
Abstract
Experiments on use of an agar-gel method for recovery of migrating Ascaris suum larvae from the liver and lungs of pigs were conducted to obtain fast standardized methods. Subsamples of blended tissues of pig liver and lungs were mixed with agar to a final concentration of 1% agar and the larvae allowed to migrate out of the agar-gel into 0.9% NaCl at 38°C. The results showed that within 3 h more than 88% of the recoverable larvae migrated out of the liver agar-gel and more than 83% of the obtained larvae migrated out of the lung agar-gel. The larvae were subsequently available in a very clean suspension which reduced the sample counting time. Blending the liver for 60 sec in a commercial blender showed significantly higher larvae recovery than blending for 30 sec. Addition of gentamycin to reduce bacterial growth during incubation, glucose to increase larval motility during migration or ice to increase sedimentation of migrated larvae did not influence larvae recovery significantly.
Sammanfattning
Optimering af agar-gel metoden til isolering af migrerendeAscaris suum larver fra lever og lunger af svin.
Migrerende Ascaris suum larver kan isoleres fra lever og lunger af svin ved hjælp af en agar-gel teknik. Der er her foretaget en række undersøgelser af denne teknikfor at finde frem til en hurtig og standardiseret metode. Stikprøver af blendet lever- eller lungevæv blev blandet med agar til en slutkoncentration på 1% agar, hvorefter larverne vandrede ud af agar-gelen ved inkubering i 0.9% NaCl ved 38°C. Resultaterne viste, at >88% lever-larver og >83% lungelarver migrerede ud af agar-gelerne inden for de første 3 timer. Efter denne korte inkubering var larve-suspensionerne meget rene, hvilket reducerede det efterfølgende laboratorie-arbejde betydeligt. Det gav signifikant højere genfindelse af migrerende larver atblende levervæv i 60 sek sammenlignet med 30 sek. Derimod var der ingen effekt af tilsætning af gentamycin (for at reducere bakterie-vækst under inkuberingen), glucose (for at øge larvernes motilitet) eller is (for at øge sedimentationen af larver).
Similar content being viewed by others


Introduction
Simple microscopical quantification of Ascaris suum larvae migrating in the liver and lungs of pigs as carried out by e.g. [2] is very labourous, while the use of a modified Baermann technique on blended tissue [3] has been shown to reduce the work-load considerably. [7] and [9], however, developed an agar-gel technique for recovering nematode larvae from herbage samples, and this technique was later modified by [17] to isolate nematodes from the gastro-intestinal tract of sheep. Recently, the agar-gel technique has successfully been used for large scale recovery of minute A. suum larvae from pig intestinal contents [14], pig intestinal mucosa [8] and from mice tissues [15], where 97% of the present larvae were easily isolated in very clean suspensions. When using the same technique to recover tissue migrating larvae from blended liver and lung, [13, 16] found that comparable numbers of larvae could be obtained by the Macrobaermann technique and the agar-gel technique. However, the larval suspensions obtained from liver samples subjected to the agar-gel technique were much cleaner and therefore less time-consuming to count than the macrobaermann samples. A similar difference was not found for lung samples.
Various modifications of the agar-gel technique of [13, 16] for quantification of A. suum larvae in the liver and lungs of experimentally infected pigs have now been used in our laboratory (e.g. [5, 6, 4]), although there have been no systematic attempts to optimize the method or to evaluate the impact of the factors that may influence the recovery. Therefore, the aim of the present study was to optimize and standardize the agar-gel method for fast and reliable isolation of migrating A. suum larvae from pig livers and lungs.
Materials and methods
Experimental pigs
Twenty-four crossbred Danish Landrace/Yorkshire/Duroc pigs of 20–25 kg body weight were obtained from a helminth-free research farm (Sjælland III). The pigs had free access to water and were fed a standard ration of ground barley with a supplement of proteins, minerals and vitamins throughout the experiments.
Parasite
The CEP-strain of Ascaris suum was isolated in 1993 and since then maintained by passage in helminth naive pigs. The eggs were isolated from fresh faeces by sieving and cultured in vermiculite for 3 months at room temperature, and thereafter stored in tap water at 10°C.
