenge [73]. Current research is also conducted in Japan in search of suitable immunodominant and protective antigens for use in vaccines against Babesia gibsoni [74].
Conclusion
Canine babesiosis/piroplasmosis remains as a significant disease that despite considerable advances during the last decade in our knowledge and understanding of the pathogens themselves, the intricacies of transmission, and their pathophysiological mechanisms, still poses significant diagnostic and therapeutics challenges for veterinary practitioners around the world. New species of piroplasm will almost certainly be described, and the geographical range of established piroplasms will expand due to international movements of dogs and expansion of vector tick habitats. The challenges for the researchers are to provide practitioners with readily accessible and accurate diagnostic tools, safer and more efficacious anti-babesial drugs, and the "Holy Grail" of this applied research – effective immunologicals for the prevention or, at least, amelioration of the clinical signs of canine babesiosis.
References
Lobetti RG: Canine babesiosis. Comp Cont Ed Pract Vet. 1998, 20: 418-431.
Irwin PJ: Babesiosis and Cytauxzoonosis. Arthropod-borne infectious diseases of the dog and cat. Edited by: Shaw SE, Day MJ. 2005, London: Manson Publishing/The Veterinary Press, 63-77.
Jacobsen LS: The South African form of severe and complicated canine babesiosis: clinical advances 1994–2004. Vet Parasitol. 2006, 138: 126-139. 10.1016/j.vetpar.2006.01.047.
Birkenheuer AJ, Correa MT, Levy MG, Breitschwerdt EB: Geographic distribution of babesiosis among dogs in the United States and association with dog bites: 150 cases (2000–2003). J Am Vet Med Assoc. 2005, 227: 942-947. 10.2460/javma.2005.227.942.
Jefferies R, Ryan UM, Jardine J, Broughton DK, Robertson ID, Irwin PJ: Blood, bull terriers and babesiosis: further evidence for direct transmission of Babesia gibsoni in dogs. Aust Vet J. 2007, 85: 459-463. 10.1111/j.1751-0813.2007.00220.x.
Bostrom B, Wolf C, Greene C, Peterson DS: Sequence conservation in the rRNA first internal transcribed spacer region of Babesia gibsoni genotype Asia isolates. Vet Parasitol. 2008, 152: 152-157. 10.1016/j.vetpar.2007.11.024.
Reichenow E: Übertagungsweise und Entwicklung der Piroplasmen. Zbl Bakt I Orig. 1935, 135: 108-199.
Reichenow E: Über die Entwicklung von Theileria parva, dem Erreger des Küstenfiebers der Rinder, in Rhipicephalus appendiculatus. Zbl Bakt I Orig. 1937, 140: 223-226.
Zahler M, Schein E, Rinder H, Gothe R: Characteristic genotypes discriminate between Babesia canis isolates of differing vector specificity and pathogenicity in dogs. Parasitol Res. 1998, 84: 544-548. 10.1007/s004360050445.
Carret C, Walas F, Carcy B, Grande N, Précigout É, Moubri K, Schetters T, Gorenflot A: Babesia canis canis, Babesia canis vogeli, Babesia canis rossi: Differentiation of three subspecies by restriction fragment length polymorphism analysis on amplified small subunit ribosomal RNA genes. J Eukaryot Microbiol. 1999, 46: 298-303. 10.1111/j.1550-7408.1999.tb05128.x.
Birkenheuer AJ, Neel J, Ruslander D, Levy MG, Breitschwerdt EB: Detection and molecular characterization of a novel large Babesia species in a dog. Vet Parasitol. 2004, 124: 151-160. 10.1016/j.vetpar.2004.07.008.
Lehtinen LE, Birkenheuer AJ, Drolesky RE, Holman PJ: In vitro cultivation of a newly recognised Babesia sp. in dogs in North Carolina. Vet Parasitol. 2008, 151: 150-157. 10.1016/j.vetpar.2007.10.022.
Conrad P, Thomford J, Yamane I, Whiting J, Bosma L, Uno T, Holshuh HJ, Shelley S: Hemolytic anemia caused by Babesia gibsoni infections in dogs. J Am Vet Med Assoc. 1991, 199: 601-605.
Kjemtrup AM, Wainwright K, Miller M, Penzhorn BL, Carreno RA: Babesia conradae, sp. nov., a small canine Babesia identified in California. Vet Parasitol. 2006, 138: 103-111. 10.1016/j.vetpar.2006.01.044.
