
Abstract
Herbal medicine is one of the forms of traditional medical practice. Traditional Chinese medicine (TCM) and traditional Vietnamese medicine (TVM) are well-known for their long-standing tradition of herbal medicine.
Secreted by many species of blister beetle, most notably by the 'Spanish fly' (Lytta vesicatoria), cantharidin inhibits protein phosphatases 1 and 2A (PP1, PP2A). Blister beetle has been used in Asian traditional medicine to treat Molluscum contagiosum virus (MCV) infections and associated warts, and is now also used for cancer treatment. A combination of both genomic and postgenomic techniques was used in our studies to identify candidate genes affecting sensitivity or resistance to cantharidin. Cantharidin was not found to be related to multidrug resistance phenotype, suggesting its potential usefulness for the treatment of refractory tumors. Oxidative stress response genes diminish the activity of cantharidin by inducing DNA strand breaks which may be subject to base excision repair and induce apoptosis in a p53- and Bcl2-dependent manner.
Cantharidin is one of many natural products used in traditional Chinese medicine and traditional Vietnamese medicine for cancer treatment. Combined methods of pharmaceutical biology and molecular biology can help elucidate modes of action of these natural products.
Similar content being viewed by others


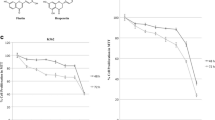
Background
Herbal medicine represents a traditional form of medical practice in human history. Current ethnobotany and ethnopharmacology focus on the systematic exploration of medicinal herbs among folk medicines [1].
Half a century after the launch of chemotherapy for tumor treatment [2], anti-neoplastic drugs is now indispensable for treating hematopoietic malignancies. The concept of combination therapy in oncology is based on the notion that leukemia cells resistant to one drug may be susceptible to other drugs. Clinically, the success of combination treatments can be hampered by the development of broad-spectrum or multidrug resistance (MDR). Two strategies to cope with this problem are to modulate multidrug resistance by inhibitors of MDR-conferring proteins [3] or to develop new anticancer drugs without involvement in MDR phenotypes. Enormous efforts have been spent to develop such as resistance modulation to improve treatment for leukemia, but resistance modulators still frequently show intolerable high toxicity [4]. Natural products provide a rich source for developing novel drugs with anti-leukemia activities. The Natural Products Branch of the National Cancer Institute (USA) has collected and tested over 100,000 natural extracts of plants and invertebrates [5]. The vast experience of traditional medicines (e.g., TCM) may facilitate the identification of novel active substances. This approach has been successful. Camptothecin from Camptotheca acuminata represents only one outstanding example for such compounds derived from TCM [6]. Considering the severe limitations of current cancer chemotherapy, it would be desirable to have novel drugs which are active against otherwise resistant tumor cells.
In 1996, we started a research program on the molecular pharmacology and pharmacogenomics of the natural products derived from TCM [7]. This project became fruitful for the identification and characterization of compounds with anti-tumor and anti-viral activities. Apart from artemisinin and its prominent semi-synthetic derivative artesunate which are both approved drugs [8, 9], we have analyzed cellular and molecular mechanisms of several other chemically characterized natural products derived from TCM, e.g., arsenic trioxide, ascaridol, berberine, cantharidin, cephalotaxine, curcumin, homoharringtonine, luteolin, isoscopoletin, scopoletin and others [10–30]. Furthermore, several novel bioactive compounds, namely tetracentronsine A, tetracentronside A, B and C, the two novel α-tetralone derivatives, berchemiaside A and B, and the novel flavonoid quercetin-3-O-(2-acetyl-α-L-arabinofuranosid), were described and analyzed in our investigations, [31–33]. Artemisinin displays a marked anti-malarial activity [34]. Various derivatives (artesunate, artemether, arteether, artelinate) have been synthesized to improve this anti-malarial activity.
In 2002, we began a study on the antiviral effects of artesunate. We demonstrated for the first time that artesunate inhibits NF-κB activity, leading to the inhibition of viral replication. NF-κB is involved in the transcriptional regulation of early and late proteins of human cytomegalovirus (HCMV) necessary for viral replication [35]. Artesunate also acts against cytomegaloviruses in vivo [36]. We also showed that the antiviral activity of artesunate is not limited to HCMV. Herpes simplex virus 1, hepatitis B and C viruses and others can also be inhibited by artemisinin and artesunate [35, 37, 38].
TCM is well-known for its unique diagnosis and treatment system, whereas other traditional medicines in Asia, such as TVM, have not gained the same acceptance in medical practice. Profoundly influenced by TCM, TVM exhibits its uniqueness via an influence from indigenous medicine of southern Vietnam. Detailed surveys of TVM and TCM can be found in recent publications [39–41].
Cantharidin
Screening of extracts
A major problem in conventional cancer chemotherapy is the severe side-effects on normal tissues prevent treatment with doses sufficient to kill all cancer cells, which in turn fosters the development of drug resistance. There is an urgent need to develop new drugs with improved resistance modulation for tumor therapy. It is the current situation in cancer chemotherapy in Western medicine that prompted us to study TCM and TVM. We have been searching for natural products from medicinal plants with activity against cancer cells.
In this study, we focused on medicinal plants and animals used in Vietnam. They were bought by of the authors at medicinal markets in Ho Chi Minh City, Vietnam (Figure 1). The criteria for medicinal plants and animals to be included in this study were their traditional use to treat cancer. The material was further processed by extraction with solvents of different polarity (petroleum ether, n-hexane, methanol, chloroform, ethyl acetate, or water). Extracts with organic solvents were made in a Soxhlet apparatus, while water extracts were made as decoctions imitating the original medicinal use. The aim of this approach was to divide plant constituents into fractions of different polarity upon extraction.

