
Abstract
Background
Pterostilbene, a naturally occurring phenolic compound produced by agronomically important plant genera such as Vitis and Vacciunium, is a phytoalexin exhibiting potent antifungal activity. Additionally, recent studies have demonstrated several important pharmacological properties associated with pterostilbene. Despite this, a systematic study of the effects of pterostilbene on eukaryotic cells at the molecular level has not been previously reported. Thus, the aim of the present study was to identify the cellular pathways affected by pterostilbene by performing transcript profiling studies, employing the model yeast Saccharomyces cerevisiae.
Methods
S. cerevisiae strain S288C was exposed to pterostilbene at the IC50 concentration (70 μM) for one generation (3 h). Transcript profiling experiments were performed on three biological replicate samples using the Affymetrix GeneChip Yeast Genome S98 Array. The data were analyzed using the statistical methods available in the GeneSifter microarray data analysis system. To validate the results, eleven differentially expressed genes were further examined by quantitative real-time RT-PCR, and S. cerevisiae mutant strains with deletions in these genes were analyzed for altered sensitivity to pterostilbene.
Results
Transcript profiling studies revealed that pterostilbene exposure significantly down-regulated the expression of genes involved in methionine metabolism, while the expression of genes involved in mitochondrial functions, drug detoxification, and transcription factor activity were significantly up-regulated. Additional analyses revealed that a large number of genes involved in lipid metabolism were also affected by pterostilbene treatment.
Conclusion
Using transcript profiling, we have identified the cellular pathways targeted by pterostilbene, an analog of resveratrol. The observed response in lipid metabolism genes is consistent with its known hypolipidemic properties, and the induction of mitochondrial genes is consistent with its demonstrated role in apoptosis in human cancer cell lines. Furthermore, our data show that pterostilbene has a significant effect on methionine metabolism, a previously unreported effect for this compound.
Similar content being viewed by others


Background
Pterostilbene is a naturally-occurring phytoalexin identified in several plant species. It belongs to a group of phenolic compounds known as stilbenes, and is found in the heartwood of sandalwood (Pterocarpus santalinus) [1] and P. marsupium [2]. It was also identified in the leaves of Vitis vinifera [3], in infected grape berries of var. Chardonnay and Gamay [4], and in healthy and immature berries of var. Pinot Noir and Gamay [5]. Pterostilbene has also been found in berries of some Vacciunium species [5]. Pterostilbene, one of the most extensively studied secondary metabolites found in grapes and wine, is a dimethylether analog of resveratrol that is well known for its hypolipidemic activity. A considerable amount of research effort has been expended to address the biochemical and physiological effects of pterostilbene in animal and microbial systems. For example, the antioxidative activity of pterostilbene was first demonstrated in vitro by its inhibition of methyl linoleate oxidation [6]. Pterostilbene was reported to scavenge 1,1-diphenyl-2-picryl-hydrazyl (DPPH) free radicals and to inhibit the oxidation of citronellal, and lipid peroxidation in rat liver microsomes and in cultured human fibroblasts [7]. Pterostilbene isolated from Anogeissus acuminata (Family Combretaceae) is cytotoxic against a number of cancer cell lines, including human breast cancer and murine lymphoid neoplasma cells [8, 9]. More recently, it has been demonstrated that pterostilbene can reduce cholesterol levels in vivo in hamsters through the activation of the peroxisome proliferator-activated receptor α (PPARα) [10]. Pterostilbene has been reported to reduce glucose and increase plasma insulin levels significantly in normal and diabetic rats [11]. Furthermore, pterostilbene has been shown to be cancer-chemopreventive [8, 12] and anti-inflammatory [13].
Investigation of the pathogen-host interactions of Vitis vinifera has led to the hypothesis that resistance is not due to preformed physical or chemical factors, but rather to an active defense mechanism that is triggered by the pathogen, of which stress metabolites including resveratrol, α-viniferin and ε-viniferin are an important component [14]. Pterostilbene, produced in leaf tissues by various species of the Vitaceae family following fungal infection, proved to have more potent antifungal activity than resveratrol (reviewed in [3, 15, 16]). However, the mechanism by which pterostilbene inhibits fungi is not well understood. Results from early studies suggested that the biological activities of the compound mainly involved effects on the plasma membrane [5, 17], and destruction of ribosomes, endoplasmic reticulum, and mitochondrial membranes [17]. Further information on its precise mechanism of action would be useful not only for its potential development as a drug, but also in understanding its ecological significance to producing plant species. In the present study, using transcript profiling analysis, we monitored the gene expression profile of yeast cells treated with pterostilbene in an effort to identify the molecular pathways affected by this compound.
Methods
Yeast strains and media
S. cerevisiae S288C (MATα, SUC2, mal, mel, gal2, CUP1, flo1, flo8-1), obtained from ATCC (Manassas, VA), was used in the microarray experiments. The deletion strains and the isogenic wild type strain (BY4742) were obtained from Open Biosystems (Huntsville, AL). Synthetic dextrose (SD) medium, containing 0.67% (w/v) yeast nitrogen base without amino acids and 2% (w/v) dextrose, was used to grow the wild type S288C strain. Standard Yeast Peptone Dextrose (YPD) medium (1% yeast extract, 2% peptone, 2% dextrose) was used to grow the deletion strains and their respective isogenic wild type control. In all cases, the medium was buffered with 0.165 M 3- [N-Morpholino]propanesulfonic acid (MOPS) (Sigma-Aldrich Corp.) and the pH was adjusted to 7.0 with NaOH.
IC50determination and yeast cultures for microarray studies
IC50 value (concentration of inhibitor required for 50 percent inhibition of growth) was first determined in a conventional microplate assay. The obtained IC50 was then further confirmed in 50-ml large-scale cultures as previously described in order to closely mimic the microarray experimental conditions [18]. For microarray experiments, a single colony of S. cerevisiae was inoculated into 25 ml of SD medium and grown overnight at 30°C with shaking at 200 rpm. The culture was used to inoculate 50-ml of SD medium to an A600 of 0.1. Three independent 50-ml cultures were grown for pterostilbene treatments. In parallel, another set of three independent cultures were grown for solvent treatments which served as the untreated controls. When the cultures reached an A600 of 0.2, pterostilbene was added at 70 μM concentration from a stock solution of 0.5 M dissolved in dimethyl sulfoxide (DMSO). In the control cultures, 0.25% of DMSO was simultaneously added. The cultures were allowed to grow until an A600 of 0.5 was reached [18]. Cells were harvested by centrifugation at 500 × g in a Sorvall centrifuge using a SH-3000 rotor. The cells were flash-frozen in liquid nitrogen and stored at -80°C until use.
