
Abstract
Background
The P-glycoprotein, encoded by the ABCB1 gene, is expressed in human endothelial and mesangial cells, which contribute to control renal plasma flow and glomerular filtration rate. We investigated the association of ABCB1 variants with renal function in African and Caucasian subjects.
Methods
In Africans (290 subjects from 62 pedigrees), we genotyped the 2677G>T and 3435 C>T ABCB1 polymorphisms. Glomerular filtration rate (GFR) was measured using inulin clearance and effective renal plasma flow (ERPF) using para-aminohippurate clearance. In Caucasians (5382 unrelated subjects), we analyzed 30 SNPs located within and around ABCB1, using data from the Affymetrix 500 K chip. GFR was estimated using the simplified Modification of the Diet in Renal Disease (MDRD) and Cockcroft-Gault equations.
Results
In Africans, compared to the reference genotype (GG or CC), each copy of the 2677T and 3435T allele was associated, respectively, with: GFR higher by 10.6 ± 2.9 (P < 0.001) and 4.4 ± 2.3 (P = 0.06) mL/min; ERPF higher by 47.5 ± 11.6 (P < 0.001) and 28.1 ± 10.5 (P = 0.007) mL/min; and renal resistances lower by 0.016 ± 0.004 (P < 0.001) and 0.011 ± 0.004 (P = 0.004) mm Hg/mL/min. In Caucasians, we identified 3 polymorphisms in the ABCB1 gene that were strongly associated with all estimates of GFR (smallest P value = 0.0006, overall P = 0.014 after multiple testing correction).
Conclusion
Variants of the ABCB1 gene were associated with renal function in both Africans and Caucasians and may therefore confer susceptibility to nephropathy in humans. If confirmed in other studies, these results point toward a new candidate gene for nephropathy in humans.
Similar content being viewed by others
Background
A better knowledge of the determinants of renal function is of paramount importance considering the high and increasing burden of chronic kidney disease worldwide. [1] The familial aggregation of renal function [2–4] suggests that genetic factors determine, in part, renal function and that there are candidate genes for nephropathy in humans.
The transmembrane efflux-P-glycoprotein (PGP; also known as multidrug resistance-associated protein 1) is encoded by the ABCB1 gene, several genetic variants of which have been shown to influence PGP expression in humans. PGP has been extensively studied in a pharmacogenetic context, in particular for its role in multi-drug resistance in cancer treatment. PGP is widely expressed in the human kidney [5–7] and is involved in ciclosporin-induced post-transplantation nephrotoxicity [5, 8, 9], possibly because of its influence on ciclosporin absorption. [10]ABCB1 genetic variants have been associated with post-transplantation ciclosporin-induced nephrotoxicity. [10–12] In contrast to the extensive data about the physiological role of PGP on the transport of xenobiotics, little is known about the role of PGP in the transport of endogenous substrates and to our knowledge, ABCB1 gene variants have not been previously associated with renal function in the absence of xenobiotics in humans. [13]
Since PGP is expressed in human endothelial and mesangial cells, which contribute to the control of renal plasma flow and glomerular filtration, we hypothesized that variants in the ABCB1 gene could be associated with renal hemodynamics and glomerular filtration rate (GFR), even in the absence of treatment by PGP substrates and/or modulators, such as ciclosporin. In particular, we investigated whether the 3435 C>T and the 2677G>T ABCB1 variants are associated with GFR, effective renal plasma flow (ERPF) and renal vascular resistance (RVR) in families of African descent. We then confirmed an association of variants in the ABCB1 gene with renal function using the 30 genetic markers located within the ABCB1 gene in a large ongoing population-based genome-wide association study of Caucasians employing the Affymetrix 500 K chip.
Methods
Seychelles study
Study population
In the Seychelles islands (Indian Ocean, African region), we enrolled 494 subjects of East African descent from 76 families enriched in hypertensive individuals between August 1999 and January 2002. The detailed family selection process has been previously described. [2] Renal hemodynamics and ABCB1 genotype were determined in 297 individuals. We excluded seven individuals with extreme outlier values for any of the renal measures, i.e. observations lying beyond three interquartile ranges from the first and third quartiles, leaving 290 individuals to analyze the associations of interest. The study was approved by the Ethical Committees of the Ministry of Health in the Seychelles and of the University of Lausanne (Switzerland). All participants provided written informed consent.