Medicinal plants and animals used in Vietnamese medicine and cytotoxicity of hexane, water or methanol extracts (10 μg/ml) to CCRF-CEM leukemia cells. (1) Lytta vesicatoria (whole beetles), (2) Pueralia lobata (roots), (3) Momordica charantia (roots, branches), (4) Momordica charantia (fruits), (5) Dichroa febrifuga (roots), (6) Panax ginseng C.A.Mey., (7) Curcuma longa (rhizoma), (8) Trichosanthes kirilowii Maxim (kernels), (9) Glycyrrhiza uralensis (roots). Growth inhibitory activity was measured using a growth inhibition assay [16].
Only two out of the 27 extracts, namely Curcuma longa and blister beetle ('Spanish fly', Lytta vesicatoria), reduced cell growth of human CCRF-CEM leukemia cells to below 20% of untreated controls (Figure 1). We further tested curcumin and cantharidin, the active ingredients of Curcuma longa and of Lytta vesicatoria, respectively [16].
Multidrug resistance
Parental CCRF-CEM leukemia cells and their multidrug-resistant sub-lines CEM/ADR5000, CEM/VLB100, and CEM/E1000 were used to investigate whether these two substances are able to inhibit tumor cell growth and to analyze the involvement of these compounds in multidrug resistance (MDR) phenotype. CEM/ADR5000 and CEMJ/VLB100 are characterized by over-expression of the MDR-mediating ATP-binding cassette (ABC) transporter P-glycoprotein (MDR1, ABCB1), while CEM/E1000 over-expresses the multidrug-resistance related protein 1 (MRP1, ABCC1) [42–44]. Cantharidin is more active than curcumin and no cross-resistance was observed in multidrug-resistant cell lines indicating that both drugs are not involved in the MDR phenotype (Table 1). Since cantharidin was active at lower concentrations compared to curcumin, we focused our efforts on cantharidin. An overview on the chemotherapeutic features of curcumin can be found in a recent publication [45].
Use of canthardin in Asian and Western medicine
Topical application of cantharidin has a long tradition in Asian medicine for the treatment of warts caused by Molluscum contagiosum virus (MCV) infections [46], while the use of cantharidin to treat pediatric MCV infections in Western academic medicine has been found effective [47, 48]. Blister beetles and cantharidin are also used in China and Vietnam to treat cancer [49]. Despite its usefulness, the potential poisonous effects of cantharidin would have fatal consequences in the event of careless mistakes in the use of this compound [50, 51]. Not only is exact dosage is required for the use of cantharidin itself, but also for raw preparations of blister beetles to be used in traditional medicines. Cantharidin content varies among individual blister beetles as well as among different species. Blister beetles belong to the order Coleoptera which comprises approximately 1500 species.
In this review, our data are compared with well-known results regarding the molecular and cellular mechanisms of cantharidin in cancer cells.
Pharmacogenomics of cantharidin
Cantharidin and norcantharidin (a demethylated cantharidin derivative, which also has clinical potential) are protein phosphatase 1 (PP1) and protein phosphatase 2A (PP2A) inhibitors [52–59]. This activity appears necessary for the growth inhibition activity of these compounds [57]. Protein phosphatases are involved, among others, in the regulation of multiple cellular processes including apoptosis, signal transduction pathways, cell cycle progression, glucose metabolism and calcium transport [60]. Thus, although the biochemical target of cantharidin and norcantharidin is known, the critical molecular pathways by which cantharidin and norcantharidin cause growth inhibition and cell death are unclear [61–63].
In an attempt to identify the molecular determinants that predict sensitivity or resistance of tumor cells to cantharidin, we analyzed the microarray database of the National Cancer Institute (USA). Out of 9706 genes, 21 genes whose mRNA expression in 60 tumor cell lines correlated with the highest correlation coefficients to inhibition concentration 50% (IC50) values were selected by COMPARE analysis and false discovery rate calculation [10]. These genes were subjected to hierarchical cluster analysis to reveal whether the expression profiles of these genes are useful to predict sensitivity or resistance of cell lines to cantharidin. The mRNA expression of the 21 identified genes were subjected to hierarchical cluster analysis and cluster image mapping (Figure 2). The resulting dendrogram with the genes analyzed on the right can be divided into three major branches. The dendrogram on the top shows the cell lines and can also be separated into three major branches. By generation of a cluster image map from both dendrograms, areas with different mRNA expression levels became apparent (Figure 2). The distribution of sensitive or resistant cell lines on the dendrogram was significantly different indicating that cellular response to cantharidin is predictable by these genes [10].

Dendrograms and cluster image map obtained by hierarchical cluster analysis (complete linkage method) of mRNA expression of 21 genes in 60 NCI cell lines. The genes are published [10]. The dendrogram on the right shows the clustering of cell lines and the dendrogram on the top shows clustering of genes. The cluster image map corresponds to each mRNA expression value obtained by microarray analysis. The expression values have been normalized and color-coded as indicated. The figure is taken from [10].
