
Abstract
Background
Staphylococcus aureus and Coagulase-negative staphylococci (CoNS) are a major source of infections associated with indwelling medical devices. Many antiseptic agents are used in hygienic handwash to prevent nosocomial infections by Staphylococci. Our aim was to determine the antibiotic susceptibility and resistance to quaternary ammonium compound of 46 S. aureus strains and 71 CoNS.
Methods
S. aureus (n = 46) isolated from auricular infection and CoNS (n = 71), 22 of the strains isolated from dialysis fluids and 49 of the strains isolated from needles cultures were investigated. Erythromycin resistance genes (erm A, erm B, erm C, msr A and mef) were analysed by multiplex PCR and disinfectant-resistant genes (qac A, qac B, and qac C) were studied by PCR-RFLP.
Results
The frequency of erythromycin resistance genes in S. aureus was: erm A+ 7.7%, erm B+ 13.7%, erm C+ 6% and msr A+ 10.2%. In addition, the number of positive isolates in CoNS was respectively erm A+ (9.4%), erm B+ (11.1%), erm C+ (27.4%), and msr A+ (41%). The MIC analyses revealed that 88 isolates (74%) were resistant to quaternary ammonium compound-based disinfectant benzalkonium chloride (BC). 56% of the BC-resistant staphylococcus isolates have at least one of the three resistant disinfectants genes (qac A, qac B and qac C). Nine strains (7.7%) among the CoNS species and two S. aureus strains (2%) harboured the three-qac genes. In addition, the qac C were detected in 41 strains.
Conclusions
Multi-resistant strains towards macrolide and disinfectant were recorded. The investigation of antibiotics and antiseptic-resistant CoNS may provide crucial information on the control of nosocomial infections.
Similar content being viewed by others


Background
The increasing number of infections caused by oxacillin-resistant staphylococci makes glycopeptide antibiotics an important choice [1]. The significant prevalence of nosocomial infections caused by multi-resistant S. aureus and coagulase-negative staphylococci (CoNS) has been documented [2, 3]. These species have the ability to survive in medical devices for months [4]. Resistance to erythromycin in staphylococci is usually associated with resistance to other macrolides. Three genes (erm A, erm B, and erm C) encoding methyltransferases responsible of resistance to macrolides, lincosamides and type B streptogramins (MLSB phenotype) by modification of the ribosomal target site have been found in staphylococci [5]. The msr A gene displays another mechanism of inducible resistance to erythromycin by encoding an ATP-dependent efflux pump [6]. On the other hand, macrolide efflux is affected by a membrane protein encoded by the mef gene [7]. Antiseptic agents include various compounds with different chemical structures [8]. The widespread use of quaternary ammonium compounds (QAC) in hospitals actually contributes to the emergence of disinfectant-resistant bacteria [9, 10]. Epidemiological data on antiseptic susceptibility and the distribution of resistance genes are both useful for nosocomial infection control. In several staphylococcal species, qac-resistant genes have been identified [11–13]. Found in clinical staphylococci (qac A, qac B, and qac C), these genes are generally carried by plasmids [14–16]. Some of these plasmids (pST6, pSK4, and pSK41) contain antibiotic resistance genes encoding resistance to gentamicin, penicillin, kanamycin, and tobramycine [17, 18]. Multidrug resistance pumps have been recognized as mediators of a number of commonly used ammonium antiseptics and detergents [19]. The nearly identical qac A/B gene is normally harboured by large plasmids [20, 21]. The relation between qac resistance and penicillin resistance in human clinical staphylococci is quite prevalent [22].
In the present study, we examined the antibiotic susceptibility and resistance to quaternary ammonium compound-based disinfectant benzalkonium chloride (BC) of 46 S. aureus strains isolated from auricular infection and 71 CoNS isolated from dialysis fluids and needles cultures. In order to examine the genetic drug resistance mechanisms, erythromycin resistance genes (erm A, erm B, and erm C) and macrolide efflux genes (msr A and mef) were analysed by PCR multiplex. In addition, quaternary ammonium resistance genes (qac A, qac B, and qac C) were detected by PCR-RFLP.