Experimental protocol
Eight groups of 3 pigs were experimentally infected with infective A. suum eggs via stomach tube. Pigs of experiments 1–4 were each inoculated with 100.000 eggs and slaughtered day 4 post infection (pi), while pigs of experiments 5–8 received 10.000 eggs and were slaughtered 7 days pi. This design secured a high number of larvae from the liver day 4 pi and from the lungs day 7 pi [11].
Experiment 1
Effect of blending time and gentamycin. This experiment was designed to examine the rate of migration out of liver agar-gels (1, 2, 3 and 4 h), the effect of blending time and the effect of adding gentamycin to the samples.
The liver tissue blocks were blended for either 30 or 60 sec. At some occasions (see Table 1), gentamycin (60 μg/ml final concentration) was added to the blended tissue and the incubation jars. During incubation the gels were transferred one by one to new jars with saline (or saline + gentamycin) every 60 min for totally 4 h.
Experiment 2
Effect of incubation time, glucose and cooling. Here, we tested the effect of incubation time of the liver agar-gels (3 or 5 h), and addition of glucose to the saline plus the agar to promote larval migration out of the agar (see Table 2). Rapid cooling of the agar gel jars with ice (after removal of the gels) was also tested to prevent larvae from "swimming" in the jars.
Glucose was added to the saline of some of the samples to a final concentration of 1% in the agar-gels and the incubation saline. About 500 ml of crushed ice was added to some of the jars after the removal of the agar-gels to lower the temperature quickly.
Experiments 3 and 5
Effect of incubation time. The objective of these 2 experiments was to examine migratory ability of larvae from the liver (Exp. 3) and the lungs (Exp. 5) following incubation of agar-gels for varying length of time (1, 2, 3, 4 and 24 h).
Experiments 4, 6 and 7
Effect of sedimentation time in the jars. These experiments were designed to examine the effect of sedimentation time in the agar-gel jars after the removal of agar-gels with liver (Exp. 4, see Table 3) or lung (Exps. 6 and 7, see Table 4) samples. Furthermore, the effect of sedimentation time in the conical beakers was examined on both liver larvae (day 4 pi) and lung larvae (day 7 pi).
Experiment 8
Effect of sedimentation time in the beakers, cylinder glasses and gentamycin.
The objective of this experiment was to compare the sedimentations of lung larvae in 250 ml conical beakers with sedimentation in 500 ml cylinder glasses with vertical sides and to examine the possible effect of gentamycin on migration of lung larvae out of the agar-gels (see Table 5). Gentamycin was added to some of the lung samples as described for Exp. 1. After incubation of the lung agar-gels at 38°C for 3 h and removing the gels, the samples were mixed in 2 large buckets (one with normal samples and one with gentamycin samples), whereafter each of a series of 500 ml subsamples of saline with larvae was poured into 2 conical beakers or into a 500 ml cylinder glass. The supernatants, as well as the sediments, were examined for larvae.
The general agar-gel procedure
The pigs were killed using a captive bolt pistol, bled, and eviscerated. The livers (day 4 pi) and lungs (day 7 pi) were removed. The time interval between the slaughter of pigs and the start of the blending of tissue samples was about 1 h. The liver and lungs were examined for macroscopic lesions and cut into 2 × 2 × 5 cm tissue blocks and homogenized in a kitchen blender (Electronic, Braun, Germany) together with a small volume (about 10 ml) of normal saline for 60 sec unless otherwise specified (Exp.1), resulting in tissue pieces of 3–5 mm in diameter. Masses of 700–800 g of lung tissue or 800–1000 g of liver tissue were blended at a time. Fresh livers and lungs from an abattoir were used as supplements in Exps. 2 and 4 (livers) and Exps. 6, 7 and 8 (lungs) to increase the amount of tissue available for analysis. Pieces of tissue obtained from the abattoir were blended together with the infected organs and the whole mass of blended tissue from all pigs was thereafter mixed thoroughly for 10 min.
A modified version of the agar-gel method described by [13–16] was used. Subsamles of 200 g blended tissue were mixed with 0.9% NaCl (38°C) to 300 ml and subsequently mixed with 300 ml 2% agar solution (45°C) and immediately poured onto 3 horizontal trays (32 × 23 cm) with disposable cotton cloths. After the agar-gel had solidified, the 3 cloths with the adhering agar-gels were placed vertically in an incubation jar (34 × 23 × 6 cm) filled with warm (38°C) 0.9% NaCl and incubated in a room at 38°C.