While the specific functions of the proteins encoded by the 21 identified genes are diverse, it is intriguing that many of them are in one way or another involved in DNA damage response, DNA repair, and/or apoptosis [10]. Since cantharidin is an inhibitor of protein phosphatases 1 (PP1) and 2A (PP2A), PPP1R13B, the regulatory subunit 13B of PP1, is particularly interesting. PPP1R13B plays a central role in the regulation of apoptosis via its interaction with the tumor suppressor gene p53. It regulates p53 by enhancing DNA binding activity and transactivation function of p53 on the promoters of proapoptotic genes in vivo [64]. The role of PP1 in the repair of UV-induced DNA lesions [65] and the induction of cytosine arabinoside-induced apoptosis have been shown [66]. It is, therefore, reasonable to hypothesize that PPP1R13B also has specific functions in cantharidin-induced DNA repair and apoptosis. PP1 is one of the four major serine/threonine protein phosphatases, and new protein phosphatases are still emerging. They are crucial regulators of many cellular functions by altering the phosphorylation of target proteins. The level of phosphorylation is a well-controlled balance by the opposing actions of protein kinases and protein phosphatases. The PP1 holoenzyme consists of catalytic and regulatory subunits. Four catalytic (a, b, c, d) and more than a dozen regulatory subunits have so far been identified. The regulatory subunits modulate the substrate specificity and target the holoenzyme to specific subcellular localizations.
Apoptosis and DNA damage and repair induced by cantharidin
The microarray analyses were used to find the genes responsible for the action of cantharidin. These studies showed that many apoptosis-related genes and genes involved in DNA damage and repair correlated with the IC50 values for cantharidin in the NCI cell line panel. We analyzed the relevance of apoptosis and DNA damage and repair induced by cantharidin in more detail. In a recent study, we reported that cantharidin induces apoptosis by a p53-dependent mechanism in leukemia cells [24]. Cantharidin causes both DNA single- and double-strand breaks. Colony forming assays with knockout and transfectant cell lines showed that DNA polymerase β conferred increased cell survival after cantharidin treatment, indicating that base excision repair rather than nucleotide excision repair is important for cantharidin-induced DNA lesions. Oxidative-stress resistant thymic lymphoma-derived WEHI7.2 variants are also more resistant to cantharidin. These data suggest that cantharidin treatment causes oxidative stress which damages DNA and triggers p53-dependent apoptosis.
It has been difficult to determine all critical events responsible for cantharidin-induced cytotoxicity. PP1 and PP2A which are inhibited by cantharidin [52–59], modulate a large number of cellular processes by counteracting the activity of kinases to provide the critical on/off switch for many pathways [60]. Cell cycle progression is one process where the increases and decreases of both kinases and phosphatases are necessary to complete the cycle. Studies from different laboratories agree that cantharidin and norcantharidin treatment results in a G2/M cell cycle block in many cell types [68–72]. However, several of these studies suggest that cell cycle blockade does not cause cantharidin-induced apoptosis [68–72].
Our data show that cantharidin treatment causes DNA strand breaks in CCRF-CEM cells [24]. DNA strand breaks have also been documented in oral cancer KB cells after norcantharidin treatment [73]. A correlation between an increase in the mRNA level for several DNA damage repair genes in the 60 cell line panel and resistance to cantharidin argues for DNA damage as a mechanistic component of cantharidin-induced apoptosis. This is consistent with the role of p53 in cantharidin-induced apoptosis in this study because one of the major functions of p53 is to induce apoptosis when DNA damage exceeds a threshold [74]. Moreover, p53 plays a role in norcantharidin-induced apoptosis in glioblastoma cells [68]. Phosphorylation of p53 stabilizes the protein [74, 75]; inhibition of phosphatases may enhance the ability of p53 to exert its effect. The ability of Bcl-2 to protect against cantharidin-induced apoptosis seen in this study indicates that DNA damage-triggered mitochondrial pathway is involved. Mitochondrial dysfunction and activation of caspases involved in the intrinsic (mitochondrial) pathway of apoptosis have also been detected in other cell types after cantharidin or norcantharidin treatment [69, 70, 76–83]. A role of the Fas/CD95 extrinsic pathway of apoptosis has been reported [84], but not confirmed by other authors [85].
The cross-resistance of the oxidative stress resistant WEHI7.2 variants to cantharidin suggests that cantharidin causes oxidative stress which plays a role in cantharidin-induced apoptosis. Analogs of cantharidin increase xanthine oxidase activity which would increase intracellular reactive oxygen species (ROS) [86]. It is, therefore, tempting to speculate that oxidative stress is involved in the induction of DNA damage by cantharidin. Increase of endogenous ROS level has repeatedly been shown to cause significant DNA breakage [87].
Resistance to oxidative stress, increases of Bcl-2, or the presence of wild type p53 have a modest effect on cantharidin-induced toxicity. Mutational inactivation of PolB but not of ERCC1, key enzymes of base excision repair and nucleotide excision repair pathways respectively, exerted an effect on cantharidin cytotoxicity. This suggests that cantharidin induces non-bulky DNA lesions that are repaired by base excision repair but not by nucleotide excision repair. Lesions induced by oxidative stress are repaired by base excision repair and non-homologous end joining [88]. This suggests the possibility that multiple mechanisms are responsible for cantharidin-induced toxicity. In this study, for example, p53 status affected the IC50 of cantharidin, however, cells with mutated p53 still died. A p53-independent mechanism of cantharidin-induced cytotoxicity has been detected in hepatoma cells [70]. Because cantharidin and norcantharidin inhibit phosphatases, it would not be surprising that alterations in multiple pathways are critical for apoptosis. The microarray data comparing cantharidin treated and untreated HL-60 cells suggest that cantharidin affects multiple pathways [89]. Given that multiple mechanisms are involved in cantharidin-induced toxicity, the drug will likely be most effective against cancer cells with a specific phenotype. It is important to test whether this is a cancer cell that has acquired mutations in DNA repair pathways but retains wild-type p53 and sensitivity to oxidative stress.