RNA preparation and hybridization to Affymetrix GeneChips
Total RNA was isolated using the Qiagen RNeasy® Midi-kit (Qiagen, Inc., Valencia, CA) with modifications as previously described [18]. RNA was processed for target preparation using the GeneChip One-Cycle Target Labeling and Control Reagents kit (Affymetrix, Santa Clara, CA), according to the manufacturer's protocol. Microarray hybridization was performed using the Affymetrix GeneChip Yeast Genome S98 Array using protocols described by Affymetrix (Santa Clara, CA). All microarray data are deposited at the Gene Expression Omnibus as GSE10554.
Microarray data analysis
Normalized data from the Affymetrix software were analyzed using the GeneSifter microarray data analysis system (VizX Labs LLC, Seattle, WA; [19]). This program identifies differentially expressed genes and establishes the biological significance based on Gene Ontology (GO) classification [20, 21] into biological process, molecular function and cellular component. The GeneSifter program also produces z-score reports. A z-score is a statistical rating of gene ontologies and indicates whether each GO term occurs more or less frequently than expected. The differential expression of genes was calculated by averaging the normalized triplicate samples and running a pairwise analysis [22, 23]. Statistical significance was determined using Student t-test (two tail, unpaired) with correction factor for false discovery rate (Benjamini and Hochberg) included [24]. Gene lists generated using GeneSifter were further analyzed using the Saccharomyces Genome Database [25].
Quantitative real-time RT-PCR assay
To confirm the results from microarray experiments, real-time quantitative RT-PCR was carried out using the same RNA preparations that were used in the microarray experiments. Total RNA was treated with DNase I and the RT-PCR reaction was performed as described [18]. The sequences of the primers for each gene selected for the assays are listed in Table 1. Additionally, linear regression analysis as well as an F-Test to assess goodness-of-fit was performed on log2-transformed expression ratios (pterostilbene treated:untreated cells) obtained from real-time RT-PCR and microarray experiments (see Additional file 1: Correlation between expression ratios obtained from quantitative real-time RT-PCR and microarray experiments) using the Microsoft Excel Analysis ToolPak add-in module, and a statistically-significant (p < 0.05) correlation between the two methods was indicated.
Sensitivity assays with yeast mutants
Cells were grown overnight in YPD broth (pH 7.0, MOPS buffered) until the optical density of 3.0 at A600 was reached. For mutants, G418 at the concentration of 200 μg/ml was added to the broth. Seven serial 1:5 dilutions of the overnight cultures were prepared in appropriate medium and plated on YPD agar containing either 110 μM of pterostilbene or 1% DMSO as control. The plates were photographed after incubating for 2 days at 30°C.
Results
IC50 determination in S. cerevisiae
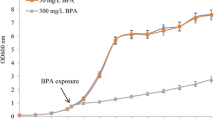
Pterostilbene is known to possess strong antifungal activity [16]. In this study, in order to investigate how pterostilbene functions in fungal cells, we determined the consequences of exposure to this compound by monitoring the whole-genome transcriptional response in the model yeast Saccharomyces cerevisiae. We made use of S. cerevisiae since it has been extensively used for elucidating the molecular targets of antifungal and therapeutic compounds (reviewed in [26, 27]). We first determined the IC50 value for pterostilbene in S. cerevisiae cells using a combination of microplate-based and large-scale culture assays as previously described [18]. The IC50 was determined to be 70 μM based on the average value from four independent experiments (Figure 1), and this concentration was used for subsequent microarray studies.

Determination of IC50 concentration for pterostilbene in yeast cells. Results are shown from a microtiter assay (experiment 1) and three independent assays performed in large scale cultures (experiment 2–4) as described in "Methods".
Categories of genes responding to pterostilbene
Our results demonstrate that pterostilbene significantly (p < 0.05) affected the expression of over a thousand genes under our experimental conditions, with the up- and down-regulation of 1007 and 182 genes, respectively (see Additional file 2: List of all significant pterostilbene-responding genes). The most immediate observation is the very high fraction of up-regulated genes, indicating many different biological processes are affected by pterostilbene (Table 2). Of these, 44 genes were up-regulated more than 10-fold. The majority of the genes showed fold-changes ranging between 2.0 to 5.0, while approximately 200 genes showed fold-changes of 5.0 to10.0 (Table 2).
To explore the biological significance of the observed transcriptome response to pterostilbene, a z-score report was generated (Table 3) using the GeneSifter software [19]. A positive z-score indicates that more genes than expected by chance alone fulfill the criterion in a specific ontology term, while a negative z-score indicates that the term occurs less frequently than expected [22]. Pterostilbene affected the expression of genes that were associated with a diverse array of biological, molecular and cellular ontologies. As shown in Table 3, the most significantly enriched ontologies identified for the up-regulated genes included "response to drug," "regulation of transcription from PolII promoter," "mitochondrion organization and biogenesis," and "protein folding," with z-up scores of 5.05, 2.4, 2.12, and 4.24 respectively (Table 3, under Biological Process). Related processes were also significantly over-represented under Molecular Function, as represented by the following categories: "ABC transporter activity," "transcription factor activity," "ATPase activity," "peptidase activity," and "chaperone activity," with z-scores of 3.02, 4.67, 2.24, 2.12, and 3.56 respectively (Table 3, under Molecular Function). Among the down-regulated genes, terms associated with the biological process "amino acid and derivative metabolism" (specifically, the subcategory of "methionine metabolism"), and the cellular component "cell wall" were over-represented with z-scores of 5.08 and 8.22, respectively.