Zahler M, Rinder H, Schein E, Gothe R: Detection of a new pathogenic Babesia microti-like species in dogs. Vet Parasitol. 2000, 89: 241-248. 10.1016/S0304-4017(00)00202-8.
Camacho AT, Pallas E, Gestal JJ, Guitiàn FJ, Olmeda AS, Goethert HK, Telford SR: Infection of dogs in north-west Spain with a Babesia microti-like agent. Vet Rec. 2001, 149: 552-555.
Criado-Fornelio A, Martinez-Marcos A, Buling-Saraña A, Barba-Carretero JC: Molecular studies on Babesia, Theileria and Hepatozoon in southern Europe Part I: Epizootiological aspects. Vet Parasitol. 2003, 113: 189-201. 10.1016/S0304-4017(03)00078-5.
Criado A, Martinez J, Buling A, Barba JC, Merino S, Jefferies R, Irwin PJ: New data on epizootiology and genetics of piroplasms based on sequences of small ribosomal subunit and cytochrome b genes. Vet Parasitol. 2006, 142: 238-247. 10.1016/j.vetpar.2006.07.004.
Matjila PT, Leisewitz AL, Ooshuizen MC, Jongejan F, Penzhorn B: Detection of a Theileria species in dogs in South Africa. Vet Parasitol. 2008, 157: 34-40. 10.1016/j.vetpar.2008.06.025.
Uilenberg G: Babesia – a historical overview. Vet Parasitol. 2006, 138: 3-10. 10.1016/j.vetpar.2006.01.035.
Yamasaki M, Inokuma H, Sugimoto C, Shaw S, Aktas M, Yabsley MJ, Yamato O, Maede Y: Comparison and phylogenetic analysis of the heat shock protein 70 gene of Babesia parasites from dogs. Vet Parasitol. 2007, 145: 217-227. 10.1016/j.vetpar.2007.01.003.
Kjemtrup AM, Kocan AA, Whitworth L, Meinkoth J, Birkenheuer AJ, Cummings J, Boudreaux MK, Stockham SL, Irizarry-Rovira A, Conrad P: There are at least three genetically distinct small piroplasms from dogs. Int J Parasitol. 2000, 30: 1501-1505. 10.1016/S0020-7519(00)00120-X.
Goethert HK, Telford SR: What is Babesia microti?. Parasitology. 2003, 127: 301-309. 10.1017/S0031182003003822.
Mehlhorn H, Raether W, Schein E, Weber M, Uphoff M: Licht-und elektronenmikroskopische Untersuchungen zum Entwick-lungszyklus und Einfluss von Pentamidin auf die Morphologie der intraerythrocytären Stadien von Babesia microti. Dtsch Tierärztl Wochenschr. 1986, 93: 400-405.
Sasaki M, Omobowale O, Tozuka M, Ohta K, Matsuu A, Nottidge HO, Hirata H, Ikadai H, Oyamada T: Molecular survey of Babesia canis in dogs in Nigeria. J Vet Med Sci. 2007, 69: 1191-1193. 10.1292/jvms.69.1191.
Oyamada M, Davoust B, Boni M, Dereure J, Bucheton B, Hammad A, Itamoto K, Okuda M, Inokuma H: Detection of Babesia canis rossi, B. canis vogeli, and Hepatozoon canis in dogs in a village of Eastern Sudan by using a screening PCR and sequencing methodologies. Clin Diagn Lab Immunol. 2005, 12: 1343-1346. 10.1128/CDLI.12.11.1343-1346.2005.
Yeagley TJ, Reichard MV, Hempstead JE, Allen KE, Parsons LM, White MA, Little SE, Meinkoth JH: Detection of Babesia gibsoni and the small Babesia sp. 'Spanish isolate' in confiscated pit bull terriers. J Am Vet Med Assoc.
Trapp SM, Messick JB, Vidotto O, Jojima FS, Autran de Morais HS: Babesia gibsoni genotype Asia in dogs from Brazil. Vet Parasitol. 2006, 141: 177-180. 10.1016/j.vetpar.2006.04.036.
Solano-Gallego L, Trotta M, Carli E, Carcy B, Caldin M, Furlanello T: Babesia canis canis and Babesia canis vogeli clinicopathological findings and DNA detection by means of PCR-RFLP in blood from Italian dogs suspected of tick-borne disease. Vet Parasitol. 2008, 157: 211-221. 10.1016/j.vetpar.2008.07.024.
Irwin PJ, Hutchinson GW: Clinical and pathological findings of Babesia infection in dogs. Aust Vet J. 1991, 68: 204-209. 10.1111/j.1751-0813.1991.tb03194.x.