Discussion
Although the potential use of natural products is increasingly recognized in oncology, it has been estimated that so far only 5000 plant species have been properly studied for possible medical applications [90]. Considering that there are 250,000 to 300,000 plant species on this planet, the majority of this treasure still awaits retrieval.
The isolation of natural products and the elucidation of their chemical structures enable pharmacological and molecular biological investigations comparable to those conducted on chemically synthesized compounds to be conducted. The identification of target molecules relevant to diseases allows screening for natural products that are able to inhibit these targets. This may lay groundwork for the development of rational treatment of diseases such as cancer. This kind of research also opens avenues for the prediction of individual response of a cancer patient to therapy. We expect that strategies for individualized tumor therapies will lead to improved results for the patients. Small molecule inhibitors have the potential to increase tumor specificity and reduce adverse side effects on normal tissues. This concept of individualized tumor therapy is also of great importance for small molecule inhibitors derived from traditional herbal medicines such as TCM and TVM. Applying this strategy, we identified cantharidin as a potential drug candidate. These results are supported by findings that cantharidin does not cause myelosuppression in patients [70] and is effective against cells with a multidrug resistance phenotype [16, 91], both of which are major obstacles of established anti-cancer drugs. A drawback of cantharidin is its acute toxicity due to its effect on mucus membranes and the urinary tract [49, 73]. Understanding the mechanism of cantharidin action will help in the derivation of related compounds that have reduced toxicity while preserving the anti-tumor effect [92, 93].
Cantharidin's ability to act against multidrug-resistant cells makes it an ideal compound for individualized cancer treatment. If multidrug resistance of a tumor can be detected by molecular and pharmacological means prior to standard chemotherapy, the therapy regimen may be altered and drugs that act against multidrug-resistant tumors (e.g., cantharidin) may be applied. Such escape treatment strategies are promising.
Our pharmacogenomic approach points to genes involved in oxidative stress response, DNA repair and apoptosis. The generation of hypotheses by genomic technologies and their verification by molecular biological methods provides an attractive approach. On the other hand, microarray technologies alone are not sufficient to elucidate molecular mechanisms of cytotoxic compounds in cancer cells. Microarray expression profiling can deliver candidate genes on a transcriptome-wide level. While this approach is much faster than traditional techniques, not all associations of genes with the response of tumor cells to a drug under investigation are of causal relationships and some may be even unrelated processes. Therefore, we should only take validated findings seriously.
Conclusion
TCM and TVM are valuable sources for identifying potential natural products to treat cancer. In the case with cantharidin, the use of pharmacogenomic and molecular biological techniques help elucidate the modes of action of the natural product. While cantharidin is not involved in the multidrug resistance phenotype, it induces the generation of ROS and DNA damage, thereby leading to apoptosis.
References
Heinrich M, Bremner P: Ethnobotany and ethnopharmacy – their role for anti-cancer drug development. Curr Drug Targets. 2006, 7: 239-45. 10.2174/138945006776054988.
Rhoads CP: Report on a cooperative study of nitrogen mustard (HN2) therapy of neoplastic disease. Trans Assoc Am Physicians. 1947, 60: 110-117.
Tan B, Piwnica-Worms D, Ratner L: Multidrug resistance transporters and modulation. Curr Opin Oncol. 2000, 12: 450-458. 10.1097/00001622-200009000-00011.
Volm M, Pommerenke EW, Efferth T, Löhrke H, Mattern J: Circumvention of multidrug resistance in human kidney and kidney carcinoma in vitro. Cancer. 1991, 67: 2484-2489. 10.1002/1097-0142(19910515)67:10<2484::AID-CNCR2820671016>3.0.CO;2-I.
DTP – Natural Product Extract Cancer Screening Data. [http://dtp.nci.nih.gov/docs/cancer/natural_products/natural_products_data.html]
Liu LF, Desai SD, Li TK, Mao Y, Sun M, Sim SP: Mechanism of action of camptothecin. Ann NY Acad Sci. 2000, 922: 1-10.
Efferth T, Rücker G, Falkenberg M, Manns D, Olbrich A, Fabry U, Osieka R: Detection of apoptosis in KG-1a leukemic cells treated with investigational drugs. Arzneimittelforschung. 1996, 46: 196-200.
Efferth T: Mechanistic perspectives for 1,2,4-trioxanes in anti-cancer therapy. Drug Resist Updat. 2005, 8: 85-97. 10.1016/j.drup.2005.04.003.
Efferth T: Molecular pharmacology and pharmacogenomics of artemisinin and its derivatives in cancer cells. Curr Drug Target. 2006, 7: 407-421. 10.2174/138945006776359412.
Efferth T: Microarray-based prediction of cytotoxicity of tumor cells to cantharidin. Oncol Rep. 2005, 13: 459-463.
Efferth T, Kaina B: Microarray-based prediction of cytotoxicity of tumor cells to arsenic trioxide. Cancer Genomics Proteomics. 2004, 1: 363-370.
Efferth T, Oesch F: Oxidative stress response of tumor cells: microarray-based comparison between artemisinins and anthracyclines. Biochem Pharmacol. 2004, 68: 3-10. 10.1016/j.bcp.2004.03.003.
Efferth T, Volm M: Glutathione-related enzymes contribute to resistance of tumor cells and low toxicity in normal organs to artesunate. In Vivo. 2005, 19: 225-232.
Efferth T, Dunstan H, Sauerbrey A, Miyachi H, Chitambar CR: The anti-malarial artesunate is also active against cancer. Int J Oncol. 2001, 18: 767-773.