Methods
Biochemical characterization and antimicrobial susceptibility
A total of 117 clinical staphylococcal strains were isolated from Kairouan, in central of Tunisia including 46 S. aureus strains isolated from auricular infection and 71 CoNS strains isolated from dialysis fluid and needles from a dialysis service. All strains were identified using the Api ID32 Staph system (bioMérieux, Marcy l'Étoile, France) according to the manufacturer's recommendations and the results were read using an automated microbiological mini-Api (bioMérieux, Marcy l'Étoile, France). Each strain was tested for 18 antibiotics (penicillin, oxacillin, kanamycin, tobramycin, gentamicin, tetracycline, minocyclin, erythromycin, lincomycin, pristinamycin, fosfomycin, nitrofurantoin, pefloxacin, rifampicin, fusidic acid, vancomycin, teicoplanin and cotrimoxazol) using the ATB Staph system (bioMérieux, France) according to the manufacturer's specifications.
Minimum inhibitory concentration determination
The broth microdilution method was used to determine the minimum inhibitory concentration (MIC) of benzalchonium chloride (BC) (Acros organics, USA) (ranging from 0.5 to 256 μg/ml, by serial twofold dilutions) against the tested strains as recommended by the Clinical and Laboratory Standards Institute (CLSI) [23]. An inoculum of 104 -105 cells ml-1 was used. The lowest concentration of antimicrobial agent totally preventing growth after 24 h was taken to be the MIC.
Multiplex PCR for the detection of macrolide-resistance encoding genes
Macrolide resistance genes erm A, erm B, erm C, msr A, and mef were examined in all strains using multiplex PCR, as previously described [24, 25]. Multiplex PCR assays were performed in 25 μl PCR mixture 1 and mixture 2. Mixture 1 contained a DNA template (50 ng), 100 μM concentrations (each) of the four dNTPs, 1 U of Go Taq DNA polymerase (Promega, Lyon, France), 5 μL green Go Taq buffer (5X), 25 pM each of forward and reverse primers erm A, erm C, and msr A (Table 1). The PCR mixtures were subjected to thermal cycling (3 min at 96°C, followed by 30 cycles of 30 sec at 95°C for denaturation, 30 sec at 55°C for annealing extension, and extension at 72°C for 2 min). A final elongation at 72°C for 10 min was achieved in a DNA thermal cycler (GenAmp PCR system 9700-Applied Biosystem Int., USA). For mixture 2, the forward and reverse primers (25 pM) of genes erm B and mef were used. PCR products were analysed in agarose gel (1%) electrophoresis in 1X Tris-borate-EDTA buffer (TBE) at pH 8.3. The amplification products were photographed and their size was determined using a 100 bp molecular size marker (Bio-Rad, France).
Detection of qac A, qac B, and qac C genes by PCR-RFLP
Multiplex PCR-RFLP analysis of the qac A/B and qac C genes was achieved as described by Sekiguchi et al. [26]. For the fragment qac A/qac B, the forward primer (corresponding to nucleotides 924-946) and the reverse primer (corresponding to nucleotides 1143-1124) produced 220 bp. For the qac C gene, the forward primer (nucleotides 73-97) and the reverse primer (nucleotides 321-302) produced 249 bp. PCR was performed in a 25 μl reaction volume containing 50 ng of extracted DNA, 5 μl green Go Taq buffer (5X), 200 μM each of deoxynucleoside triphosphates (dNTP), 25 μM each of qac A, qac B, and qac C forward and reverse primers, 1 U of GO Taq DNA polymerase (Promega, USA). Each PCR was performed in duplicate. The reaction mixtures were heated to 94°C for 5 min and were then subjected to 30 denaturation cycles at 94°C for 1.5 min, annealing at 56°C for 0.5 min and extension at 72°C for 1.5 min, ending with a final extension at 72°C for 10 min.