As mentioned, pterostilbene is involved in fatty acid and lipid catabolism through activation of the peroxisome proliferator-activated receptor α (PPARα) [10]. With respect to the gene ontology analysis (Table 3), the effect on lipid metabolism by pterostilbene was observed to be less statistically significant, however further examination revealed several important genes associated with lipid metabolism within the microarray data (Table 4). These included several genes involved in fatty acid breakdown including FOX2 (multifunctional beta-oxidation protein), ECI1 (peroxisomal Δ3, Δ2-enoyl-CoA isomerase), and OAF1 (oleate-activated transcription factor). In addition, many genes involved in the biosynthesis of fatty acids, phospholipids and sphingolipids were up-regulated by pterostilbene treatment, as well as 8 genes involved in the regulation of lipid metabolism. The latter include OAF1 (involved in regulation of beta-oxidation of fatty acids) [28, 29], UPC2 (involved in regulation of sterol biosynthesis and transport) [30], and INO4 (involved in regulation of phospholipid metabolism) [31]. Furthermore, two additional genes, MGA2 and SPT23, encoding endoplasmic reticulum membrane proteins, Spt23p and Mga2p, respectively, were also up-regulated. Spt23p and Mga2p regulate expression of OLE1, a gene encoding Ole1p, an intrinsic membrane-bound -9 fatty acid desaturase required for the synthesis of oleic acid in yeast cells [32].
Pterostilbene exposure also resulted in a significant up-regulation of genes associated with the ontology term "response to drug" (z-score of 5.05 – Table 3; genes listed in Table 5). Three of these genes encode transcription factors, including PDR3 involved in the regulation of the pleiotropic drug resistance (PDR) response in yeast [33], YAP1, encoding a basic leucine zipper transcription factor required for oxidative stress tolerance [34], and YRR1, encoding a Zn2-Cys6 zinc-finger transcription factor that activates genes involved in multidrug resistance [35]. Notably, the significantly induced genes in this category were transporters, especially the ABC transporters whose function is known to be involved in multidrug resistance against a broad range of antifungal agents [36]. It should be also noted that the AZR1 gene, which encodes a plasma membrane protein required for adaptation to acetic acid and resistance to azole drugs such as ketoconazole and fluconazole in yeast [37], was induced by more than 73-fold (Table 5).
Including the transcription factors discussed above involved in the regulation of lipid metabolism and pleiotropic drug resistance, in total more than 30 transcription factors were up-regulated in response to pterostilbene (see Additional file 3: List of pterostilbene-responding genes in the "cell wall," "transcription factor activity," and "mitochondrion" categories). Of particular interest are genes encoding transcription factors involved in mitochondrial function (RTG1 and RTG2), cell wall-related functions (RLM1), and methionine metabolism (MET4 and MET31), considering that these functional categories were over-represented in the gene ontology analysis (Table 3). Thus, the genes present in these functional categories include transcription factors as well as their cognate targets, suggesting the modulation of these processes, at least in part, via altered levels of these specific transcription factors in response to pterostilbene exposure.
In the Cellular Component-based ontology analysis, genes required for mitochondrial and cell wall-related functions were also over-represented (Table 3). The cell wall category had a z-score of 8.22 (Table 3), and most of the genes assigned within this category were down-regulated by pterostilbene treatment (Table 3). The majority of these genes are involved in mating responses [e.g., FIG1, FIG2, SAG1, AGA1, MF(ALPHA)1, and MF(ALPHA)2, see Additional file 3: op. cit.], which is also in agreement with the over-representation of the gene ontology term "reproduction" in the Biological Process category (z-score of 3.01, Table 3). Additional down-regulated cell wall-related genes included chitin synthase 2(CHS2), cell wall mannoproteins (TIP1, CWP1) and a major exo-1,3-beta-glucanase of the cell wall (EXG1) (see Additional file 3: op. cit.). These genes are responsible for the biosynthesis of three major constituents (chitin, mannoproteins and glucan) found in yeast cell walls [38]. The gene BAG7 (Rho GTPase activating protein), however, was induced by ~137-fold in pterostilbene treated cells (see Additional file 3: op. cit.). It has been reported that Bag7p plays a role in the control of cell wall synthesis and is involved in regulating key components of the cell wall stress-response pathway [39].
With respect to mitochondrial genes, it is interesting to note that more than 100 genes involved in diverse mitochondrial functions were up-regulated by pterostilbene treatment (Table 3, and Additional file 3: op. cit.). These genes play important roles in respiration, electron transport, mitochondrial protein targeting, and mitochondrial protein synthesis (see Additional file 3: op. cit.).
Of particular interest was the finding that a group of genes involved in sulfur metabolism was significantly over-represented, generating a z-score of 10.5 for the category "sulfur amino acid metabolism," (the highest score obtained in the present study). Within this category, a subset of genes assigned to the ontology term "methionine metabolism" (z-score of 11.78) were down-regulated upon exposure to pterostilbene (Table 3). Of the 21 genes assigned to this ontology term on the array, eleven were differentially regulated. As shown in Figure 2 and Table 6, all responsive genes involved in sulfur-containing amino acid biosynthesis were down-regulated, with the exception of MET4 (transcriptional activator) and MET31 (transcriptional regulator), which were up-regulated (Table 6).

Summary of gene expression responses to pterostilbene in the sulfur metabolism pathway. Methionine biosynthetic pathway and corresponding genes are based on Thorsen et al. [66] and the Saccharomyces Genome Database [25]. Genes shown in boldface in the pathway are those affected by pterostilbene. The up- and down-regulated genes were indicated by arrows, (↑) and (↓).
Validation of microarray data by quantitative real-time RT-PCR
For further verification of the transcript profiling results, selected genes were analyzed by quantitative real time RT-PCR with cDNAs prepared from the identical RNA samples used for microarray target preparations. For these analyses, eleven genes associated with gene ontology categories discussed above were selected, including genes involved in lipid metabolism (UPC2, OAF1, RSB1 and INO4), sulfur metabolism (MET3), response to drug (AZR1 and PDR3), mitochondrial functions (RTG1 and RTG3) and cell wall-related functions (RLM1 and BAG7). The results (Figure 3) are in agreement with the microarray experiments, with ten genes showing up-regulation and one gene (MET3) showing down-regulation in response to pterostilbene treatment. A few discrepancies in fold-change values between real-time RT-PCR and microarray analyses could be attributed to technical differences between the two methods. For example, the genes RSB1, AZR1, and BAG7 showed a higher level of induction in the microarray data compared to the real-time RT-PCR data. This discrepancy could be attributed to the fact that the Affymetrix software assigned an "absent" call to these genes in the untreated sample – thus the signal values assigned were artifactually low in the untreated sample compared to the treated sample, making the fold-change values very high [18]. Other discrepancies could be attributed to the greater dynamic range of real-time RT-PCR compared to microarray analysis.