Phenotype and covariate measurements
In the Seychelles study, antihypertensive therapy, if any, was stopped for 2 weeks before clearance protocols. After a two-hour equilibration period, two one-hour inulin and PAH clearances were obtained to measure GFR and ERPF, respectively [see Additional file 1]. The inulin and PAH clearances (Cx) were calculated with the formula Cx = Ux*V/Px, where Ux and Px are urinary and plasma concentrations of the x solute, and V is the urine flow rate in ml/min. Renal blood flow (RBF) was calculated as ERPF/(1-(haematocrit/100)) and RVR as (mean arterial blood pressure)/RBF. GFR was also estimated using the abbreviated Modification of the Diet in Renal Disease (MDRD) equation [14]. Creatinine concentration was measured by the picric acid method (Cobas-Mira, Roche, Basel, Switzerland). Mean arterial blood pressure (MAP) was calculated as diastolic blood pressure plus 1/3 of (systolic minus diastolic blood pressure) from the mean of 6 measurements taken with a mercury sphygmomanometer (3 on the day preceding clearances and 3 on the morning of the clearances). On the day preceding clearance, participants collected urine for 24 hours. Urinary and plasma sodium and potassium concentrations were measured by flame photometry (IL-943, Instrumentation Laboratory, Milan, Italy). Participants on antidiabetic treatment during the preceding month, or with fasting blood glucose ≥ 7.0 mmol/l (measured on venous whole blood in duplicate using a Glycotronic® C reflectometer, Macherey-Nagel, Düren, Germany), were considered as diabetics. Body surface area was calculated using the Dubois formula. [15] Body mass index (BMI) was calculated as weight (kg) divided by squared height (m2).
Genetic analyses
DNA was isolated using standard methods from blood drawn into K-EDTA tubes and stored at 4°C. The ABCB1 genotypes (exon 21, 2677 G>T [rs2032582] and exon 26, 3435 C>T [rs1045642]) were determined by real-time PCR with TaqMan®, as previously described. [16, 17] The marker genotypes did not significantly deviate from Hardy-Weinberg proportions (P > 0.09) in founders, and 2677 G>T and 3435 C>T were in strong linkage disequilibrium (D' = 0.90) with each other.
Statistical analyses
We used the ASSOC program in S.A.G.E. (V5.3) [18], which accounts for familial correlations by implementing simultaneous maximum likelihood estimation of both familial components of variance and covariate coefficients, to conduct linear regressions with GFR, ERPF or RVR as dependent variables. Based on the findings from descriptive analyses in our sample (i.e. GT heterozygotes had phenotypic values lying between those of the GG and TT homozygotes), we assumed an additive mode of action for the ABCB1 3435T and 2677T alleles. To conduct analyses stratified by hypertension status, we defined hypertensive subjects as those on antihypertensive treatment at entry into the study or having, respectively, a systolic and/or diastolic blood pressure = 140/90 mm Hg. For multiple linear regression models, we used as predictors age, sex, BMI, 24-hour urinary sodium and potassium excretion, fasting blood glucose, diabetes and MAP. All multivariable models included ascertainment as previously described. [19] We conducted sensitivity analyses that included extreme outlier values. We formally tested whether normotensive and hypertensive subjects had similar results by including a test of hypertension status by genotype interaction (i.e. a likelihood ratio test with 2 degrees of freedom) in multivariable models. To ensure that any associations were not due to population stratification: (1) because some subjects were of mixed descent, we conducted a sensitivity analysis that included only those subjects (78%) with known grand-parent's ethnicity who reported having at least 3 grand-parents of African descent (n = 153), and (2) we extended our analysis of GFR by a separate analysis of 119 subjects from the same families with missing inulin clearance but available MDRD. We also conducted analyses based on haplotypes. For the 61 doubly heterozygous participants (CT/GT) with a potentially ambiguous phase, ABCB1 haplotypes were inferred using the HAPLORE program that uses an expectation-maximisation (EM) algorithm in pedigree data. [20]
CoLaus study
Study population
A simple non-stratified random sample (n = 56,694) representing 35% of the overall population of the city of Lausanne (Switzerland) aged 35–75 years was drawn. Inclusion criteria were: a) written informed consent; b) aged 35–75 years; c) available examination and blood sample and d) Caucasian descent. The study was approved by the ethical committee of the Faculty of Medicine of Lausanne. Recruitment began in June 2003 and ended in May 2006. Among those eligible, the participation rate was 41%. Of the 6188 participants, 781 were excluded because of missing or low quality genotyping data, genetic relatedness with other subjects, or missing phenotyping data, leaving 5407 subjects for the present analyses.
Phenotype and covariate measurements
Venous blood samples (50 mL) were drawn after an overnight fast, and clinical chemistry assays were performed by the CHUV Clinical Laboratory on fresh blood samples on a Modular P apparatus (Roche Diagnostics, Switzerland) within 2 hours of blood collection. Serum creatinine was measured by the Jaffe kinetic compensated method (2.9% – 0.7% maximum inter and intra-batch CVs). GFR was estimated using the abbreviated MDRD equation [14] and the Cockcroft-Gault (CG) formula [21]. Body weight and height were measured with participants standing without shoes in light indoor clothes. Smoking was defined as present if a participant reported to be a current smoker at the time of examination, and alcohol consumption as present for participants reporting drinking alcohol at least once a day. Diabetes was defined as being on current antidiabetic treatment or having fasting plasma blood glucose ≥ 7 mmol/L.