Incubation of the agar-gels lasted 1–24 h as specified above, whereafter the gels were removed and the jar left to sediment at room temperature for 1–3 h as specified. The supernatant was aspirated and the sediment (approximately 400 ml) poured into 2 conical beakers (or a 500 ml glass cylinder, Exp. 4) at room temperature and left for 1/2–2 h. The supernatants were aspirated, and the sediments poured into a 50 ml tube and stored at 4°C. The larvae were allowed to settle overnight, whereafter they were counted at 40 × magnification using a stereomicroscope. At some occassions (see below) the supernatants normally discarded after sedimentation in the incubation jars and the conical beakers were examined for larvae by spinning the whole volumes down (7 min at 1200 rpm) before counting.
Statistical analyses
All statistical analyses were performed using a student's t-test (GraphPad Prism version 2.01, 1996).
Results
The larval suspensions obtained after 1–5 h incubation of gels with embedded liver or lung tissue were always very clean and especially the liver samples were much clearer than when the gels were incubated for 24 h. Furthermore, incubation of liver or lung agar-gels overnight at 38°C smelled bad compared to the short term incubation.
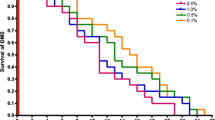
It was observed that significantly more A. suum larvae (p < 0.05) migrated out when the liver tissue was blended for 60 sec compared to 30 sec, and that gentamycin, did not influence migration (Table 1). Most larvae were found to migrate out of the gels within the first hours of incubation, and the migration rate seemed independent of blending time and addition of gentamycin. Table 2 shows that cooling with ice (reduced the temperature of the saline with 5–6°C) or addition of 1% glucose to the saline and agar did not influence recovery of the larvae from liver samples. Furthermore, most larvae had (as in exp.1) migrated out of the agar-gels within 3 h and there was no significant difference in larval numbers between 3 h and 5 h incubation. Therefore, 3 h incubation was used in the following experiments. Based on much higher larval counts, exp. 3 confirmed that most larvae (88%) migrated out of the liver agar samples within 3 h. Furthermore, larvae from embedded lungs tissue also migrated out in the largest numbers within the first few hrs of incubation (83% within 3 h, Fig. 1). Table 3 shows that sedimentation time in incubation jars and in conical beakers did not influence the recovered numbers of liver larvae significantly. The sedimentation time in exp. 6 for (1, 2 or 3 h) in the jars did not influence the recovery of lung tissue larvae significantly (the numbers were 67, 76 and 74 respectivly). This result was partly repeated in exp. 7 which also demonstrated that sedimentation time in conical beakers did not change the recovery significantly (Table 4). No larvae could be recovered from the supernatants of the conical sedimentation beakers. Conical sedimentation beakers gave a significantly higher yield of lung larvae compared to cylinder glasses (p < 0.05, Table 5), while the addition of gentamycin had no or even a significantly negative influence (Table 5).

Impact of incubation time of agar-gels on numbers of A. suum larvae recovered from liver and lung tissue estimated by transfer of the gels to new jars at selected time intervals. All samples: one h sedimentation in agar-gel jars, 2 h sedimentation in conical beakers, n = 8.