Böhm M, Leisewitz AL, Thompson PN, Schoeman JP: Capillary and venous Babesia canis rossi parasitaemias and their association with outcome of infection and circulatory compromise. Vet Parasitol. 2006, 141: 18-29. 10.1016/j.vetpar.2006.05.002.
Fukumoto S, Xuan X, Shigeno S, Kimbita E, Igarashi I, Nagasawa H, Fujisaki K, Mikami T: Development of a polymerase chain reaction methods for diagnosing Babesia gibsoni infection in dogs. J Vet Med Sci. 2001, 63: 977-981. 10.1292/jvms.63.977.
Jefferies R, Ryan U, Irwin P: PCR-RFLP for the detection and differentiation of the canine piroplasm species and its use with filter paper-based technologies. Vet Parasitol. 2007, 144: 20-27. 10.1016/j.vetpar.2006.09.022.
Birkenheuer AJ, Levy MG, Breitschwerdt EB: Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J Clin Microbiol. 2003, 41: 4172-4177. 10.1128/JCM.41.9.4172-4177.2003.
Duarte SC, Linhares GFC, Romanowsky TN, Neto O, Borges LMF: Assessment of primers designed for the subspecies-specific discrimination among Babesia canis canis, Babesia canis vogeli and Babesia canis rossi by PCR assay. Vet Parasitol. 2008, 152: 16-20. 10.1016/j.vetpar.2007.12.013.
Ikadai H, Tanaka H, Shibahara N, Matsuu A, Uechi M, Itoh N, Oshiro S, Kudo N, Igarashi I, Oyamada T: Molecular evidence of infections with Babesia gibsoni parasites in Japan and evaluation of the diagnostic potential of a loop-mediated isothermal amplification method. J Clin Microbiol. 2004, 42: 2465-2469. 10.1128/JCM.42.6.2465-2469.2004.
Georges K, Ezeokoli CD, Newaj-Fyzul A, Campbell M, Mootoo N, Mutani A, Sparagano OAE: The application of PCR and reverse line blot hybridization to detect arthropod-borne haemopathogens of dogs and cats in Trinidad. Ann N Y Acad Sci. 2008, 1149: 196-199.
Matsuu A, Ono S, Ikadai H, Uchide T, Imamura S, Onuma M, Okanao S, Higuchi S: Development of a SYBR green real-time polymerase chain reaction assay for quantitative detection of Babesia gibsoni (Asian genotype) DNA. J Vet Diagn Invest. 2005, 17: 569-573.
Tani H, Tada Y, Sasai K, Baba E: Improvement of DNA extraction method for dried blood spots and comparison of four methods for detection of Babesia gibsoni (Asian genotype) infection in canine blood samples. J Vet Med Sci. 2008, 70: 461-467. 10.1292/jvms.70.461.
Birkenheuer AJ, Levy MG, Breitschwerdt EB: Efficacy of combined atovaquone and azithromycin for therapy of chronic Babesia gibsoni (Asian genotype) infections in dogs. J Vet Intern Med. 2004, 18: 494-498. 10.1892/0891-6640(2004)18<494:EOCAAA>2.0.CO;2.
Jefferies R, Ryan UM, Jardine J, Robertson ID, Irwin PJ: Babesia gibsoni: Detection during experimental infections and after combined atovaquone and azithromycin therapy. Exp Parasitol. 2007, 117: 115-123. 10.1016/j.exppara.2007.03.016.
Goo Y, Jia H, Aboge GO, Terkawi MA, Kuriki K, Nakamura C, Kumagai A, Zhou J, Lee EG, Nishikawa Y: Babesia gibsoni: Serodiagnosis of infection in dogs by an enzyme-linked immunosorbent assay with recombinant BgTRAP. Exp Parasitol. 2008, 118: 555-560. 10.1016/j.exppara.2007.11.010.
Anderson JF, Magnarelli LA, Sulzer AJ: Canine babesiosis: Indirect fluorescent antibody test for a North American isolate of Babesia gibsoni. Am J Vet Res. 1980, 41: 2102-2105.
Levy MG, Breitschwerdt EB, Moncol DJ: Antibody activity to Babesia canis in dogs in North Carolina. Am J Vet Res. 1987, 48: 339-341.
Aboge GO, Jia H, Terkawi MA, Goo Y, Kuriki K, Nishikawa Y, Igarashi I, Suzuki H, Xuan X: A novel 57-kDa merozoite protein of Babesia gibsoni is a prospective antigen for diagnosis and serosurvey of canine babesiosis by enzyme-linked immunosorbent assay. Vet Parasitol. 2007, 149: 85-94. 10.1016/j.vetpar.2007.06.025.