Efferth T, Olbrich A, Sauerbrey A, Ross DD, Gebhart E, Neugebauer M: Activity of ascaridol from the anthelmintic herb Chenopodium anthelminticum L. against sensitive and multidrug resistant tumor cells. Anticancer Res. 2002, 22: 4221-4224.
Efferth T, Davey M, Olbrich A, Rücker G, Gebhart E, Davey R: Activity of drugs from traditional Chinese medicine toward sensitive and MDR1- or MRP1-overexpressing multidrug-resistant human CCRF-CEM leukemia cells. Blood Cells Mol Dis. 2002, 28: 160-168. 10.1006/bcmd.2002.0492.
Efferth T, Olbrich A, Bauer R: mRNA expression profiles for the response of human tumor cell lines to the antimalarial drugs artesunate, arteether, and artemether. Biochem Pharmacol. 2002, 64: 617-623. 10.1016/S0006-2952(02)01221-2.
Efferth T, Sauerbrey A, Halatsch ME, Ross DD, Gebhart E: Molecular modes of action of cephalotaxine and homoharringtonine from the coniferous tree Cephalotaxus hainanensis in human tumor cell lines. Naunyn Schmiedebergs Arch Pharmacol. 2003, 367: 56-67. 10.1007/s00210-002-0632-0.
Efferth T, Sauerbrey A, Olbrich A, Gebhart E, Rauch P, Weber HO, Hengstler JG, Halatsch ME, Volm M, Tew KD, Ross DD, Funk JO: Molecular modes of action of artesunate in tumor cell lines. Mol Pharmacol. 2003, 64: 382-394. 10.1124/mol.64.2.382.
Efferth T, Briehl MM, Tome ME: Role of antioxidant genes for the activity of artesunate against tumor cells. Int J Oncol. 2003, 23: 1231-1235.
Efferth T, Benakis A, Romero MR, Tomicic M, Rauh R, Steinbach D, Hafer R, Stamminger T, Oesch F, Kaina B, Marschall M: Enhancement of cytotoxicity of artemisinins toward cancer cells by ferrous iron. Free Radic Biol Med. 2004, 37: 998-1009. 10.1016/j.freeradbiomed.2004.06.023.
Efferth T, Ramirez T, Gebhart E, Halatsch ME: Combination treatment of glioblastoma multiforme cell lines with the anti-malarial artesunate and the epidermal growth factor receptor tyrosine kinase inhibitor OSI-774. Biochem Pharmacol. 2004, 67: 1689-1700. 10.1016/j.bcp.2003.12.035.
Efferth T, Chen Z, Kaina B, Wang G: Molecular determinants of response of tumor cells to berberine. Cancer Genomics Proteomics. 2005, 2: 115-124.
Efferth T, Rauh R, Kahl S, Tomicic M, Bochzelt H, Tome ME, Briehl MM, Bauer R, Kaina B: Molecular modes of action of cantharidin in tumor cells. Biochem Pharmacol. 2005, 69: 811-818. 10.1016/j.bcp.2004.12.003.
Dell'Eva R, Pfeffer U, Vene R, Anfosso L, Forlani A, Albini A, Efferth T: Inhibition of angiogenesis in vivo and growth of Kaposi's sarcoma xenograft tumors by the anti-malarial artesunate. Biochem Pharmacol. 2004, 68: 2359-2366. 10.1016/j.bcp.2004.08.021.
Rinner B, Siegl V, Pürstner P, Efferth T, Brem B, Greger H, Pfragner R: Activity of novel plant extracts against medullary thyroid carcinoma cells. Anticancer Res. 2004, 24: 495-500.
Berger TG, Dieckmann D, Efferth T, Schultz ES, Funk JO, Baur A, Schuler G: Artesunate in the treatment of metastatic uveal melanoma – first experiences. Oncol Rep. 2005, 14: 1599-1603.
Adams M, Efferth T, Bauer R: Activity-guided isolation of scopoletin and isoscopoletin the inhibitory active principles towards CCRF-CEM leukaemia cells and multi-drug resistant CEM/ADR5000 cells from Artemisia argyi. Planta Med. 2006, 72: 862-864. 10.1055/s-2006-947165.
Anfosso L, Efferth T, Albini A, Pfeffer U: Microarray expression profiles of angiogenesis-related genes predict tumor cell response to artemisinins. Pharmacogenomics J. 2006, 6: 269-278.
Fu YJ, Efferth T, Zu YG: Optimization of luteolin separation from pigeonpea [Cajanus Cajan (L.) Millsp.] leaves by macroporous resins. J Chromatogr A. 2006, 1137: 145-152. 10.1016/j.chroma.2006.08.067.
Wang J, Zheng Y, Efferth T, Wang R, Shen Y, Hao X: Indole and carbazole alkaloids from Glycosmis montana with weak anti-HIV and cytotoxic activities. Phytochemistry. 2005, 66: 697-701. 10.1016/j.phytochem.2005.02.003.
Wang YF, Cao JX, Efferth T, Lai GF, Luo SD: Cytotoxic and new tetralone derivatives from Berchemia floribunda (Wall.) Brongn. Chem Biodivers. 2006, 3: 646-653. 10.1002/cbdv.200690067.
Wang YF, Lai GF, Efferth T, Cao JX, Luo SD: New glycosides from Tetracentron sinense and their cytotoxic activity. Chem Biodivers. 2006, 3: 1023-1030. 10.1002/cbdv.200690100.
Efferth T: Willmar Schwabe Award 2006: Antiplasmodial and anti-tumor activity of artemisinin – From the bench to the bedside. Planta Med. 2007, 73: 299-309. 10.1055/s-2007-967138.