Quantitative real-time RT-PCR analysis of genes identified as differentially expressed by microarray experiments. Assays were performed in triplicate as described previously [18]. Data were normalized to an internal control (18S rRNA) and the ΔΔCT method was used to obtain the relative expression level for each gene. Data are shown as mean ± standard deviation (SD). "Ctrl" represents samples treated with solvent (0.25% DMSO) alone.
Sensitivity of yeast mutants to pterostilbene
In order to further investigate the relationship between the gene expression responses to pterostilbene and their phenotypic consequences in yeast mutants, we analyzed a number of yeast mutants with deletions in selected genes from each of the categories discussed above for their sensitivity to pterostilbene. The same genes selected for real-time RT-PCR analysis were also chosen for this study, and these included genes involved in lipid metabolism (UPC2, OAF1, RSB1 and INO4), sulfur metabolism (MET3), response to drug (AZR1 and PDR3), mitochondrial functions (RTG1 and RTG3) and cell wall-related functions (RLM1 and BAG7). As shown in Figure 4, the loss of Pdr3p (mutant pdr3Δ) resulted in increased sensitivity to pterostilbene, while the loss of Upc2p, Oaf1p, Bsr1p, Ino4, Met3p, Azr1p, Rlm1p or Bag7p resulted in neither increased nor decreased sensitivity to pterostilbene under the experimental conditions used. Surprisingly, the loss of Rtg1p or Rtg3p resulted in slightly increased resistance to the compound.

Growth of selected yeast mutant strains in the presence of pterostilbene. A parent strain BY4742 and 11 mutants (upc2Δ, oaf1Δ, rsb1Δ, ino4Δ, met3Δ, azr1Δ, pdr3Δ, rtg1Δ, rtg3Δ, rlm1Δ, and bag7Δ) were analyzed. Serial dilutions of overnight cultures were spotted onto solid YPD (pH 7.0, MOPS buffered) agar plates containing either 1% DMSO or 110 μM pterostilbene, and the plates were photographed after incubation for 2 days at 30°C.
Discussion
Stilbenes, such as resveratrol and pterostilbene, represent biologically active secondary metabolites of plants known to have diverse pharmacological properties, which include cancer-chemopreventive activity. They are also considered to be phytoalexins due to their potent antifungal activities [14–16]. To investigate the molecular pathways affected by pterostilbene in eukaryotes, we conducted DNA microarray analysis using the model yeast S. cerevisiae, and identified large-scale transcriptome alterations associated with pterostilbene exposure in this organism. The responsive genes are associated with a diversity of biological, molecular and cellular processes, suggesting that pterostilbene exerts wide-ranging effects on the physiological and biochemical processes of eukaryotic cells (Tables 2 and 3).
Pterostilbene affected the expression of genes involved in sulfur metabolism and/or sulfur assimilation, associated with the highest z-score in this study (Tables 3 and 6). Within this category, six genes (ECM17, MET3, MET14, MET16, MET10, MET6) encoding enzymes involved in methionine biosynthesis were repressed (Figure 2, Table 6). Methionine metabolism plays an important role in many aspects of cellular physiology. Its derivative S-adenosyl methionine (SAM) serves as a methyl donor in many biosynthetic processes [40]. Metabolism of sulfur-containing amino acids like methionine has been linked to cell cycle progression, and perturbations of these processes lead to diverse cellular anomalies [41, 42]. The down-regulation of genes involved in methionine biosynthesis by pterostilbene may cause cellular stress by not only decreasing methionine levels, but also by compromising the supply of donor methyl groups required for methylation reactions in various biosynthetic pathways. Our results suggest that one of the molecular effects of pterostilbene might involve the disruption of methionine biosynthesis, an observation that has not been previously reported for this compound.
The observed down-regulation of genes involved in methionine metabolism in response to pterostilbene treatment (Figure 2, Table 6) is perhaps surprising given that several genes in this pathway are actually up-regulated during conditions of oxidative stress and amino acid starvation [43, 44]. However, a transcriptional profiling study conducted in yeast cells exposed to the herbicide sulfometuron methyl (SM), which inhibits branched-chain amino acid biosynthesis, indicated that exposure to SM resulted in the down-regulation of several genes involved in methionine metabolism, including MET3, MET6, MET14, SAM1, and SAM2 [45]. One mechanism proposed for this down-regulation was the potentially reduced levels of ATP in SM-treated cells, given that methionine biosynthesis and the production of SAM are ATP-requiring processes. Interestingly, in the present work pterostilbene treatment altered the expression of more than 100 mitochondrial genes (Additional file 3: op. cit.), suggesting large-scale perturbations in mitochondrial function which would eventually lead to ATP deficiency. A second potential mechanism emerged from queries using MET1, MET3, MET6, MET10, MET13, MET14, and MET16 against the Serial Pattern of Expression Levels Locator (SPELL) database [46], which revealed that all of the corresponding transcripts are down-regulated in response to osmotic stress [47]. Given the significant effects pterostilbene exposure is likely to have on lipid metabolism (Table 4), it is possible that membrane integrity could be compromised leading to an osmotic imbalance in yeast cells. Consistent with this notion, genes involved in osmotic stress response regulation such as GRE1, GRE2, SSK1, PPZ1, and STE11 were induced in pterostilbene-treated cells (Additional file 3: op. cit.).
Of further significance, the present results show that pterostilbene up-regulated OAF1, which encodes a transcription factor that regulates the expression of genes involved in the beta-oxidation of fatty acids in peroxisomes in yeast cells [28]. In addition, genes encoding enzymes required for fatty acid β-oxidation were also up-regulated by pterostilbene. It has been previously shown that pterostilbene lowers lipid/lipoprotein levels in hypercholestrolemic hamsters through activation of the peroxisome proliferator-activated receptor α (PPARα) [10]. PPARα is involved in fatty acid and lipid metabolism, through the activation of genes involved in fatty acid β-oxidation in the liver, heart, kidney and skeletal muscles [10, 48, 49]. Thus, the up-regulation of genes involved in fatty acid beta-oxidation by pterostilbene in the present report is consistent with previous observations of its effects on mammalian cells. In addition, pterostilbene also up-regulated several genes involved in sterol, phospholipid and sphingolipid metabolism, including genes involved in the regulation of lipid metabolism. Taken together, these results suggest that lipid metabolism is likely to be an important molecular pathway that is affected by pterostilbene.