Genetic analyses
Nuclear DNA was extracted from whole blood for whole genome scan analysis. Genotyping was performed on 6014 participant samples, using the Affimetrix 500 K SNP chip, as recommended by the manufacturer. Individuals with less than 95% genotyping efficiency overall (or <90% efficiency on either array, N = 399), and individuals with possible gender inconsistencies (N = 5), were removed. Duplicate samples, and first and second degree relatives were identified by estimating genomic identity-by-descent (i.b.d.) coefficients; the younger individual from each such pair was removed (N = 200). A further 3 individuals had missing phenotype data, leaving 5407 individuals with complete data available for analysis. Monomorphic SNPs (N = 4052), SNPs with less than 70% genotyping efficiency (N = 157), and SNPs not in Hardy-Weinberg proportions (N = 35417) were excluded from analysis.
Statistical analyses
A more detailed description of the statistical analyses can be viewed in the Additional file 1. We excluded individuals with outlier phenotypic values defined as values lying more than 4 SD away from the mean (N = 25), leaving 5382 individuals for association tests. Phenotype data were transformed and adjusted for covariates using R. [22] We used age, height, weight and BMI as continuous covariates, and sex, alcohol consumption, smoking, diabetes, antidiabetic or antihypertensive treatment, the presence (or absence) of each of the major classes of antihypertensive drugs (ACE-inhibitors, angiotensin receptor blockers, beta-blockers, channel calcium blockers and diuretics) and currently (i.e. during the last 6 months) taking any prescription drug as binary covariates in normal linear regression. To confirm the association, we used only directly genotyped SNPs first and imputed SNPs in further exploratory analyses. Association tests were performed using PLINK (v0.99s) [23], with an additive mode of action.
Results
Seychelles Study
The allele frequency was 34.1% overall and 30.8% in founders for 2677T, and 54.5% and 53.9%, respectively, for 3435T. Participants' characteristics by genotypes and allele carrier status are presented in Table 1. There was no significant trend across genotypes. Carriers of the 3435T allele had a higher BMI than non-carriers (28.8 ± 0.4 versus 27.3 ± 0.5 kg/m2; P = 0.02).
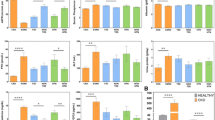
The 2677T allele was associated, in an additive manner, with higher GFR (P = 0.003) and ERPF (P = 0.005) and lower RVR (P = 0.001) (Figure 1). Results were similar, although slightly less significant with the 3435T allele. These results suggest that the T allele is a protective allele with respect to renal function. We obtained similar results for renal haemodynamic phenotypes adjusted for body surface area (Table 2). It is therefore unlikely that the difference in BMI observed between 3435T carriers and non-carriers represents a major confounding factor for these associations.

Renal haemodynamics by genotypes for each ABCB1 variant. Bar heights are means and vertical lines standard errors. P: P value for linear trend across genotypes using the ASSOC program in S.A.G.E. in models without additional covariates. GFR: glomerular filtration rate measured using inulin clearance. ERPF: effective renal plasma flow measured using PAH clearance. RVR: renal vascular resistance.
Because PGP is responsible for the transport of many drugs, we conducted analyses stratified by hypertension status (Figure 2). As the association was similar in normotensive subjects and hypertensive subjects, it is unlikely that our results reflect a confounding effect of antihypertensive drugs. The hypertension status by genotype interaction was not significant (P > 0.05) in multivariable models, which confirms that the associations were similar in normotensive and hypertensive subjects.

Renal haemodynamics by hypertension status for the 2677 C>T ABCB1 variants. Bar heights are means and vertical lines standard errors. P: P value for linear trend across genotypes using the ASSOC program in S.A.G.E. in models without additional covariates. GFR: glomerular filtration rate measured using inulin clearance. ERPF: effective renal plasma flow measured using PAH clearance. RVR: renal vascular resistance.
Table 3 presents regression coefficients of the 2677T and the 3435T allele, respectively, for the association with the renal haemodynamic phenotypes. Models adjusted for only age and sex differed little (i.e. by less than 30 percent of a standard error; P > 0.60) from complete multivariable models, which suggests that the extra covariate adjustment had little influence on the results. In multivariable models, each copy of the 2677T allele was associated with a 10.6 mL/min higher GFR, a 47.5 mL/min higher ERPF, and a 0.016 lower RVR as compared to the GG reference genotype (P < 0.0001). While the effect sizes of the 2677T allele were about 10% of the trait values, those of the 3435T allele were smaller and less significant. These results suggest that, even after accounting for major known confounding factors, ABCB1 genetic variants were associated with renal function in this large sample. The results of sensitivity analyses including extreme outlier values did not differ by more than 1 standard error from the analyses conducted after excluding extreme outliers (P > 0.40).