Discussion
After having been proved practically very useful for quantitative large-scale isolation of A. suum larvae migrating in pig livers and lungs, the agar-gel method of [13, 16] had with time been modified in the various experiments performed in our laboratory. Originally, [13, 16] blended 5-cm tissue block for 60 sec, incubated the agar-gels overnight (without use of antibiotics), and harvested the larvae directly on a 15 μm sieve (the other steps were largely as carried out in the present study). Afterwards, the sieving procedure for collecting the very small larvae has been replaced by sedimentation in the incubation jars followed by sedimentation in conical beakers, which is more safe. Thus, sedimentations replaced sieving in the experiments of [5, 6]. Furthermore, preliminary studies have indicated that most larvae leave the gels within the first few h of incubation (Jungersen, unpublished), thus incubation time has been reduced to 3 h in order to overcome the whole post mortem procedure within one day and to reduce the debris in the final samples (e.g. [6]). The latter quality saves time by facilitating the microscopical counting of larvae. [5, 6] also added antibiotics in order to reduce the bacterial growth during incubation and from small scale optimizations indicating higher recovery of lung larvae after addition of gentamycin (unpublished). Experiment 1 showed that blending time is critical, as 60 sec blending of 800–1000 g liver gave more than 3 times more larvae than 30 sec blending. Blending to very small tissue pieces on one hand facilitates the migration of larvae, while it on the other hand also must increase the number of larvae that become injuried, and which therefore may not be able to migrate in the gels afterwards (personal observation). The presently examined livers all had numerous white spots, but since they all origin from day 4 after a primary infection they may not yet have had the highly increased contents of connective tissue that characterises more chronically A. suum affected livers [18], and the degree of fibrosis may influence the outcome of blending, just like the sharpness of the knifes.
In the present study, gentamycin did not influence the larval recovery. Adding antibiotics was also suggested in order to reduce bacterial growth, but the reduction of the incubation period from 24 h to 3 h has largely eliminated this problem.
As the whole technique relies on the active migration of the larvae out of the gels, it was suggested that an easily available energy source, like 1% glucose, in the incubation saline and in the agar may increase the motility of the larvae and thereby increase the recovery. The results did not, however, indicate any effect on the liver larvae, which are the smallest and may be regarded as those which may be most sensitive to energy deficiency.
When observing macroscopically visible immature A. suum emerge from agar-embedded small intestinal contents, they are seen to be extremely active, being able to swim around in the saline of the incubation jars (personal observation). Therefore, we tested the effect of lowering the temperature of the incubation fluid with ice during sedimentation to immobilise the larvae. However, the decrease in temperature with only 5–6°C did not increase the recovery. Furthermore, repeated examinations of aspirated supernatants of both the incubation jars and the conical beakers only revealed a loss of 0–2% of the total numbers of recovered larvae, indicating that the problem with swimming, non-sedimented larvae is of minor importance. It may be concluded that sedimentations of 1 h in both incubation jars and conical beakers is sufficient. On the other hand, both liver and lung larvae may have been sufficient active so that they did not passively rest on the sides of the conical beakers, but assembled effectively on the bottom. This could also be the reason why cylindric sedimentation glasses did not give a higher recovery than the conical beakers, which has been found to be the case in other contexts [1].
The present results recommend a standardization of the agar-gel method for recovering of A. suum larvae from pig livers and lungs. This standard method includes a 60 sec blending of 800–1000 g liver tissue blocks or 700–800 g lung tissue blocks. A maximum of 300 ml sample/agar mixture on each cotton cloth, the agar should be incubated for exactly 3 h. By having this short incubation there is no need for addition of antibiotics. The sedimentation times in the incubation jars and the conical beakers should both be 1 h. This standard method is reproducible and in comparison with the original method of [13, 16] it is fast, results in very clean samples of larvae, and avoids the problematic sieving for concentration of the minute migrating larvae.
References
Andersen FL, Walters GT: Efficacy of the Baermann technique for recovery of Dictyocaulus viviparus larvae from bovine feces. Am J Vet Res. 1973, 34: 39-40.
Douvres FW, Troma FG, Malakatis GM: Morphogenesis and migration of Ascaris suum larvae developing to fourth stage in swine. J Parasitol. 1969, 55: 689-712. 10.2307/3277198.
Eriksen L, Nansen P, Roepstorff A, Lind P, Nilsson O: Response to repeated inoculations with Ascaris suum eggs in pigs during the fattening period. I. Studies on worm population kinetics. Parasitol Res. 1992, 78: 241-246. 10.1007/BF00931733.
Helwigh AB, Nansen P: Establishment of Ascaris suum in the pig: Development of immunity following a single primary infection. Acta Vet Scand. 1999, 40: 121-132.
Jungersen G, Fagerholm HP, Nansen P, Eriksen L: Development of patent Ascaris suum infections in pigs following intravenous administration of larvae hatched in vitro. Parasitology. 1999, 119: 503-508. 10.1017/S0031182099004928.