Following PCR, the product was digested with 5 U of Alu I at 37°C for 90 min (Promega, France). Ten microlitres of treated product were analysed by gel electrophoresis in 3% agarose gel in 1X Tris-borate-EDTA buffer (TBE, pH 8.3). The amplification products were photographed and their size was determined using a 25 bp molecular size marker (Promega, USA). The qac B gene was expected to be digested into two fragments: a visible 176 bp fragment and another fragment of 44 bp which is invisible. Qac A and qac C genes were not expected to be digested [26].
Statistical analysis
Statistical analysis was performed on SPSS v.13.0 statistics software. Pearson's chi-square-2 test was used to asses inter-group significance. In addition statistical significance was set at P < 0.05.
Results
Biotyping and antimicrobial susceptibility
In this study, 46 S. aureus strains isolated from auricular infection were identified. In addition 71 CoNS were isolated and were subdivided into eight species: S. epidermidis (n = 32) (45%) followed by S. hominis (n = 10) (14.1%), S. haemolyticus (n = 9) (12.7%), S. warneri (n = 5) (7%), S. simulans (n = 6) (8.5%), S. capitis (n = 4) (5.6%), S. chromogenes (n = 3) (4.2%), and S. equorum (n = 2) (2.8%).
The results of the antibiotic susceptibility test confirmed the multi-resistance of 117 staphylococcal strains toward the 18 antibiotics mentioned previously. The majority of these strains were resistant to penicillin (91.1%). The isolated strains were also resistant to kanamycin (45.7%), tetracycline (47.8%), erythromycin (37.7%), lincomycin (21.2%), fosfomycin (20.8%), fusidic acid (17.3%), pefloxacin (18.4%), cotrimoxazol (24.3%), teicoplanin (10.5%), gentamicin (13.4%), rifampicin (12.1%), and tobraycin (16.8%).
Oxacillin-resistant phenotype was found in 9 S. aureus (7.7%) and 24 CoNS (20.5%) isolated strains. All of the strains were susceptible to pristinamycin, nitrofurantoin, and vancomycin (Table 2 and 3).
Resistance to disinfectants agents
The 117 staphylococcus isolates were screened for QAC (BC) resistance. The strains were categorized as BC resistant or sensitive according to the BC MICs. Twenty four (20%) isolates were considered BC highly resistant (BC MICs between 16 and 32 μg/ml), 64 (54%) isolates were resistant to BC (BC MICs between 4 and 8 μg/ml), and 28 (23%) strains were sensitive to BC (BC MICs ≤ 2 μg/ml). 117 staphylococci isolates were analyzed for correlation between BC and antibiotic resistance (Figure 1). This analysis showed that the frequency of erythromycin resistance 71% and oxacillin resistance 84% was higher among the BC-resistant strains.

Percentages of antibiotic-resistance Staphylococci isolates among BC-resistance and BC-sensitive isolates. Antibiotics used are erythromycin (ERY) and oxacillin (OXA).
Multiplex PCR for the detection of genes encoding macrolide resistance
In this study we found that the incidence of the three erythromycin ribosomal methylase genes tested was 9 (7.7%) strains of S. aureus contained erm A, 16 (13.7%) strains harbored erm B gene and 7 (6%) strains were positive for ermC in the total of 117 isolated strains (Figure 2A, B). Furthermore among the CoNS 11 (9.4%) strains contained erm A. In addition, on the total of 117 isolated strains 13 CoNS strains carried the erm B gene, while 32 (27.3%) were positive for erm C. The msrA gene was present in 12 (10.2%) of S. aureus strains and in 48 (41%) CoNS strains (Figure 2A). In contrast, the mef gene was absent in all the staphylococcal strains tested. Sixteen strains of S. epidermidis, eight strains of S. aureus and eighteen strains of CoNS were positive for the mec A gene, yet were susceptible to oxacillin. Furthermore, only nine strains of CoNS (7.7%) and nineteen strains (16.2%) of S. aureus were susceptible to erythromycin, those isolated did not contain any of the erythromycin-resistance genes tested. However, in five S. epidermidis strains (E7, E20, S23, S27, and S40), and in ten S. aureus stains, the erm A and erm C genes were not detected, although it was resistant to erythromycin (Table 2 and 3).