Transcript levels of a number of genes involved in the pleiotropic/multiple drug resistance response were also found to increase dramatically following pterostilbene exposure (Table 5). These genes include ABC transporters, multidrug resistance transcription factors, and other drug-responsive genes. The plasma membrane-associated efflux pumps Pdr5p and Snq2p are under the genetic control of the transcription factors Pdr1p and Pdr3p [50]. In yeast cells exposed to pterostilbene, the transcripts of PDR3, PDR5 and SNQ2 increased 9.5-, 6.1- and 8.2-fold, respectively (Table 5). In addition, a yeast mutant with a deletion in the PDR3 gene showed strong hypersensitivity to pterostilbene, confirming the importance of this gene in conferring resistance to this compound (Figure 4). Notably, the ABC transporter YOR1, involved in mediating the export of many different organic anions including oligomycin, was also up-regulated 10.3-fold in pterostilbene-treated cells. Moreover, the expression of a plasma membrane transporter, encoded by AZR1, which is involved in the resistance to azole drugs, dramatically increased by 73.4-fold following exposure. Taken together, the strong up-regulation of this set of transporters suggests that drug efflux is one of the major mechanisms involved in the cellular detoxification of pterostilbene in yeast cells.
As mentioned above, an additional outcome from the present work is the observation of the up-regulation of a large number of genes involved in diverse mitochondrial functions, including mitochondrial respiration, mitochondrial protein synthesis and mitochondrial protein targeting (associated with a z-score of 6.1 – Tables 3, and Additional file 3: op. cit.). Furthermore, RTG1 and RTG3, genes encoding transcription factors that play an important role in regulating the communication between the mitochondria and the nucleus in yeast cells, were up-regulated by pterostilbene. Mitochondria are vital for energy production, and their disruption in humans has been implicated in aging, diabetes, heart disease, and various neurodegenerative disorders [51]. Recently, one of the mechanisms by which pterostilbene induces apoptosis in human gastric carcinoma cells was found to involve the activation of the caspase cascade via the mitochondrial pathway [52]. Thus, the induction of mitochondrial genes by pterostilbene in the present study is apparently consistent with its effects on this organelle in human cancer cell lines.
Of the eleven mutants tested carrying deletions in selected genes from pterostilbene-responsive pathways, only the pdr3Δ mutant showed hypersensitivity to pterostilbene (Figure 4). Lack of correlation between transcriptional profiling and deletion mutant analyses has been reported in the literature by investigators using both selected individual mutants as well as whole-genome mutant populations [53–57]. Such discrepancies could be attributable to factors such as post-transcriptional regulation of target genes [56], functional redundancy [53], as well as the specific nature of the inhibitor tested (e.g., genes required for survival in the presence of the inhibitor could be distinct from those which are transcriptionally responsive; [55]). Nevertheless, the hypersensitivity to pterostilbene exhibited by the pdr3Δ mutant in the present study, coupled with elevated levels of PDR3 as well as other multidrug resistance-related transcripts (Figure 4, Table 5), confirms the importance of efflux-mediated detoxification pathways in the cellular response to this compound. Further mechanistic insights could potentially be gained by extending these analyses to whole-genome mutant collections of S. cerevisiae generated by The Saccharomyces Genome Deletion Project [57], which have been successfully used by others for the identification of molecular targets of various clinically relevant drugs, as well as for the investigation of interactions between the cellular pathways affected [58–61]. Such studies, coupled with transcriptional profiling experiments, could provide further corroborative evidence indicating the relevance of specific genes to the mechanism of action of pterostilbene.
The present study provides an overview of the major transcriptional responses to pterostilbene using the model eukaryote S. cerevisiae. While transcriptional profiling was employed as the principal tool to investigate the molecular effects of pterostilbene, an examination of the response of the yeast proteome would further refine our understanding of pterostilbene-induced gene expression changes, given that transcript and protein levels do not correlate in all cases [62–65]. Additionally, the transcriptional profiling results presented in the current work rely on a single drug concentration (IC50) and time point (one doubling), thus it is likely that many of the transcriptional responses observed represent indirect consequences of drug exposure. An expansion of the current data set to include multiple time points and drug concentrations could therefore prove invaluable in distinguishing between the primary and secondary effects of pterostilbene, such as mechanism-related vs. adaptation-related cellular responses. Nevertheless, the present work yields important clues and provides a foundation for further studies directed towards elucidating the precise mechanism of action of pterostilbene.
Conclusion
We have identified the molecular pathways affected by pterostilbene, and our results show that pterostilbene affects the expression of a diverse group of genes in yeast cells. Using Gene Ontology-based analysis, the most significant effects were observed in genes involved in "methionine metabolism," "response to drug," "transcription factor activity," and "mitochondrion" functions. Additional analyses indicated that many genes involved in lipid metabolism were also affected. The observed response of lipid metabolism genes is in agreement with the known hypolipidemic properties of pterostilbene mediated through the activation of PPARα. The induction of a large number of mitochondrial genes by pterostilbene is consistent with its previously-demonstrated role in apoptosis in human cancer cells. Our data also show that pterostilbene has a significant effect on methionine metabolism, perhaps resulting in the depletion of methionine by the inhibition of methionine biosynthesis. The effect of pterostilbene on methionine metabolism has not been previously observed and merits further investigation.
References
Sehadri R: Polyphenols of Pterocarpus and Dalbergia woods. Phytochemistry. 1972, 11: 881-898. 10.1016/S0031-9422(00)88430-7.
Maurya R, Ray AB, Duah FK, Slatkin DJ, Schiff PL: Constituents of Pterocarpus marsupium. Journal of Natural Products. 1984, 47: 179-181. 10.1021/np50031a029.
Langcake P, Cornford CA, Pryce RJ: Identification of pterostilbene as a phytoalexin from Vitis vinifera leaves. Phytochemistry. 1979, 18: 1025-1027. 10.1016/S0031-9422(00)91470-5.
Adrian M, Jeandet P, Douillet-Breuil AC, Tesson L, Bessis R: Stilbene content of mature Vitis vinifera berries in response to UV-C elicitation. J Agric Food Chem. 2000, 48: 6103-6105. 10.1021/jf0009910.