Fukumoto S, Xuan X, Nishikawa Y, Inoue N, Igarashi I, Nagasawa H, Fujisaki K, Mikami T: Identification and expression of a 50-kilodalton surface antigen of Babesia gibsoni and evaluation of its diagnostic potential in an enzyme-linked immunosorbent assay. J Clin Microbiol. 2001, 39: 2603-2609. 10.1128/JCM.39.7.2603-2609.2001.
Fukumoto S, Xuan X, Inoue N, Igarashi I, Sugimoto C, Fujisaki K, Nagasawa H, Mikami T, Suzuki H: Molecular characterization of a gene encoding a 29-kDa cytoplasmic protein of Babesia gibsoni and evalution of its diagnostic potentiality. Mol Biochem Parasitol. 2003, 131: 129-136. 10.1016/S0166-6851(03)00199-3.
Zhou J, Fukumoto S, Jia H, Yokoyama N, Zhang G, Fujisaki K, Lin J, Xuan X: Characterization of the Babesia gibsoni P18 as a homologue of thrombospondin related adhesive protein. Mol Biochem Parasitol. 2006, 148: 190-198. 10.1016/j.molbiopara.2006.03.015.
Jia H, Zhou J, Ikadai H, Matsuu A, Suzuki H, Igarashi I, Fujisaki K, Xuan X: Identification of a novel gene encoding a secreted antigen 1 of Babesia gibsoni and evaluation of its use in Serodiagnosis. Am J Trop Med Hyg. 2006, 75: 843-850.
Aboge GO, Jia H, Kuriki K, Zhou J, Nishikawa Y, Igarashi I, Fujisaki K, Suzuki H, Xuan X: Molecular characterization of a novel 32-kDa merozoite antigen of Babesia gibsoni with a better diagnostic performance by enzyme-linked immunosorbent assay. Parasitology. 2007, 134: 1185-1194. 10.1017/S0031182007002594.
Zhou J, Jia H, Nishikawa Y, Fujisaki K, Xuan X: Babesia gibsoni rhoptry-associated protein 1 and its potential use as a diagnostic antigen. Vet Parasitol. 2007, 145: 16-20. 10.1016/j.vetpar.2006.10.022.
Konishi K, Sakata Y, Miyazaki N, Jia H, Goo YK, Xuan X, Inokuma H: Epidemiological survey of Babesia gibsoni infection in dogs in Japan by enzyme-linked immunosorbent assay using B. gibsoni thrombospondin-related adhesive protein antigen. Vet Parasitol. 2008, 155: 204-208. 10.1016/j.vetpar.2008.04.025.
Birkenheuer AJ: Personal communication. 2009
Máthé A, Dobos-Kovacs M, Voros K: Histological and ultra-structural studies of renal lesions in Babesia canis infected dogs treated with imidocarb. Acta Vet Hung. 2007, 55: 511-523. 10.1556/AVet.55.2007.4.10.
Camacho AT, Guitian FJ, Pallas E, Gestal JJ, Olmeda AS, Goethert HK, Telford SR, Spielman A: Azotaemia and mortality among Babesia-microti-like infected dogs. J Vet Int Med. 2004, 18: 141-146. 10.1892/0891-6640(2004)18<141:AAMABM>2.0.CO;2.
Lobetti RG, Jacobsen LS: Renal involvement in dogs with babesiosis. J Sth Afr Vet Assoc. 2001, 72: 23-28.
de Scally MP, Lobetti RG, Reyers F, Humphris D: Are urea and creatinine values reliable indicators of azotaemia in canine babesiosis?. J Sth Afr Vet Assoc. 2004, 75: 121-124.
Schoeman JP, Rees P, Herrtage ME: Endocrine predictors of mortality in canine babesiosis caused by Babesia canis rossi. Vet Parasitol. 2007, 148: 75-82. 10.1016/j.vetpar.2007.06.010.
Brandao LP, Hagiwara MK, Myiashiro SI: Humoral immunity and reinfection resistance in dogs experimentally inoculated with Babesia canis and either treated or untreated with imidocarb dipropionate. Vet Parasitol. 2003, 114: 253-265. 10.1016/S0304-4017(03)00130-4.
Wulansari R, Wijaya A, Ano H, Horii Y, Nasu T, Yamane S, Makimura S: Clindamycin in the treatment of Babesia gibsoni infections in dogs. J Am Anim Hosp Assoc. 2003, 39: 558-562.