Analyses restricted to subjects with at least 3 grand-parents of African descent showed results that were similar in direction and significance level to those obtained in the whole sample. There was also a trend toward a similar association of GFR estimated using MDRD with the 2677T (8.6 ± 5.8 P = 0.14) and the 3435T (10.1 ± 5.3, P = 0.058) alleles in analysis of MDRD in the 119 subjects for whom inulin clearance values were missing.
Haplotype analyses showed that each copy of the TT haplotype was associated with an 11.6 ml/min higher GFR, a 50.7 mL/min higher ERPF, and a 0.017 mm Hg/mL/min lower RVR than the reference GC haplotype (P < 0.001). These results suggest that most of the signal was carried by the 2677T allele because the 3435T allele adds only 1 mL/min GFR or 3 ml/min ERPF.
CoLaus study
The 2526 men and 2856 women analyzed were aged 53.4 ± 10.7 years (mean ± SD), had mean 25.8 ± 4.5 kg/m2 BMI and 79.6 ± 14.6 μmol/L serum creatinine. The SNPs measured in the Seychelles (rs1045642 [3435 C>T], genotyped in CoLaus, and rs2032582 [2677 G>T], which is tagged by 3 genotyped SNPs at r2 > 0.9 and 2 further genotyped SNPs at r2 > 0.5) did not show any association with any estimate of GFR [see Additional file 2]. We did however find strong evidence of association between GFR and other SNPs in ABCB1. Testing the 30 directly genotyped SNPs within 10 kb of ABCB1, and using square-root transformed MDRD, we obtained P values of 0.0006, 0.0007, 0.0071, 0.0079 and 0.0085 for the five most significant SNPs, using 105 permutations. All other SNPs had P values > 0.01 [see Additional file 2]. The overall permutation P value for the most significant SNP, corrected for multiple testing, was P = 0.014. We take this as positive evidence for association.
The significance levels achieved here were sensitive to the removal of 25 outlying individuals (using a 4 SD cutoff). The overall permutation P value increased to 0.033 when only 13 outliers were excluded (using a 5 SD cutoff), and increased further to 0.10 (non-significant) when no outliers were excluded. Using other estimates of GFR, the associations were slightly less significant. For square-root transformed CG, P = 0.0007, 0.0018, 0.0079, 0.0046 and 0.0103 for the same top five SNPs, and P = 0.018 overall (i.e. corrected for multiple testing). For log transformed creatinine, P = 0.0009, 0.0010, 0.0060, 0.0069 and 0.0073 for the top five SNPs and P = 0.017 overall.
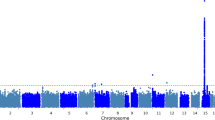
To explore the possibility that SNPs not genotyped on the Affymetrix 500 K chip might be associated with GFR, we repeated our analyses using imputed SNP genotypes over a 400 kb region surrounding ABCB1. Results both with and without outlying individuals removed are shown in Figure 3. Three non-coding SNPs, including our top two hits listed above, showed strong associations (P < 0.001) [see Additional file 2].

Genetic associations with GFR, in the region around the ABCB1 gene. Associations are shown for both directly genotyped and imputed SNPs. The bottom panel shows the pattern of linkage disequilibrium in the HapMap CEU panel. We imputed genotypes for all HapMap SNPs in the region around ABCB1. In this region, linkage disequilibrium patterns in CoLaus were similar to the ones observed in HapMap, although they were resolved at a coarser scale due to lower SNP density. Since it was not computationally feasible to combine imputation and permutation approaches, we plot P values calculated assuming the normal linear model. For directly genotyped SNPs, the differences between calculated and permutation P values were small. The top three hits (and P values) are rs17327624 (0.0006), rs4148733 (0.0008) and rs17327442 (0.0008). The former two were directly genotyped and are in strong LD (D' = 0.96 in CoLaus and 1.00 in HapMap CEU, r2 = 0.63 and 0.72 respectively), and the imputed SNP rs17327442 is in perfect LD with rs17327624 in HapMap CEU.
In analyses restricted to 2224 individuals not taking any prescription drugs (excluding 3 outlying individuals at a 5 SD cutoff), 5 SNPs had permutation P values less than 0.05, and the overall permutation P value corrected for multiple testing was 0.11. Given the expected reduced power in this smaller sample size, we conclude that a similar association is observed in untreated subjects.
Discussion
In the Seychelles' sample, we measured renal function using the accepted gold standard, inulin clearance, and found that ABCB1 genetic variants are associated with the haemodynamic profile, i.e. GFR, ERPF and RVR in these individuals of African descent with a positive family history of hypertension. We confirmed this association of the ABCB1 gene with renal function, estimated using the simplified MDRD equation, in a large population-based study of Caucasian subjects. This is, to our knowledge, the first time that the ABCB1 gene has been shown to be associated with renal function in the general population.