Efferth T, Marschall M, Wang X, Huong SM, Hauber I, Olbrich A, Kronschnabl M, Stamminger T, Huang ES: Antiviral activity of artesunate towards wild-type, recombinant, and ganciclovir-resistant human cytomegaloviruses. J Mol Med. 2002, 80: 233-242. 10.1007/s00109-001-0300-8.
Kaptein SJ, Efferth T, Leis M, Rechter S, Auerochs S, Kalmer M, Bruggeman CA, Vink C, Stamminger T, Marschall M: The anti-malaria drug artesunate inhibits replication of cytomegalovirus in vitro and in vivo. Antiviral Res. 2006, 69: 60-69. 10.1016/j.antiviral.2005.10.003.
Romero MR, Efferth T, Serrano MA, Castano B, Macias RI, Briz O, Marin JJ: Effect of artemisinin/artesunate as inhibitors of hepatitis B virus production in an "in vitro" replicative system. Antiviral Res. 2005, 68: 75-83. 10.1016/j.antiviral.2005.07.005.
Romero MR, Serrano MA, Vallejo M, Efferth T, Alvarez M, Marin JJ: Antiviral effect of artemisinin from Artemisia annua against a model member of the Flaviviridae family, the bovine viral diarrhoea virus (BVDV). Planta Med. 2006, 72: 1169-1174. 10.1055/s-2006-947198.
Tang W, Hemm I, Bertram B: Recent development of antitumor agents from Chinese herbal medicines; part I. Low molecular compounds. Planta Med. 2003, 69: 97-108. 10.1055/s-2003-37718.
Tang W, Hemm I, Bertram B: Recent development of antitumor agents from Chinese herbal medicines. Part II. High molecular compounds (3). Planta Med. 2003, 69: 193-201. 10.1055/s-2003-38494.
Ngoc H, Borton L: Traditional medicine. 2005, The Gioi Publishers, Ha Noi
Beck WT, Mueller TJ, Tanzer LR: Altered surface membrane glycoproteins in Vinca alkaloid-resistant human leukemic lymphoblasts. Cancer Res. 1979, 39: 2070-2076.
Kimmig A, Gekeler V, Neumann M, Frese G, Handgretinger R, Kardos G, Diddens H, Niethammer D: Susceptibility of multidrug-resistant human leukemia cell lines to human interleukin 2-activated killer cells. Cancer Res. 1990, 50: 6793-6799.
Davey RA, Longhurst TJ, Davey MW, Belov L, Harvie RM, Hancox D, Wheeler H: Drug resistance mechanisms and MRP expression in response to epirubicin treatment in a human leukaemia cell line. Leuk Res. 1995, 19: 275-282. 10.1016/0145-2126(94)00159-8.
Singh RP, Agarwal R: Natural flavonoids targeting deregulated cell cycle progression in cancer cells. Curr Drug Targets. 2006, 7: 345-354. 10.2174/138945006776055004.
Moed L, Shwayder TA, Chang MW: Cantharidin revisited: a blistering defense of an ancient medicine. Arch Dermatol. 2001, 137: 1357-1360.
Silverberg N: Pediatric molluscum contagiosum: optimal treatment strategies. Paediatr Drugs. 2003, 5: 505-512. 10.2165/00148581-200305080-00001.
Smolinski KN, Yan AC: How and when to treat molluscum contagiosum and warts in children. Pediatr Ann. 2005, 34: 211-221.
Wang GS: Medical uses of mylabris in ancient China and recent studies. J Ethnopharmacol. 1989, 26: 147-162. 10.1016/0378-8741(89)90062-7.
Karras DJ, Farrell SE, Harrigan RA, Henretig FM, Gealt L: Poisoning from "Spanish fly" (canthardin). Am J Emerg Med. 1996, 14: 478-483. 10.1016/S0735-6757(96)90158-8.
Swingle M, Ni L, Honkanen RE: Small-molecule inhibitors of ser/thr protein phosphatases: specificity, use and common forms of abuse. Methods Mol Biol. 2006, 365: 23-38.
Li YM, Casida JE: Cantharidin-binding protein: identification as protein phosphatase 2A. Proc Natl Acad Sci USA. 1992, 89: 11867-11870. 10.1073/pnas.89.24.11867.
Honkanen RE: Cantharidin, another natural toxin that inhibits the activity of serine/threonine protein phosphatases types 1 and 2A. FEBS Lett. 1993, 330: 283-286. 10.1016/0014-5793(93)80889-3.
Eldridge R, Casida JE: Cantharidin effects on protein phosphatases and the phosphorylation state of phosphoproteins in mice. Toxicol Appl Pharmacol. 1995, 130: 95-100. 10.1006/taap.1995.1013.
Laidley CW, Cohen E, Casida JE: Protein phosphatase in neuroblastoma cells: [3H] cantharidin binding site in relation to cytotoxicity. J Pharmacol Exp Ther. 1997, 280: 1152-1158.
McCluskey A, Ackland SP, Gardiner E, Walkom CC, Sakoff JA: The inhibition of protein phosphatases 1 and 2A: a new target for rational anti-cancer drug design?. Anticancer Drug Des. 2001, 16: 291-303.
McCluskey A, Ackland SP, Bowyer MC, Baldwin ML, Garner J, Walkom CC, Sakoff JA: Cantharidin analogues: synthesis and evaluation of growth inhibition in a panel of selected tumour cell lines. Bioorg Chem. 2003, 31: 68-79. 10.1016/S0045-2068(02)00524-2.
To KK, Wang X, Yu CW, Ho YP, Au-Yeung SC: Protein phosphatase 2A inhibition and circumvention of cisplatin cross-resistance by novel TCM-platinum anticancer agents containing demethylcantharidin. Bioorg Med Chem. 2004, 12: 4565-4573. 10.1016/j.bmc.2004.07.009.