Jungersen G, Eriksen L, Roepstorff A, Lind P, Meeusen ENT, Rasmussen T, Nansen P: Experimental Ascaris suum infection in the pig: Protective memory response after three immunizations and effect of intestinal adult worm population. Parasite Immunol. 1999, 21: 619-630. 10.1046/j.1365-3024.1999.00261.x.
Jørgensen RJ: Isolation of infective Dictyocaulus larvae from herbage. Vet Parasitol. 1975, 1: 61-67. 10.1016/0304-4017(75)90008-4.
Murrell KD, Slotved H-C, Eriksen L, Bjerregaard J, Nansen P, Roepstorff A: Improved method for the recovery of Ascaris suum larvae from pig intestinal mucosa. J Parasitol. 1997, 83: 321-324. 10.2307/3284466.
Mwegoha WH, Jørgensen RJ: Recovery of infective 3rd stage larvae of Haemonchus contortus and Ostertagia ostertagi by migration in agar gel. Acta Vet Scand. 1977, 18: 293-299.
Oksanen A, Eriksen L, Roepstorff A, Ilsø B, Nansen P, Lind P: Embryonation and infectivity of Ascaris suum eggs. A comparison of eggs collected from worm uteri with eggs isolated from pig faeces. Acta Vet Scand. 1990, 31: 393-398.
Roepstorff A, Eriksen L, Slotved H-C, Nansen P: Experimental Ascaris suum infection in the pig: Worm populations kinetics following single inoculation with three doses of infective eggs. Parasitology. 1997, 115: 443-452. 10.1017/S0031182097001480.
Roepstorff A, Murrell KD: Transmission dynamics of helminth parasites of pigs on continuous pasture: Ascaris suum and Trichuris suis. Int J Parasitol. 1997, 27: 563-572. 10.1016/S0020-7519(97)00022-2.
Slotved H-C, Roepstorff A, Barnes EH, Eriksen L, Nansen P: Comparison of two methods for recovering migrating Ascaris suum larvae from the liver and lungs of pigs. J Parasitol. 1996, 82: 612-615. 10.2307/3283786.
Slotved H-C, Barnes EH, Eriksen L, Roepstorff A, Nansen P, Bjørn H: Use of an agar-gel technique for large scale application to recover Ascaris suum larvae from intestinal contents of pigs. Acta Vet Scand. 1997, 38: 207-212.
Slotved H-C, Eriksen L, Murrell KD, Nansen P: Comparison of methods for recovery of Ascaris suum larvae from tissues of mice. Int J Parasitol. 1997, 27: 1305-1310. 10.1016/S0020-7519(97)00101-X.
Slotved H-C, Roepstorff A, Barnes EH, Eriksen L, Nansen P: Comparison of two methods for recovering migrating Ascaris suum larvae from the liver and lungs of pigs. J Parasitol. 1996, 82: 612-615. 10.2307/3283786.
Van Wyk JA, Gerber HM, Groeneveld HT: A technique for the recovery of nematodes from ruminants by migration from gastro-intestinal ingesta gelled in agar: Large-scale application. Onderstepoort J Vet Res. 1980, 47: 147-158.
Wismer-Pedersen J, Møller AJ, Eriksen L, Nansen P, Roepstorff A: (Infection with Ascaris suum in swine liver). Dansk Vet Tidsskr. 1990, 73: 126-132.
Acknowledgements
We thank the Danish National Research Foundation for financial support.
Author information
Authors and Affiliations
Additional information
Reprints may be obtained from: Isam Saeed, Danish Centre For Experimental Parasitology, Royal Veterinary and Agricultural University, Dyrlaegevej 100, DK-1870 Frederiksberg C, Denmark. E-mail: iss@kvl.dk, tel: +45 35 28 27 93, fax: +45 35 28 27 74.
Rights and permissions
About this article
Cite this article
Saeed, I., Roepstorff, A., Rasmussen, T. et al. Optimization of the Agar-gel Method for Isolation of Migrating Ascaris suumLarvae From the Liver and Lungs of Pigs. Acta Vet Scand 42, 279 (2001). https://doi.org/10.1186/1751-0147-42-279
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1751-0147-42-279