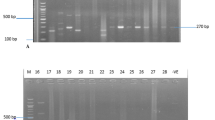
(A) PCR analysis of erythromycin-resistant determinants: erm A (139 bp), msr A (163 bp) and erm C (190 bp) with clinical strains; source of DNA: 1: 100 bp DNA molecular size marker; 2: negative control; 3: S16; 4: E6; 5: E18; 6: S40; 7: E24; 8: E20; 9: S22; 10: S12; 11: S9; 12: S21 and 13: E15. (B) Agarose gel electrophoresis of PCR amplicon of erythromycin-resistant erm B gene (142 bp) obtained with DNA of clinical strains; 1: 100 bp DNA molecular size marker; 2: negative control; 3: E10 and 4: E11.
PCR-RFLP detection of qac A, qac B, and qac C genes
Among the tested S. aureus two strains (Sa22 and Sa23) carried the three qac genes (qac A, qac B and qac C) (Figure 3.). While, nine CoNS strains, contained the three qac genes (qac A, qac B and qac C). In addition, qac C was the most present (35%), followed by qac A (24%), and qac B (15.4%) in the total of 117 isolated strains (Table 2 and 3).

PCR-RFLP analysis for the detection of qac A (220 bp), qac B (174 bp), and qac C (249 bp) genes in S.aureus ; 1: 25 bp DNA molecular size marker; 2: negative control; 3: Sa1; 4: Sa5; 5: Sa10; 6: Sa13; 7: Sa19; 8: Sa20; 9: Sa22 and 10: Sa23.
Qac-resistant genes (qac A) were identified in (5.12%) followed by qac C (4.27%) and qac B (3.41%) in the isolated S. aureus strains, while 39 (33%) strains were qac-negative in the total of 117 isolated strains (Figure 4.). On the other hand, among the 32 S. epidermidis strains under study, five were qac A+, two were qac B+, and 16 were qac C+, while 29 strains were qac-negative (Table 2 and 3). The three Qac-resistant genes (qac A, qac B, and qac C) were identified in 9.4% of the total isolated strains. Nine strains (7.7%) of six different CoNS species harboured the three qac-resistant genes.

PCR-RFLP analysis for the detection of qac A (220 bp), qac B (174 bp), and qac C (249 bp) in CoNS; 1: 25 bp DNA molecular size marker; 2: negative control; 3: S9; 4: E15; 5: E13; 6: E6; 7: S18; 8: E7; 9: S13; 10: S19; 11: S11; 12: S45 and 13: S39.
Discussion
Recently, the increasing numbers of device-related infections associated with methicillin-resistant staphylococci have raised awareness toward the need for alternative agents to prevent these infections. S. epidermidis species represent the CoNS most recovered from clinical specimens [3]. Among the 71 CoNS isolated in this study, the most prevalent were S. epidermidis (n = 32), S. hominis (n = 10), and S. haemolyticus (n = 9). The antibiotic susceptibility of the 117 staphylococal (S.aureus and CoNS) isolated in this study confirmed the multi-resistance of these strains toward the 18 antibiotics cited previously. Oxacillin resistance occurred in 7.7% of S. aureus and in 20.5% of CoNS tested by ATB Staph.