As this is not a true replication study (i.e. different ABCB1 marker variants were found to be associated with renal function in the two populations), these results are preliminary and need to be confirmed in other studies. This may result from the fact that the Seychelles and Swiss populations have different ethnic backgrounds and different linkage disequilibrium structures. In addition, the Seychelles sample was enriched in hypertensive individuals, whereas the Swiss one was not, which may further explain differences in findings between the two populations. We corrected our models for ascertainment (i.e. for the fact that recruitment was constrained to include two hypertensive siblings in each family). Such a correction aims to determine what the results would have been had the participants not been ascertained this way. This correction results in better generalizability of our observations.
In the African sample, (1) analyses restricted to subjects with at least 3 grand-parents of African descent led to very similar results; and (2) we confirmed the association with GFR estimated using MDRD in an analysis that included 119 additional African subjects. These results are thus unlikely to be due to population stratification. The confirmation of an association in the Caucasian sample with variants in the same gene adds further evidence against a spurious association.
The frequency of the 2677T allele typically varies between 0–10% in subjects of African descent, 39–46% in Caucasians and 36–44% in Asians [24], whereas the frequency of the 3435T allele typically varies between 16–27% in subjects of African descent, 48–57% in Caucasians, and 41–66% in Asians. [25] Of note, the frequencies of the 2677T (31%) and 3435T (54%) alleles in our sample are higher than those reported in other groups of African descent. The Seychelles' population is a relatively young population (the islands were first inhabited at the end of the 18th century) of small size (currently 84,000 inhabitants) that has undergone a population bottleneck during the middle of the 19th century followed by rapid growth (census data). Genetic drift may explain why the allele frequency differs in Seychelles versus other African populations (in Africa or elsewhere). It is not clear to what extent our observations may reflect the facts that homozygosity at the 3435T allele is a marker of African descent and African ethnicity is a risk factor for developing chronic kidney disease. Given the large interethnic differences in allele frequencies observed for ABCB1 variants, it is important that the associations of ABCB1 variants with renal function are explored further in other ethnic groups.
The TT genotype of the C3435T polymorphism is associated with a significant reduction in PGP expression in various cell types, including renal proximal tubular cells. [26–29] Although synonymous (i.e. not leading to an amino-acid change in PGP), the 3435 C>T variant influences mRNA expression by acting on its stability [30] and on the substrate specificity of PGP [31]. The 2677 G>T variant is functional and leads to an Ala893Ser amino acid change in PGP. The 3435 C>T and 2677 G>T polymorphisms are in strong linkage disequilibrium. The fact that the 2677 G>T and 3435 C>T ABCB1 variants (and markers in strong linkage disequilibrium with these variants) were not associated with estimated GFR in the CoLaus study suggests that the true causal variant(s) is (are) in linkage disequilibrium with the variants genotyped in the Seychelles sample or that the two populations differ in either modifier genes or environmental components (e.g. the Balkanic toxin [32, 33]) affecting the impact of the causal variant. Further studies are needed to clarify how ABCB1 genetic variants are associated with renal function.
The association reported here between the ABCB1 gene and renal haemodynamics does not necessarily mean a causal relationship. However, several independent lines of evidence of an association with renal function strengthen our confidence that these results are not spurious. First, several experimental findings in vitro and in vivo [5, 8, 9] and findings in humans [11, 12] suggest that either PGP itself and/or ABCB1 genetic variants play a role in post-transplantation nephrotoxicity induced by calcineurin inhibitors, such as ciclosporine. Although inconsistent (i.e. the T allele is associated with nephrotoxicity in one study, but with nephroprotection in the other), these findings nevertheless suggest that PGP influences renal function via exogenous substances. Hence PGP may influence renal function via the control of intracellular concentration of a known toxin in the tubular epithelial cells. By analogy, we put could speculate that PGP influences renal function via the control of intracellular concentration of endogenous substances, by an as yet unknown mechanism. Second, Atanasova et al [32] found the ABCB1 haplotype 2677G/3435T to be protective against Balkan endemic nephropathy, a slowly progressive nephropathy associated with a high incidence of epithelial tumors of the upper urinary tract. Because of the very specific and stable geographical localization of this nephropathy, it has been proposed that an environmental toxin could play a role in its etiology. [33] However, the familial aggregation of this nephropathy suggests that genetic factors are also likely to be involved. [33] Third, endothelins are important regulators of renal blood flow and glomerular filtration rate [34] that can influence PGP-mediated transport [35, 36]. Bosentan, an endothelin receptor antagonist, activates the pregnane X receptor [37], which itself activates ABCB1 expression [38].
Conclusion
In conclusion, we found that selected ABCB1 gene variants were associated with renal function and haemodynamics in families of African descent, while other ABCB1 gene variants were associated with GFR estimates (MDRD and CG) in another study of Caucasian individuals. Although the associations were not based on the same markers in the two populations, these results suggest that variants of the ABCB1 gene influence renal function in the general population. The ABCB1 gene therefore represents a new candidate gene for nephropathy in humans. Replication in additional studies is warranted.