Pezet R, Pont V: Identification of pterostilbene in grape berries of Vitis vinifera. Plant Physiol Biochem (Paris). 1988, 26: 603-607.
Charvet-Faury S, Derbesy M, Cochini F, Derbesy F: andalwood extract (Pterocarpus santalinus): anti-oxidant and anti-UV effects study. Riv Ital EPPOS. 1998, spec no: 435-458.
Stivala LA, Savio M, Carafoli F, Perucca P, Bianchi L, Maga G, Forti L, Pagnoni UM, Albini A, Prosperi E, Vannini V: Specific structural determinants are responsible for the antioxidant activity and the cell cycle effects of resveratrol. J Biol Chem. 2001, 276: a22586-22594. 10.1074/jbc.M101846200.
Rimando AM, Cuendet M, Desmarchelier C, Mehta RG, Pezzuto JM, Duke SO: Cancer chemopreventive and antioxidant activities of pterostilbene, a naturally occurring analogue of resveratrol. J Agric Food Chem. 2002, 50: 3453-3457. 10.1021/jf0116855.
Rimando AM, Pezzuto JM, Farnsworth NR, Santisuk T, Reutrakul V: Revision of the NMR assignments of pterostilbene and of dihydrodehydrodiconiferyl alcohol: Cytotoxic constituents from Anogeissus acuminata. Nat Prod Lett. 1994, 4: 267-272.
Rimando AM, Nagmani R, Feller DR, Yokoyama W: Pterostilbene, a new agonist for the peroxisome proliferator-activated receptor alpha-isoform, lowers plasma lipoproteins and cholesterol in hypercholesterolemic hamsters. J Agric Food Chem. 2005, 53: 3403-3407. 10.1021/jf0580364.
Pari L, Satheesh MA: Effect of pterostilbene on hepatic key enzymes of glucose metabolism in streptozotocin- and nicotinamide-induced diabetic rats. Life Sci. 2006, 79: 641-645. 10.1016/j.lfs.2006.02.036.
Suh N, Paul S, Hao X, Simi B, Xiao H, Rimando AM, Reddy BS: Pterostilbene, an active constituent of blueberries, suppresses aberrant crypt foci formation in the azoxymethane-induced colon carcinogenesis model in rats. Clin Cancer Res. 2007, 13: 350-355. 10.1158/1078-0432.CCR-06-1528.
Hougee S, Faber J, Sanders A, de Jong RB, van den Berg WB, Garssen J, Hoijer MA, Smit HF: Selective COX-2 inhibition by a Pterocarpus marsupium extract characterized by pterostilbene, and its activity in healthy human volunteers. Planta Med. 2005, 71: 387-392. 10.1055/s-2005-864130.
Langcake P: Disease resistance of Vitis spp. and the production of the stress metabolites resveratrol, e-viniferin, a-viniferin and pterostilbene. Physiological Plant Pathology. 1981, 18: 312-226.
Hart JH: Role of Phytostilbenes in Decay and Disease Resistance. Ann Rev Phytopathol. 1981, 19: 437-458. 10.1146/annurev.py.19.090181.002253.
Alessandro M, Marco SD, Osti F, Cesari A: Bioassays on the activity of resveratrol, pterostilbene and phosphorous acid towards fungal associated with esca of grapevine. Phytopathol Mediterr. 2000, 39: 357-365.
Pezet R, Pont V: Ultrastructural observations of pterostilbene funcgitoxicity in dormant conidia of Botrytis cinera Pers. J Phytopathology. 1990, 129: 19-30.
Agarwal AK, Rogers PD, Baerson SR, Jacob MR, Barker KS, Cleary JD, Walker LA, Nagle DG, Clark AM: Genome-wide expression profiling of the response to polyene, pyrimidine, azole, and echinocandin antifungal agents in Saccharomyces cerevisiae. J Biol Chem. 2003, 278: 34998-35015. 10.1074/jbc.M306291200.
GeneSifter microarray data analysis system. [http://www.genesifter.net/web/]
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G: Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000, 25: 25-29. 10.1038/75556.
Gene Ontology. [http://www.geneontology.org/GO.doc.html]
Cheadle C, Vawter MP, Freed WJ, Becker KG: Analysis of microarray data using Z score transformation. J Mol Diagn. 2003, 5: 73-81.
Doniger SW, Salomonis N, Dahlquist KD, Vranizan K, Lawlor SC, Conklin BR: MAPPFinder: using Gene Ontology and GenMAPP to create a global gene-expression profile from microarray data. Genome Biol. 2003, 4: R7-10.1186/gb-2003-4-1-r7.
Reiner A, Yekutieli D, Benjamini Y: Identifying differentially expressed genes using false discovery rate controlling procedures. Bioinformatics. 2003, 19: 368-375. 10.1093/bioinformatics/btf877.
Saccharomyces Genome Database. [http://www.yeastgenome.org/]
Parsons AB, Geyer R, Hughes TR, Boone C: Yeast genomics and proteomics in drug discovery and target validation. Prog Cell Cycle Res. 2003, 5: 159-166.
Sturgeon CM, Kemmer D, Anderson HJ, Roberge M: Yeast as a tool to uncover the cellular targets of drugs. Biotechnol J. 2006, 1: 289-298. 10.1002/biot.200500039.
Karpichev IV, Small GM: Global regulatory functions of Oaf1p and Pip2p (Oaf2p), transcription factors that regulate genes encoding peroxisomal proteins in Saccharomyces cerevisiae. Mol Cell Biol. 1998, 18: 6560-6570.
Rottensteiner H, Kal AJ, Hamilton B, Ruis H, Tabak HF: A heterodimer of the Zn2Cys6 transcription factors Pip2p and Oaf1p controls induction of genes encoding peroxisomal proteins in Saccharomyces cerevisiae. Eur J Biochem. 1997, 247: 776-783. 10.1111/j.1432-1033.1997.00776.x.
Wilcox LJ, Balderes DA, Wharton B, Tinkelenberg AH, Rao G, Sturley SL: Transcriptional profiling identifies two members of the ATP-binding cassette transporter superfamily required for sterol uptake in yeast. J Biol Chem. 2002, 277: 32466-32472. 10.1074/jbc.M204707200.