Erythromycin resistance in staphylococci is predominantly mediated by erythromycin-resistant methylase encoded by erm genes [27]. The inducible gene ermA is found on the transposon Tn554 and has a single specific site for insertion into the S. aureus chromosome [28]. The ermB gene is found on the transposon Tn551 of a penicillinase plasmid [29]. The ermC gene is responsible for constitutive or inducible resistance to erythromycin and is generally located on small plasmids [5, 27, 30]. On the other hand, the investigation of the prevalence of erm A, erm B, erm C and msr A genes in Staphylococci showed that only 28 strains of S. aureus (n = 46) and CoNS (n = 71) were found to be susceptible to erythromycin, yet contained erythromycin-resistant gene, and 44 strains (13 S.aureus and 31 CoNS) have at least one of the four genes (ermA, ermB, ermC and msrA) and were susceptible to erythromycin. Similarly, Sekiguchi et al. [26] found discordance among phenotypic susceptibility and the presence of erm genes. They stated that this discordance might be due to a mutation in the coding or promoter region of the PCR-detected genes. We noted also that eight strains of S.aureus and CoNS strains were found to be resistant to erythromycin but did not carry any erythromycin resistance gene (Table 1 and 2). This result may be explained by the location of these genes in small plasmids, which were occasionally lost. Fluit et al. [31] demonstrated that the erm C gene responsible for erythromycin resistance is located on a small plasmid. In this study, the incidences of ermA in erythromycin-resistant staphylococci were 7.7% for S. aureus and 3.4% for S. epidermidis. These findings are in disagreement with the study by Eady et al. [5] conducted with coagulase-negative staphylococci (CoNS) in the United Kingdom in which an incidence of 5.9% for ermA was reported. In a study performed in Denmark, 16% of S. aureus strains were carrying ermA, while only 3% of CoNS strains had this gene [30]. Regarding erm B, we found that this gene was more frequently encountered than erm A in erythromycin-resistant staphylococci with 24.8% (13.7% for S.aureus and 11% for CoNS) of total strains carrying erm B. In the United Kingdom, an incidence of 7.2% for erm B in CoNS has been reported [5]. Staphylococcal strains resistant to macrolides and type-B streptogramins frequently harbour msr A, which encodes an ATP-dependent efflux pump [5]. Erythromycin resistance may be caused by the msr A or erm B gene, as previously reported with staphylococci [5]. Our results are similar to those of a recent study investigating a high level of erm A and erm C genes in CoNS [32].
Staphylococcal multidrug-resistant gene qac A is generally mediated by plasmids mediated resistance to various toxic organic cations and ethidium bromide, as well as a number of commonly used antiseptics and disinfectants, such as benzalkonium chloride and chlorhexidine [19]. The qac A gene also encodes resistance to both monovalent and divalent organic cations. In addition, qac B characteristically differs from qac A by conferring lower or no resistance to divalent organic cations [33]. Some investigations have implied that there is disinfectant cross-resistance with antibiotics [34, 35].
Among the tested bacteria, 2 strains of S.aureus and 9 strains of CoNS carried the three qac genes (qac A, qac B, and qac C), as shown in Table 1 and 2. Qac C gene was the most present in the 117 isolated strains (35%), followed by qac A (24%), and qac B (15. 4%). Of the 117 isolates investigated in this study, 74% were phenotypically resistant to BC. 56% of the BC-resistant staphylococcus isolates have at least one of the three resistant disinfectants genes (qacA/B and qacC). Previous investigators have reported a similar distribution of these three qac resistance genes in clinical S. aureus and CoNS [36, 15]. Little is known about the occurrence and possible genetic linkage of qac and antibiotic resistance in staphylococci. Interestingly, we observed that staphylococci resistant to BC were generally more often resistant to antibiotics (Ery and Oxa) than BC-sensitive isolates (Figure 1).