Abbreviations
- PGP:
-
P-glycoprotein
- GFR:
-
glomerular filtration rate
- ERPF:
-
effective renal plasma flow
- MDRD:
-
Modification of the Diet in Renal Disease
- CG:
-
Cockcroft-Gault
- RVR:
-
renal vascular resistance
- MAP:
-
Mean arterial blood pressure
- BMI:
-
body mass index
References
Meguid El NA, Bello AK: Chronic kidney disease: the global challenge. Lancet. 2005, 365: 331-340.
Bochud M, Elston RC, Maillard M, Bovet P, Schild L, Burnier M: Heritability of renal function in hypertensive families of African descent in the Seychelles (Indian Ocean). Kidney Int. 2005, 67: 61-69. 10.1111/j.1523-1755.2005.00055.x.
Lei HH, Perneger TV, Klag MJ, Whelton PK, Coresh J: Familial aggregation of renal disease in a population-based case-control study. J Am Soc Nephrol. 1998, 9: 1270-1276.
Langefeld CD, Beck SR, Bowden DW, Rich SS, Wagenknecht LE, Freedman BI: Heritability of GFR and albuminuria in Caucasians with type 2 diabetes mellitus. Am J Kidney Dis. 2004, 43: 796-800. 10.1053/j.ajkd.2003.12.043.
Hauser IA, Koziolek M, Hopfer U, Thevenod F: Therapeutic concentrations of cyclosporine A, but not FK506, increase P-glycoprotein expression in endothelial and renal tubule cells. Kidney Int. 1998, 54: 1139-1149. 10.1046/j.1523-1755.1998.00095.x.
Thiebaut F, Tsuruo T, Hamada H, Gottesman MM, Pastan I, Willingham MC: Cellular localization of the multidrug-resistance gene product P-glycoprotein in normal human tissues. Proc Nat Acad Sci U S A. 1987, 84: 7735-7738. 10.1073/pnas.84.21.7735.
Ernest S, Rajaraman S, Megyesi J, Bello-Reuss EN: Expression of MDR1 (multidrug resistance) gene and its protein in normal human kidney. Nephron. 1997, 77: 284-289.
Del Moral RG, Olmo A, Osuna A, Aguilar M, Carvia R, Becerra P, Arrebola F, Guillen M, Reguero ME, Asensio C, O'Valle F: Role of P-glycoprotein in chronic cyclosporine nephrotoxicity and its relationship to intrarenal angiotensin II deposits. Transplant Proc. 1998, 30: 2014-2016. 10.1016/S0041-1345(98)00515-6.
Koziolek MJ, Riess R, Geiger H, Thevenod F, Hauser IA: Expression of multidrug resistance P-glycoprotein in kidney allografts from cyclosporine A-treated patients. Kidney Int. 2001, 60: 156-166. 10.1046/j.1523-1755.2001.00782.x.
Foote CJ, Greer W, Kiberd BA, Fraser A, Lawen J, Nashan B, Belitsky P: MDR1 C3435T polymorphisms correlate with cyclosporine levels in de novo renal recipients. Transplant Proc. 2006, 38: 2847-2849. 10.1016/j.transproceed.2006.08.120.
Hauser IA, Schaeffeler E, Gauer S, Scheuermann EH, Wegner B, Gossmann J, Ackermann H, Seidl C, Hocher B, Zanger UM, Geiger H, Eichelbaum M, Schwab M: ABCB1 genotype of the donor but not of the recipient is a major risk factor for cyclosporine-related nephrotoxicity after renal transplantation. J Am Soc Nephrol. 2005, 16: 1501-1511. 10.1681/ASN.2004100882.
Hebert MF, Dowling AL, Gierwatowski C, Lin YS, Edwards KL, Davis CL, Marsh CL, Schuetz EG, Thummel KE: Association between ABCB1 (multidrug resistance transporter) genotype and post-liver transplantation renal dysfunction in patients receiving calcineurin inhibitors. Pharmacogenetics. 2003, 13: 661-674. 10.1097/00008571-200311000-00002.
Zolk O, Jacobi J, Pahl A, Fromm MF, Schmieder RE: MDR1 genotype-dependent regulation of the aldosterone system in humans. Pharmacogenet Genomics. 2007, 17: 137-144.
Levey AS, Coresh J, Balk E, Kausz AT, Levin A, Steffes MW, Hogg RJ, Perrone RD, Lau J, Eknoyan G: National Kidney Foundation Practice Guidelines for Chronic Kidney Disease: Evaluation, Classification, and Stratification. Ann Intern Med. 2003, 139: 137-147.