Shan HB, Cai YC, Liu Y, Zeng WN, Chen HX, Fan BT, Liu XH, Xu ZL, Wang B, Xian LJ: Cytotoxicity of cantharidin analogues targeting protein phosphatase 2A. Anticancer Drugs. 2006, 17: 905-911. 10.1097/01.cad.0000217428.90325.35.
Wera S, Hemmings BA: Serine/threonine protein phosphatases. Biochem J. 1995, 311: 17-29.
Wu LT, Chung JG, Chen JC, Tsauer W: Effect of norcantharidin on N-acetyltransferase activity in HepG2 cells. Am J Chin Med. 2001, 29: 161-172. 10.1142/S0192415X01000186.
Chen YJ, Shieh CJ, Tsai TH, Kuo CD, Ho LT, Liu TY, Liao HF: Inhibitory effect of norcantharidin, a derivative compound from blister beetles, on tumor invasion and metastasis in CT26 colorectal adenocarcinoma cells. Anticancer Drugs. 2005, 16: 293-299. 10.1097/00001813-200503000-00008.
Huan SK, Lee HH, Liu DZ, Wu CC, Wang CC: Cantharidin-induced cytotoxicity and cyclooxygenase 2 expression in human bladder carcinoma cell line. Toxicology. 2006, 223: 136-143. 10.1016/j.tox.2006.03.012.
Samuels-Lev Y, O'Connor DJ, Bergamaschi D, Trigiante G, Hsieh JK, Zhong S, Campargue I, Naumovski L, Crook T, Lu X: ASPP proteins specifically stimulate the apoptotic function of p53. Mol Cell. 2001, 8: 781-794. 10.1016/S1097-2765(01)00367-7.
Herman M, Ori Y, Chagnac A, Weinstein T, Korzets A, Zevin D, Malachi T, Gafter U: DNA repair in mononuclear cells: role of serine/threonine phosphatases. J Lab Clin Med. 2002, 140: 255-262. 10.1067/mlc.2002.127738.
Wang RH, Liu CW, Avramis VI, Berndt N: Protein phosphatase 1alpha-mediated stimulation of apoptosis is associated with dephosphorylation of the retinoblastoma protein. Oncogene. 2001, 20: 6111-6122. 10.1038/sj.onc.1204829.
Liu XH, Blazsek I, Comisso M, Legras S, Marion S, Quittet P, Anjo A, Wang GS, Misset JL: Effects of norcantharidin, a protein phosphatase type-2A inhibitor, on the growth of normal and malignant haemopoietic cells. Eur J Cancer. 1995, 31A: 953-963. 10.1016/0959-8049(95)00050-X.
Hong CY, Huang SC, Lin SK, Lee JJ, Chueh LL, Lee CH, Lin JH, Hsiao M: Norcantharidin-induced post-G(2)/M apoptosis is dependent on wild-type p53 gene. Biochem Biophys Res Commun. 2000, 276: 278-285. 10.1006/bbrc.2000.3341.
Wang CC, Wu CH, Hsieh KJ, Yen KY, Yang LL: Cytotoxic effects of cantharidin on the growth of normal and carcinoma cells. Toxicology. 2000, 147: 77-87. 10.1016/S0300-483X(00)00185-2.
Chen YN, Chen JC, Yin SC, Wang GS, Tsauer W, Hsu SF, Hsu SL: Effector mechanisms of norcantharidin-induced mitotic arrest and apoptosis in human hepatoma cells. Int J Cancer. 2002, 100: 158-165. 10.1002/ijc.10479.
Sakoff JA, Ackland SP, Baldwin ML, Keane MA, McCluskey A: Anticancer activity and protein phosphatase 1 and 2A inhibition of a new generation of cantharidin analogues. Invest New Drugs. 2002, 20: 1-11. 10.1023/A:1014460818734.
Bonness K, Aragon IV, Rutland B, Ofori-Acquah S, Dean NM, Honkanen RE: Cantharidin-induced mitotic arrest is associated with the formation of aberrant mitotic spindles and lagging chromosomes resulting, in part, from the suppression of PP2Aalpha. Mol Cancer Ther. 2006, 5: 2727-2736. 10.1158/1535-7163.MCT-06-0273.
Kok SH, Hong CY, Kuo MY, Lee CH, Lee JJ, Lou IU, Lee MS, Hsiao M, Lin SK: Comparisons of norcantharidin cytotoxic effects on oral cancer cells and normal buccal keratinocytes. Oral Oncol. 2003, 39: 19-26. 10.1016/S1368-8375(01)00129-4.
Norbury CJ, Zhivotovsky B: DNA damage-induced apoptosis. Oncogene. 2004, 23: 2797-2808. 10.1038/sj.onc.1207532.
Slee EA, O'Connor DJ, Lu X: To die or not to die: how does p53 decide?. Oncogene. 2004, 23: 2809-2818. 10.1038/sj.onc.1207516.
Morana SJ, Wolf CM, Li J, Reynolds JE, Brown MK, Eastman A: The involvement of protein phosphatases in the activation of ICE/CED-3 protease, intracellular acidification, DNA digestion, and apoptosis. J Biol Chem. 1996, 271: 18263-18271. 10.1074/jbc.271.30.18263.
Huh JE, Kang KS, Chae C, Kim HM, Ahn KS, Kim SH: Roles of p38 and JNK mitogen-activated protein kinase pathways during cantharidin-induced apoptosis in U937 cells. Biochem Pharmacol. 2004, 67: 1811-1818. 10.1016/j.bcp.2003.12.025.