Furthermore, among the nine MRSA isolates, two strains (Sa22 and Sa23) were multi-resistant to antibiotic and harboured the qac genes. MRSA isolates resistant to antiseptics and disinfectants have been reported in Australia and in United Kingdom in the last decade [8]. Sekiguchi et al., [26] have found that among the 65 MRSA isolates 32 (49.23%) were positive for qac A, while one isolate was positive for qac B and seven MRSA 10% were positive for qac C. Three strains of S. hemolyticus harboured qac A and qac B genes. In a recent study, S. hemolyticus isolates were shown to contain both qac A and qac B genes [37]. Qac A/B genes are typically located on a transposon of transmissible multidrug-resistant plasmids, such as pSK1 [33]. Staphylococci resistant genes to quaternary ammonium compounds (QACs) have been detected in clinical coagulase-negative staphylococci [36]. QAC resistant Staphylococcus spp. hosting qac A/qac B and smr have been isolated from different environments [38]. Results from a recent study in Norway suggest that qac-resistant genes are common in human clinical staphylococci and that a direct link between resistance to QACs and resistance to penicillin occurs in clinical isolates of human and animal origin as well as in food-related staphylococci [11, 22]. Noguchi et al. [16] reported that when the antiseptic susceptibility and the distribution of antiseptic-resistant genes of MRSA isolated in Japan in 1992 were studied, qac A/B were detected in 10.2% (10/98). However, seven years later, qac A/B genes were detected in 47.9% (198/413) in MRSA isolates in Japan.
Conclusion
It appears that the widespread distribution of staphylococci carrying macrolides and qac-resistant genes found in dialysis biomaterial collected in Tunisia may be due to the transfer of resistance plasmids among species and strains, thereby contributing to the dissemination of staphylococcal resistance. Therefore, a closer investigation of antibiotics and antiseptic-resistant CoNS may provide crucial information on the control of nosocomial infections.
References
Hiramatsu K: Vancomycin resistance in staphylococci. Drug Resist Updat. 1998, 1: 135-150. 10.1016/S1368-7646(98)80029-0.
Brun-Buisson CH: Les Staphylococcus aureus résistants à la méticilline: Evolution et épidémiologie, impact clinique, prévention. Pathol Biol. 1998, 46: 227-34.
Kloos WE, Bannerman TL: Update on clinical significance of coagulase-negative staphylococci. Clin Microbiol Rev. 1994, 7: 117-140.
Neely AN, Maley MP: Survival of enterococci and staphylococci on hospital fabrics and plastic. J Clin Microbiol. 2000, 38: 724-726.
Eady EA, Ross JI, Tipper JL, Walters CE, Cove JH, Noble WC: Distribution of genes encoding erythromycin ribosomal methylases and an erythromycin efflux pump in epidemiologically distinct groups of staphylococci. J Antimicrob Chemother. 1993, 31: 211-7. 10.1093/jac/31.2.211.
Ross JI, Eady EA, Cove JH, Baumberg S: Identification of a chromosomally encoded ABC-transport system with which the staphylococcal erythromycin exporter Msr A may interact. Gene. 1995, 153: 93-98. 10.1016/0378-1119(94)00833-E.
Leclercq R: Mechanisms of resistance to macrolides and lincosamides: nature of the resistance elements and their clinical implications. Clin Infect Dis. 2002, 34: 482-492. 10.1086/324626.
McDonnell G, Russell AD: Antiseptics and disinfectants: activity, action, and resistance. Clin Microbiol Rev. 1999, 12: 147-179.
Reverdy ME, Bes M, Brun Y, Fleurette J: Evolution of resistance to antibiotics and antiseptics of hospital Staphylococcus aureus strains isolated from 1980 to 1991. Pathol Biol. 1993, 41: 897-904.
Russell AD: Do biocides select for antibiotic resistance?. J Pharm Pharmacol. 2000, 52: 227-233.
Anthonisen IL, Sunde M, Steinum TM, Sidhu MS, Sorum H: Organization of the antiseptic resistance gene qac A and Tn552-related βlactamase genes in multidrug-resistant Staphylococcus haemolyticus strains of animal and human origins. Antimicrob. Agents Chemother. 2002, 46: 3606-3612. 10.1128/AAC.46.11.3606-3612.2002.