Ambroziak J, Henry SA: INO2 and INO4 gene products, positive regulators of phospholipid biosynthesis in Saccharomyces cerevisiae, form a complex that binds to the INO1 promoter. J Biol Chem. 1994, 269: 15344-15349.
Chellappa R, Kandasamy P, Oh CS, Jiang Y, Vemula M, Martin CE: The membrane proteins, Spt23p and Mga2p, play distinct roles in the activation of Saccharomyces cerevisiae OLE1 gene expression. Fatty acid-mediated regulation of Mga2p activity is independent of its proteolytic processing into a soluble transcription activator. J Biol Chem. 2001, 276: 43548-43556. 10.1074/jbc.M107845200.
Balzi E, Goffeau A: Yeast multidrug resistance: the PDR network. J Bioenerg Biomembr. 1995, 27: 71-76. 10.1007/BF02110333.
Temple MD, Perrone GG, Dawes IW: Complex cellular responses to reactive oxygen species. Trends Cell Biol. 2005, 15: 319-326. 10.1016/j.tcb.2005.04.003.
Toone WM, Jones N: AP-1 transcription factors in yeast. Curr Opin Genet Dev. 1999, 9: 55-61. 10.1016/S0959-437X(99)80008-2.
Rogers B, Decottignies A, Kolaczkowski M, Carvajal E, Balzi E, Goffeau A: The pleitropic drug ABC transporters from Saccharomyces cerevisiae. J Mol Microbiol Biotechnol. 2001, 3: 207-214.
Tenreiro S, Rosa PC, Viegas CA, Sa-Correia I: Expression of the AZR1 gene (ORF YGR224w), encoding a plasma membrane transporter of the major facilitator superfamily, is required for adaptation to acetic acid and resistance to azoles in Saccharomyces cerevisiae. Yeast. 2000, 16: 1469-1481. 10.1002/1097-0061(200012)16:16<1469::AID-YEA640>3.0.CO;2-A.
Lipke PN, Ovalle R: Cell wall architecture in yeast: new structure and new challenges. J Bacteriol. 1998, 180: 3735-3740.
Schmidt A, Schmelzle T, Hall MN: The RHO1-GAPs SAC7, BEM2 and BAG7 control distinct RHO1 functions in Saccharomyces cerevisiae. Mol Microbiol. 2002, 45: 1433-1441. 10.1046/j.1365-2958.2002.03110.x.
Thomas D, Surdin-Kerjan Y: Metabolism of sulfur amino acids in Saccharomyces cerevisiae. Microbiol Mol Biol Rev. 1997, 61: 503-532.
Patton EE, Peyraud C, Rouillon A, Surdin-Kerjan Y, Tyers M, Thomas D: SCF(Met30)-mediated control of the transcriptional activator Met4 is required for the G(1)-S transition. Embo J. 2000, 19: 1613-1624. 10.1093/emboj/19.7.1613.
Kaiser P, Flick K, Wittenberg C, Reed SI: Regulation of transcription by ubiquitination without proteolysis: Cdc34/SCF(Met30)-mediated inactivation of the transcription factor Met4. Cell. 2000, 102: 303-314. 10.1016/S0092-8674(00)00036-2.
Iwahashi Y, Hosoda H, Park JH, Lee JH, Suzuki Y, Kitagawa E, Murata SM, Jwa NS, Gu MB, Iwahashi H: Mechanisms of patulin toxicity under conditions that inhibit yeast growth. J Agric Food Chem. 2006, 54: 1936-1942. 10.1021/jf052264g.
Natarajan K, Meyer MR, Jackson BM, Slade D, Roberts C, Hinnebusch AG, Marton MJ: Transcriptional profiling shows that Gcn4p is a master regulator of gene expression during amino acid starvation in yeast. Mol Cell Biol. 2001, 21: 4347-4368. 10.1128/MCB.21.13.4347-4368.2001.
Jia MH, Larossa RA, Lee JM, Rafalski A, Derose E, Gonye G, Xue Z: Global expression profiling of yeast treated with an inhibitor of amino acid biosynthesis, sulfometuron methyl. Physiol Genomics. 2000, 3: 83-92.
Hibbs MA, Hess DC, Myers CL, Huttenhower C, Li K, Troyanskaya OG: Exploring the functional landscape of gene expression: directed search of large microarray compendia. Bioinformatics. 2007, 23: 2692-2699. 10.1093/bioinformatics/btm403.
O'Rourke SM, Herskowitz I: Unique and redundant roles for HOG MAPK pathway components as revealed by whole-genome expression analysis. Mol Biol Cell. 2004, 15: 532-542. 10.1091/mbc.E03-07-0521.
Fruchart JC, Staels B, Duriez P: PPAR-alpha in lipid and lipoprotein metabolism, vascular inflammation and atherosclerosis. Process in Experimental Cardiology. 2003, 8: 3-16.
Gilde AJ, Van Bilsen M: Peroxisome proliferator-activated receptors (PPARS): regulators of gene expression in heart and skeletal muscle. Acta Physiol Scand. 2003, 178: 425-434. 10.1046/j.1365-201X.2003.01161.x.
Bauer BE, Wolfger H, Kuchler K: Inventory and function of yeast ABC proteins: about sex, stress, pleiotropic drug and heavy metal resistance. Biochim Biophys Acta. 1999, 1461: 217-236. 10.1016/S0005-2736(99)00160-1.
Wallace DC: Mitochondrial diseases in man and mouse. Science. 1999, 283: 1482-1488. 10.1126/science.283.5407.1482.
Pan MH, Chang YH, Badmaev V, Nagabhushanam K, Ho CT: Pterostilbene induces apoptosis and cell cycle arrest in human gastric carcinoma cells. J Agric Food Chem. 2007, 55: 7777-7785. 10.1021/jf071520h.
Tai SL, Snoek I, Luttik MA, Almering MJ, Walsh MC, Pronk JT, Daran JM: Correlation between transcript profiles and fitness of deletion mutants in anaerobic chemostat cultures of Saccharomyces cerevisiae. Microbiology. 2007, 153: 877-886. 10.1099/mic.0.2006/002873-0.
Malavazi I, Savoldi M, Di Mauro SM, Menck CF, Harris SD, Goldman MH, Goldman GH: Transcriptome analysis of Aspergillus nidulans exposed to camptothecin-induced DNA damage. Eukaryot Cell. 2006, 5: 1688-1704. 10.1128/EC.00167-06.