DuBois M, DuBois EF: A formula to estimate the approximate surface area if height and weight be known. Arch Intern Med. 1916, 17: 863-871.
Eap CB, Fellay J, Buclin T, Bleiber G, Golay KP, Brocard M, Baumann P, Telenti A: CYP3A activity measured by the midazolam test is not related to 3435 C>T polymorphism in the multiple drug resistance transporter gene. Pharmacogenetics. 2004, 14: 255-260. 10.1097/00008571-200404000-00005.
Bochud M, Eap CB, Elston RC, Bovet P, Maillard M, Schild L, Shamlaye C, Burnier M: Association of CYP3A5 genotypes with blood pressure and renal function in African families. J Hypertens. 2006, 24: 923-929. 10.1097/01.hjh.0000222763.84605.4a.
S.A.G.E. (Statistical Analysis for Genetic Epidemiology) version 5.3. 2006, [http://darwin.cwru.edu/sage/]
Bochud M, Bovet P, Elston RC, Paccaud F, Falconnet C, Shamlaye C, Burnier M: High heritability of ambulatory blood pressure in families of East African descent. Hypertension. 2005, 45: 445-450. 10.1161/01.HYP.0000156538.59873.86.
Zhang K, Sun F, Zhao H: HAPLORE: a program for haplotype reconstruction in general pedigrees without recombination. Bioinformatics. 2005, 21: 90-103. 10.1093/bioinformatics/bth388.
Cockcroft DW, Gault MH: Prediction of creatinine clearance from serum creatinine. Nephron. 1976, 16: 31-41. 10.1159/000130554.
Team RDC: R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing,. 2007, Vienna, Austria, [http://www.R-project.org]
Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D, Maller J, Sklar P, de Bakker PI, Daly MJ, Sham PC: PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007, 81: 559-575. 10.1086/519795.
Kurzawski M, Pawlik A, Gornik W, Drozdzik M: Frequency of common MDR1 gene variants in a Polish population. Pharmacol Rep. 2006, 58: 35-40.
Ameyaw MM, Regateiro F, Li T, Liu X, Tariq M, Mobarek A, Thornton N, Folayan GO, Githang'a J, Indalo A, Ofori-Adjei D, Price-Evans DA, McLeod HL: MDR1 pharmacogenetics: frequency of the C3435T mutation in exon 26 is significantly influenced by ethnicity. Pharmacogenetics. 2001, 11: 217-221. 10.1097/00008571-200104000-00005.
Hoffmeyer S, Burk O, von Richter O, Arnold HP, Brockmoller J, Johne A, Cascorbi I, Gerloff T, Roots I, Eichelbaum M, Brinkmann U: Functional polymorphisms of the human multidrug-resistance gene: Multiple sequence variations and correlation of one allele with P-glycoprotein expression and activity in vivo. Proc Nat Acad Sci U S A. 2000, 97: 3473-3478. 10.1073/pnas.050585397.
Owen A, Chandler B, Bray PG, Ward SA, Hart CA, Back DJ, Khoo SH: Functional Correlation of P-Glycoprotein Expression and Genotype with Expression of the Human Immunodeficiency Virus Type 1 Coreceptor CXCR4. J Virol. 2004, 78: 12022-12029. 10.1128/JVI.78.21.12022-12029.2004.
Fellay J, Marzolini C, Meaden ER, Back DJ, Buclin T, Chave JP, Decosterd LA, Furrer H, Opravil M, Pantaleo G, Retelska D, Ruiz L, Schinkel AH, Vernazza P, Eap CB, Telenti A: Response to antiretroviral treatment in HIV-1-infected individuals with allelic variants of the multidrug resistance transporter 1: a pharmacogenetics study. Lancet. 2002, 359: 30-36. 10.1016/S0140-6736(02)07276-8.
Siegsmund M, Brinkmann U, Schaffeler E, Weirich G, Schwab M, Eichelbaum M, Fritz P, Burk O, Decker J, Alken P, Rothenpieler U, Kerb R, Hoffmeyer S, Brauch H: ASSociation of the P-Glycoprotein Transporter MDR1 C3435T Polymorphism with the Susceptibility to Renal Epithelial Tumors. J Am Soc Nephrol. 2002, 13: 1847-1854. 10.1097/01.ASN.0000019412.87412.BC.
Wang D, Johnson AD, Papp AC, Kroetz DL, Sadee W: Multidrug resistance polypeptide 1 (MDR1, ABCB1) variant 3435C>T affects mRNA stability. Pharmacogenet Genomics. 2005, 15: 693-704. 10.1097/01.fpc.0000175600.26893.fa.
Kimchi-Sarfaty C, Oh JM, Kim IW, Sauna ZE, Calcagno AM, Ambudkar SV, Gottesman MM: A "silent" polymorphism in the MDR1 gene changes substrate specificity. Science. 2007, 315: 525-528. 10.1126/science.1135308.