Bjorland J, Steinum T, Sunde M, Waage S, Heir E: Novel plasmid-borne gene qacJ mediates resistance to quaternary ammonium com pounds in equine Staphylococcus aureus, Staphylococcus simulans, and Staphylococcus intermedius. Antimicrob Agents Chemother. 2003, 47: 3046-3052. 10.1128/AAC.47.10.3046-3052.2003.
Heir E, Sundheim G, Holck AL: The qacG gene on plasmid pST94 confers resistance to quaternary ammonium compounds in staphylococci isolated from the food industry. J Appl Microbiol. 1999, 86: 378-388. 10.1046/j.1365-2672.1999.00672.x.
Leelaporn A, Firth N, Paulsen IT, Hettiaratchi A, Skurray RA: Multidrug resistance plasmid pSK108 from coagulase-negative staphylococci; relationships to Staphylococcus aureus qacC plasmids. Plasmid. 1995, 34: 62-67. 10.1006/plas.1995.1034.
Mayer S, Boos M, Beyer A, Fluit AC, Schmitz FJ: Distribution of the antiseptic resistance genes qac A, qac B and qac C in 497 methicillin- resistant and susceptible European isolates of Staphylococcus aureus. J Antimicrob Chemother. 2001, 47: 896-897. 10.1093/jac/47.6.896.
Noguchi N, Hase M, Kitta M, Sasatsu M, Deguchi K, Kono M: Antiseptic susceptibility and distribution of antiseptic-resistance genes in methicillin-resistant Staphylococcus aureus. FEMS Microbiol Lett. 1999, 172: 247-253. 10.1111/j.1574-6968.1999.tb13475.x.
Berg T, Firth N, Apisiridej S, Hettiaratchi A, Leelaporn A, Skurray RA: Complete nucleotide sequence of pSK41:evolution of staphylococcal conjugative multiresistance plasmids. J Bacteriol. 1998, 180: 4350-4359.
Sidhu MS, Heir E, Sorum H, Holck A: Genetic linkage between resistance to quaternary ammonium compounds and β lactam antibiotics in food-related Staphylococccus spp. Microb Drug Resis. 2001, 7: 363-371. 10.1089/10766290152773374.
Littlejohn TG, Paulsen IT, Gillespie MT, Tennent JM, Midgley M, Jones IG, Purewal AS, Skurray RA: Substrate specificity and energetics of antiseptic and disinfectant resistance in Staphylococcus aureus. FEMS Microbiol Lett. 1992, 74: 259-265.
Littlejohn TG, DiBerardino D, Messerotti LJ, Spiers SJ, Skurray RA: Structure and evolution of a family of genes encoding antiseptic and disinfectant resistance in Staphylococcus aureus. Gene. 1991, 101: 59-66. 10.1016/0378-1119(91)90224-Y.
Paulsen IT, Brown MH, Dunstan SJ, Skurray RA: Molecular characterization of the staphylococcal multidrug resistance export protein QacC. J Bacteriol. 1995, 177: 2827-2833.
Sidhu MS, Heir E, Leegaard T, Wiger K, Holck A: Frequency of disinfectant resistance genes and genetic linkage with βlactamase transposon Tn552 among clinical staphylococci. Antimicrob Agents Chemother. 2002, 46: 2797-2803. 10.1128/AAC.46.9.2797-2803.2002.
CLSI: Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. Approved Standard-Seventh Edition CLSI Document M7-A7. 2006
Martineau F, Picard FJ, Lansac N, Ménard C, Roy PH, Ouellette M, Bergeron MG: Correlation between the resistance genotype determined by multiplex PCR assays and the antibiotic susceptibility patterns of Staphylococcus aureus and Staphylococcus epidermidis. Antimicrob Agents Chemother. 2000, 231-238.
Lim JA, Kwon AR, Kim SK, Chong Y, Lee K, Choi EC: Prevalence of resistance to macrolide, lincosamide and streptogramin antibiotics in Gram-positive cocci isolated in a Korean hospital. J Antimicrob Chemother. 2002, 49: 489-95. 10.1093/jac/49.3.489.