An WW, Wang MW, Tashiro S, Onodera S, Ikejima T: Norcantharidin induces human melanoma A375-S2 cell apoptosis through mitochondrial and caspase pathways. J Korean Med Sci. 2004, 19: 560-566.
Kok SH, Chui CH, Lam WS, Chen J, Tang JC, Lau FY, Cheng GY, Wong RS, Chan AS: Induction of apoptosis on carcinoma cells by two synthetic cantharidin analogues. Int J Mol Med. 2006, 17: 151-157.
Li JL, Cai YC, Liu XH, Xian LJ: Norcantharidin inhibits DNA replication and induces apoptosis with the cleavage of initiation protein Cdc6 in HL-60 cells. Anticancer Drugs. 2006, 17: 307-314. 10.1097/00001813-200603000-00009. Erratum in: Anticancer Drugs 2006, 17:1107
Kok SH, Chui CH, Lam WS, Chen J, Lau FY, Cheng GY, Wong RS, Lai PP, Leung TW, Tang JC, Chan AS: Apoptotic activity of a novel synthetic cantharidin analogue on hepatoma cell lines. Int J Mol Med. 2006, 17: 945-949.
Kok SH, Chui CH, Lam WS, Chen J, Lau FY, Wong RS, Cheng GY, Tang WK, Cheng CH, Tang JC, Chan AS: Mechanistic insight into a novel synthetic cantharidin analogue in a leukaemia model. Int J Mol Med. 2006, 18: 375-379.
Kok SH, Chui CH, Lam WS, Chen J, Lau FY, Wong RS, Cheng GY, Tang WK, Teo IT, Cheung F, Cheng CH, Chan AS, Tang JC: Apoptogenic activity of a synthetic cantharimide in leukaemia: implication on its structural activity relationship. Int J Mol Med. 2006, 18: 1217-1221.
Peng F, Wei YQ, Tian L, Yang L, Zhao X, Lu Y, Mao YQ, Kan B, Lei S, Wang GS, Jiang Y, Wang QR, Luo F, Zou LQ, Liu JY: Induction of apoptosis by norcantharidin in human colorectal carcinoma cell lines: involvement of the CD95 receptor/ligand. J Cancer Res Clin Oncol. 2002, 128: 223-230. 10.1007/s00432-002-0326-5.
Kok SH, Cheng SJ, Hong CY, Lee JJ, Lin SK, Kuo YS, Chiang CP, Kuo MY: Norcantharidin-induced apoptosis in oral cancer cells is associated with an increase of proapoptotic to antiapoptotic protein ratio. Cancer Lett. 2005, 217: 43-52. 10.1016/j.canlet.2004.07.045.
Tsauer W, Lin JG, Lin PY, Hsu FL, Chiang HC: The effects of cantharidin analogues on xanthine oxidase. Anticancer Res. 1997, 17: 2095-2098.
Slupphaug G, Kavli B, Krokan HE: The interacting pathways for prevention and repair of oxidative DNA damage. Mutat Res. 2003, 531: 231-251.
Christmann M, Tomicic MT, Roos WP, Kaina B: Mechanisms of human DNA repair: an update. Toxicology. 2003, 193: 3-34. 10.1016/S0300-483X(03)00287-7.
Zhang JP, Ying K, Xiao ZY, Zhou B, Huang QS, Wu HM, Yin M, Xie Y, Mao YM, Rui YC: Analysis of gene expression profiles in human HL-60 cell exposed to cantharidin using cDNA microarray. Int J Cancer. 2004, 108: 212-218. 10.1002/ijc.11405.
Abelson PH: Medicine from plants. Science. 1990, 247 (4942): 513-10.1126/science.2300807.
Yi SN, Wass J, Vincent P, Iland H: Inhibitory effect of norcantharidin on K562 human myeloid leukemia cells in vitro. Leuk Res. 1991, 15: 883-886. 10.1016/0145-2126(91)90163-N.
Mazars G, Nastari-Micheli E, Mazars C: Recherches ethnopharmacologiques sur les insectes et autres arthropodes : l'intérêt de la pharmacopée chinoise traditionnelle. Ethnopharmacologia. 2004
Nakatani T, Konishi T, Miyahara K, Noda N: Three novel cantharidin-related compounds from the Chinese blister beetle, Mylabris phalerata Pall. Chem Pharm Bull (Tokyo). 2004, 52: 807-809. 10.1248/cpb.52.807.
Acknowledgements
We are indebted to Dr Van Tre Tran of the Institute of Traditional Medicine, Ho Chi Minh City, Vietnam for his assignment of scientific Latin names to the herbs and animals and Duy Hung Nguyen for his guidance and help in collecting the herbs in medicinal markets in Ho Chi Minh City, Vietnam. We thank Prof Robert Wyn Owen for his critical reading of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author(s) declare that they have no competing interests.
Authors' contributions
RR carried out the biological assays. HB coordinated the extraction of medicinal plants and animals and revised the manuscript. RB was the academic supervisor of SK. SK generated the extracts from medicinal plants and animals in RB's lab. BK coordinated the conduct of experiments of RR concerning apoptosis and DNA damage and repair. RR carried out the biological experiments in BK's lab. TE conceived the study, bought the medicinal plants and animals from Vietnam and drafted the manuscript. TE was the academic supervisor of RR. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Rauh, R., Kahl, S., Boechzelt, H. et al. Molecular biology of cantharidin in cancer cells. Chin Med 2, 8 (2007). https://doi.org/10.1186/1749-8546-2-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1749-8546-2-8