Birrell GW, Brown JA, Wu HI, Giaever G, Chu AM, Davis RW, Brown JM: Transcriptional response of Saccharomyces cerevisiae to DNA-damaging agents does not identify the genes that protect against these agents. Proc Natl Acad Sci USA. 2002, 99: 8778-8783. 10.1073/pnas.132275199.
Giaever G, Chu AM, Ni L, Connelly C, Riles L, Veronneau S, Dow S, Lucau-Danila A, Anderson K, Andre B, Arkin AP, Astromoff A, El-Bakkoury M, Bangham R, Benito R, Brachat S, Campanaro S, Curtiss M, Davis K, Deutschbauer A, Entian KD, Flaherty P, Foury F, Garfinkel DJ, Gerstein M, Gotte D, Guldener U, Hegemann JH, Hempel S, Herman Z, et al: Functional profiling of the Saccharomyces cerevisiae genome. Nature. 2002, 418: 387-391. 10.1038/nature00935.
Winzeler EA, Shoemaker DD, Astromoff A, Liang H, Anderson K, Andre B, Bangham R, Benito R, Boeke JD, Bussey H, Chu AM, Connelly C, Davis K, Dietrich F, Dow SW, El Bakkoury M, Foury F, Friend SH, Gentalen E, Giaever G, Hegemann JH, Jones T, Laub M, Liao H, Liebundguth N, Lockhart DJ, Lucau-Danila A, Lussier M, M'Rabet N, Menard P, et al: Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science. 1999, 285: 901-906. 10.1126/science.285.5429.901.
Parsons AB, Lopez A, Givoni IE, Williams DE, Gray CA, Porter J, Chua G, Sopko R, Brost RL, Ho CH, Wang J, Ketela T, Brenner C, Brill JA, Fernandez GE, Lorenz TC, Payne GS, Ishihara S, Ohya Y, Andrews B, Hughes TR, Frey BJ, Graham TR, Andersen RJ, Boone C: Exploring the mode-of-action of bioactive compounds by chemical-genetic profiling in yeast. Cell. 2006, 126: 611-625. 10.1016/j.cell.2006.06.040.
Baetz K, McHardy L, Gable K, Tarling T, Reberioux D, Bryan J, Andersen RJ, Dunn T, Hieter P, Roberge M: Yeast genome-wide drug-induced haploinsufficiency screen to determine drug mode of action. Proc Natl Acad Sci USA. 2004, 101: 4525-4530. 10.1073/pnas.0307122101.
Giaever G, Flaherty P, Kumm J, Proctor M, Nislow C, Jaramillo DF, Chu AM, Jordan MI, Arkin AP, Davis RW: Chemogenomic profiling: identifying the functional interactions of small molecules in yeast. Proc Natl Acad Sci USA. 2004, 101: 793-798. 10.1073/pnas.0307490100.
Lum PY, Armour CD, Stepaniants SB, Cavet G, Wolf MK, Butler JS, Hinshaw JC, Garnier P, Prestwich GD, Leonardson A, Garrett-Engele P, Rush CM, Bard M, Schimmack G, Phillips JW, Roberts CJ, Shoemaker DD: Discovering modes of action for therapeutic compounds using a genome-wide screen of yeast heterozygotes. Cell. 2004, 116: 121-137. 10.1016/S0092-8674(03)01035-3.
Lee PS, Shaw LB, Choe LH, Mehra A, Hatzimanikatis V, Lee KH: Insights into the relation between mrna and protein expression patterns: II. Experimental observations in Escherichia coli. Biotechnol Bioeng. 2003, 84: 834-841. 10.1002/bit.10841.
Lu P, Vogel C, Wang R, Yao X, Marcotte EM: Absolute protein expression profiling estimates the relative contributions of transcriptional and translational regulation. Nat Biotechnol. 2007, 25: 117-124. 10.1038/nbt1270.
Nie L, Wu G, Zhang W: Correlation of mRNA expression and protein abundance affected by multiple sequence features related to translational efficiency in Desulfovibrio vulgaris: a quantitative analysis. Genetics. 2006, 174: 2229-2243. 10.1534/genetics.106.065862.
Wang Y, Chiu JF, He QY: Proteomics approach to illustrate drug action mechanisms. Curr Drug Discov Technol. 2006, 3: 199-209. 10.2174/157016306780136763.
Thorsen M, Lagniel G, Kristiansson E, Junot C, Nerman O, Labarre J, Tamas MJ: Quantitative transcriptome, proteome, and sulfur metabolite profiling of the Saccharomyces cerevisiae response to arsenite. Physiol Genomics. 2007, 30: 35-43. 10.1152/physiolgenomics.00236.2006.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1755-8794/1/7/prepub
Acknowledgements
The authors are grateful to Jamie Lamberton and Melanie Mask for excellent technical assistance. AKA, TX, and QF were supported in part by a grant from the Public Health Service, National Institute of Allergy and Infectious Diseases, Grant No. R01 AI27094, and the USDA Agricultural Research Service Specific Cooperative Agreement No. 58-6408-2-0009.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author(s) declare that they have no competing interests.
Authors' contributions
ZP conceived and coordinated the study, performed the data analysis and drafted the manuscript. AKA participated in the design of the study, contributed to the data analysis, and helped to draft the manuscript. TX carried out the yeast mutant sensitivity assays and RT-PCR primer design. SRB performed the real-time quantitative RT-PCR experiments. AMR synthesized the compound for this study. QF performed the IC50 determination and the DNA microarray experiments. SOD provided intellectual input and guidance, and assisted with manuscript preparation. All authors read and approved the final manuscript.
Electronic supplementary material
12920_2007_7_MOESM1_ESM.pdf
Additional file 1: Correlation between expression ratios obtained from quantitative real-time RT-PCR and microarray experiments. (PDF 79 KB)
12920_2007_7_MOESM3_ESM.xls
Additional file 3: List of pterostilbene-responding genes in the "cell wall," "transcription factor activity," and "mitochondrion" categories. (XLS 53 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Pan, Z., Agarwal, A.K., Xu, T. et al. Identification of molecular pathways affected by pterostilbene, a natural dimethylether analog of resveratrol. BMC Med Genomics 1, 7 (2008). https://doi.org/10.1186/1755-8794-1-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1755-8794-1-7