Sekiguchi J, Hama T, Fujino T, Araake M, Irie A, Saruta K, et al: Detection of the antiseptic- and disinfectant-resistance genes qacA, qacB, and qacC in methicillin-resistant Staphylococcus aureus isolated in a Tokyo hospital. Jpn J Infect Dis. 2004, 57: 288-91.
Weisblum B: Erythromycin resistance by ribosome modification. Antimicrob Agents Chemother. 1995, 39: 577-585.
Murphy E: Nucleotide sequence of ermA, a macrolide-lincosamide streptogramin B determinant in Staphylococcus aureus. J Bacteriol. 1985, 162: 633-640.
Khan SA, Novick RP: Terminal nucleotide sequences of Tn551, a transposon specifying erythromycin resistance in Staphylococcus aureus: homology with Tn3. Plasmid. 1980, 4: 148-154. 10.1016/0147-619X(80)90004-9.
Westh H, Hougaard DM, Vuust J, Rosdahl VT: Erm genes in erythromycin-resistant Staphylococcus aureus and coagulase-negative staphylococci. APMIS. 1995, 103: 225-232. 10.1111/j.1699-0463.1995.tb01099.x.
Fluit ADC, Visser MR, Schmitz F: Molecular detection of antimicrobial resistance. Clin Microbiol Rev. 2001, 14: 836-871. 10.1128/CMR.14.4.836-871.2001.
Ardic N, Ozyurt M, Sareyyupoglu B, Haznedaroglu T: Investigation of erythromycin and tetracycline resistance genes in methicillin-resistant staphylococci. Int J Antimicrob Agents. 2005, 26: 213-218. 10.1016/j.ijantimicag.2005.06.013.
Lyon BR, Skurray RA: Antimicrobial resistance of Staphylococcus aureus: genetic basis. Microbiol Rev. 1987, 51: 88-134.
Schweizer HP: Triclosan: a widely used biocide and its link to antibiotics. FEMS Microbiol Lett. 2001, 202: 1-7. 10.1111/j.1574-6968.2001.tb10772.x.
Chapman S: Disinfectant resistance mechanisms, cross-resistance and co-resistance. Int Biodeterior Biodegrad. 2003, 51: 271-276. 10.1016/S0964-8305(03)00044-1.
Leelaporn A, Paulsen IT, Tennent JM, Littlejohn TG, Skurray RA: Multidrug resistance to antiseptics and disinfectants in coagulase-negative staphylococci. J Med Microbiol. 1994, 40: 214-220. 10.1099/00222615-40-3-214.
Bjorland J, Steinum T, Kvitle B, Waage S, Sunde M, Heir E: Widespread Distribution of Disinfectant Resistance Genes among Staphylococci of Bovine and Caprine Origin in Norway. J Clin Microbiol. 2005, 43: 63-4368.
Heir E, Sundheim G, Holck AL: Resistance to quaternary ammonium compounds in Staphylococcus spp. Isolated from the food industry and nucleotide sequence of the resistance plasmid pST827. J Appl Bacteriol. 1995, 79: 149-156. 10.1111/j.1365-2672.1995.tb00928.x.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
TZ was the primary author of the manuscript, assisted in samples collection, antimicrobial susceptibility, detection of resistance genes and assisted in minimum inhibition concentration determination of BC. BK contributed in minimum inhibition concentration determination, assisted in detection of resistance genes and helped in the writing of the manuscript. HM participated in detection of resistance genes, data acquisition and contributed in writing of the manuscript. AB provided funding, supervised the study, and helped to finalize the manuscript.
All authors read and approved the final manuscript
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Zmantar, T., Kouidhi, B., Miladi, H. et al. Detection of macrolide and disinfectant resistance genes in clinical Staphylococcus aureus and coagulase-negative staphylococci. BMC Res Notes 4, 453 (2011). https://doi.org/10.1186/1756-0500-4-453
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1756-0500-4-453