Suzuki K, Wakabyashi H, Takahashi M, Fukushima K, Yabuki A, Endo Y: A possible treatment strategy and clinical factors to estimate the treatment response in Babesia gibsoni infection. J Vet Med Sci. 2007, 69: 563-568. 10.1292/jvms.69.563.
Matsuu A, Miyamato K, Ikada H, Okano S, Higuchi S: Short Report: Cloning of the Babesia gibsoni cytochrome B gene and isolation of three single nucleotide polymorphisms from parasites present after atovaquone treatment. Am J Trop Med. 2006, 74: 593-597.
Subeki , Matsuura H, Yamasaki M, Yamato O, Maede Y, Katakura K, Suzuki M, Trimurningsih , Chairul , Yoshihara T: Effects of central Kalimantan plant extracts on intraerythrocytic Babesia gibsoni in culture. J Vet Med Sci. 2004, 66: 871-874. 10.1292/jvms.66.871.
Subeki , Matsuura H, Takahashi K, Yamasaki M, Yamato O, Maede Y, Katakura K, Suzuki M, Trimurningsih , Chairul , Yoshihara T: Antibabesial activity of protoberberine alkaloids and 20-hydroxyecdysone from Arcangelisia flava against Babesia gibsoni in culture. J Vet Med Sci. 2005, 67: 223-227. 10.1292/jvms.67.223.
Subeki , Matsuura H, Takahashi K, Yamasaki M, Yamato O, Maede Y, Katakura K, Kobayashi S, Trimurningsih , Chairul , Yoshihara T: Anti-babesial and anti-plasmodial compounds from Phyllanthus niruri. J Nat Prod. 2005, 68: 537-539. 10.1021/np0497245.
Tsuji N, Battsetseg B, Boldbaatar D, Miyoshi T, Xuan X, Oliver J, Fujisaki K: Babesial vector tick defensin against Babesia sp. parasites. Infect Immun. 2007, 75: 3633-3640. 10.1128/IAI.00256-07.
Uilenberg G, Verdiesen AHM, Zwart D: Imidocarb: A chemoprophylactic experiment with Babesia canis. The Vet Quarterly. 1981, 3: 118-123.
Vercammen F, de Deken R, Maes L: Prophylactic treatment of experimental canine babesiosis (Babesia canis) with doxy-cycline. Vet Parasitol. 1996, 66: 251-255. 10.1016/S0304-4017(96)01016-3.
Schetters T: Vaccination against canine babesiosis. Trends in Parasitol. 2005, 21: 179-184. 10.1016/j.pt.2005.02.006.
Carcy B, Precigout E, Schetters T, Gorenflot A: Genetic basis for GPI-anchor merozoite surface antigen polymorphism of Babesia and resulting antigenic diversity. Vet Parasitol. 2006, 138: 33-49. 10.1016/j.vetpar.2006.01.038.
Schetters T, Kleuskens J, Scholtes N, Bos HJ: Strain variation limits protective activity of vaccines based on soluble Babesia canis antigens. Parasite Immunol. 1995, 17: 215-218. 10.1111/j.1365-3024.1995.tb00891.x.
Schetters TPM, Kleuskens JAGM, Scholtes NC, Gorefnflot A, Moubri K, Vermeulen AN: Vaccination of dogs against heterologous Babesia canis infection using antigens from culture supernatants. Vet Parasitol. 2001, 100: 75-86. 10.1016/S0304-4017(01)00485-X.
Fukumoto S, Tamaki Y, Shirafuji H, Harakawa S, Suzuki H, Xuan X: Immunization with recombinant surface antigen P50 of Babesia gibsoni expressed in insect cells induced parasite growth inhibition in dogs. Clin Diagn Lab Immunol. 2005, 12: 557-559. 10.1128/CDLI.12.4.557-559.2005.
Acknowledgements
This article is published as part of Parasites & Vectors Volume 2 Supplement 1, 2009: Proceedings of the 4th International Canine Vector-Borne Disease Symposium. The full contents of the supplement are available online at http://www.parasitesandvectors.com/supplements/2/S1.
Publication of the supplement has been sponsored by Bayer Animal Health GmbH.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author declares that they have no competing interests.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Irwin, P.J. Canine babesiosis: from molecular taxonomy to control. Parasites Vectors 2 (Suppl 1), S4 (2009). https://doi.org/10.1186/1756-3305-2-S1-S4
Published:
DOI: https://doi.org/10.1186/1756-3305-2-S1-S4