Atanasova S, von Ahsen N, Dimitrov T, Armstrong V, Oellerich M, Toncheva D: MDR1 haplotypes modify BEN disease risk: a study in Bulgarian patients with Balkan endemic nephropathy compared to healthy controls. Nephron Physiol. 2004, 96: e7-13. 10.1159/000075571.
Toncheva D, Dimitrov T, Stojanova S: Etiology of Balkan endemic nephropathy: a multifactorial disease?. Eur J Epidemiol. 1998, 14: 389-394. 10.1023/A:1007445120729.
Neuhofer W, Pittrow D: Role of endothelin and endothelin receptor antagonists in renal disease. Eur J Clin Invest. 2006, 36 Suppl 3: 78-88. 10.1111/j.1365-2362.2006.01689.x.
Masereeuw R, Terlouw SA, van Aubel RA, Russel FG, Miller DS: Endothelin B receptor-mediated regulation of ATP-driven drug secretion in renal proximal tubule. Mol Pharmacol. 2000, 57: 59-67.
Terlouw SA, Masereeuw R, Russel FG, Miller DS: Nephrotoxicants induce endothelin release and signaling in renal proximal tubules: effect on drug efflux. Mol Pharmacol. 2001, 59: 1433-1440.
van Giersbergen PLM, Gnerre C, Treiber A, Dingemanse J, Meyer UA: Bosentan, a dual endothelin receptor antagonist, activates the pregnane X nuclear receptor. European Journal of Pharmacology. 2002, 450: 115-121. 10.1016/S0014-2999(02)02075-7.
Masuyama H, Suwaki N, Tateishi Y, Nakatsukasa H, Segawa T, Hiramatsu Y: The Pregnane X Receptor Regulates Gene Expression in a Ligand- and Promoter- Selective Fashion. Mol Endocrinol. 2005, 19: 1170-1180. 10.1210/me.2004-0434.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1755-8794/1/21/prepub
Acknowledgements
We are indebted to all study participants. Parts of the results have been published in abstract format at the ASN's 39th 2006 Annual Meeting, San Diego, California. The computations were performed in part at the Vital-IT. Center for high-performance computing of the Swiss Institute of Bioinformatics. We thank the Ministry of Health of the Republic of Seychelles for their support of this epidemiological research; Air Seychelles and SkyChef for their logistic support in transporting material, Norma Both and Gylian Mein for data collection. The Seychelles study benefited from a grant from the Swiss National Science Foundation (TANDEM No 31-51115.97). M. Bochud is supported by a grant from the Swiss National Science Foundation (PROSPER 3200BO-111362/1 and 111361/1) and by the University Institute of Social and Preventive Medicine in Lausanne. R.C. Elston is supported by a U.S. Public Health Service resource grant (1 P41 RR03655) from the National Center for Research Resources and research grant (GM28356) from the National Institute of General Medical Sciences. S. Bergmann is supported by the Giorgi-Cavaglieri Foundation and the Swiss National Science Foundation (Grant # 3100AO-116323/1). M. Burnier is supported by the Swiss National Science Foundation (Project No 3100A0-103629). The CoLaus Study was supported by research grants from GlaxoSmithKline and from the Faculty of Biology and Medicine of Lausanne, Switzerland. The authors would like to express their gratitude to Yolande Barreau, Anne-Lise Bastian, Binasa Ramic, Martine Moranville, Martine Baumer, Marcy Sagette, Jeanne Ecoffey and Sylvie Mermoud for data collection, and to Gérard Waeber, Fred Paccaud, Allen Roses, and Lefkos T. Middleton for their support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests. Dawn Waterworth and Vincent Mooser are full-time employees of GlaxoSmithKline.
Authors' contributions
AG, CBE, DMW, MBu, MM, PB, PV and VM participated to the conduct and/or design of the studies. MBo, JSB, RCE, TJ and SB participated in the data analyses. DMW, MBo, MM, PV, and VM were involved in the data acquisition. All authors have been involved in drafting the manuscript or revising it critically for important intellectual content. All authors have given final approval of the version to be published.
Electronic supplementary material
12920_2008_21_MOESM1_ESM.doc
Additional file 1: Additional information on methods. The data provided represent additional information on phenotype and covariate measurements in the Seychelles study and statistical analyses in the CoLaus study (DOC 48 KB)
12920_2008_21_MOESM2_ESM.pdf
Additional file 2: Association of ABCB1 SNPs with square-root transformed MDRD in CoLaus. The data provided represent association data of genotyped and imputed SNPs located within and around the ABCB1 gene with square-root transformed MDRD in CoLaus. (PDF 33 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Bochud, M., Eap, C.B., Maillard, M. et al. Association of ABCB1genetic variants with renal function in Africans and in Caucasians. BMC Med Genomics 1, 21 (2008). https://doi.org/10.1186/1755-8794-1-21
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1755-8794-1-21