
Abstract
Activation of spinal microglia contributes to aberrant pain responses associated with neuropathic pain states. Endocannabinoids (ECs) are present in the spinal cord, and inhibit nociceptive processing; levels of ECs may be altered by microglia which modulate the turnover of endocannabinoids in vitro. Here, we investigate the effect of minocycline, an inhibitor of activated microglia, on levels of the endocannabinoids anandamide and 2-arachidonoylglycerol (2-AG), and the related compound N-palmitoylethanolamine (PEA), in neuropathic spinal cord. Selective spinal nerve ligation (SNL) in rats resulted in mechanical allodynia and the presence of activated microglia in the ipsilateral spinal cord. Chronic daily treatment with minocycline (30 mg/kg, ip for 14 days) significantly reduced the development of mechanical allodynia at days 5, 10 and 14 post-SNL surgery, compared to vehicle-treated SNL rats (P < 0.001). Minocycline treatment also significantly attenuated OX-42 immunoreactivity, a marker of activated microglia, in the ipsilateral (P < 0.001) and contralateral (P < 0.01) spinal cord of SNL rats, compared to vehicle controls. Minocycline treatment significantly (P < 0.01) decreased levels of 2-AG and significantly (P < 0.01) increased levels of PEA in the ipsilateral spinal cord of SNL rats, compared to the contralateral spinal cord. Thus, activation of microglia affects spinal levels of endocannabinoids and related compounds in neuropathic pain states.
Similar content being viewed by others
Introduction
Since the discovery of the cannabinoid receptors CB1 and CB2, and their endogenous ligands, there has been a rapid growth of evidence that the cannabinoid receptor system modulates the pathogenesis of various pain states. CB1 receptor agonists produce analgesic effects in models of neuropathic pain that are mediated by both peripheral and central sites of action [1]. However, activation of CB1 receptors in the central nervous system (CNS) is also associated with adverse psychoactivity [2].
Neuropathic pain responses are also attenuated by CB2 receptor agonists [3–5]. In contrast to the ubiquitous analgesic effects of CB1 receptor agonists, spinal administration of CB2 receptor agonists only has inhibitory effects in neuropathic rats and not sham-operated rats [6]. Importantly, the spinal effects of CB2 agonists in neuropathic mice are absent in CB2 knockout mice [4]. These functional data are supported by studies demonstrating an up-regulation of CB2 receptor mRNA and/or protein in the ipsilateral spinal cord [7–9] and ipsilateral dorsal horn [10] of neuropathic rats, compared to sham-operated rats. CB2 receptors in the spinal cord were co-localised with activated microglia in neuropathic rats [9], which is in keeping with earlier reports of CB2 receptor expression in microglia [11, 12].
It is well established that spinal microglia undergo various stages of morphological and immunophenotypic change following nerve injury and contribute to the development of neuropathic pain states [13]. Indeed, single injection of activated microglia into the spinal cord produces thermal hyperalgesia in mice, one of the symptoms of neuropathic pain [14]. The tetracycline derivative minocycline is neuroprotective in models of traumatic brain injury [15], Parkinson's disease and Alzheimer's disease [16]; effects which are largely attributed to inhibition of microglia activation. Up-regulation of activated microglia in the spinal cord of neuropathic rats contributes to aberrant pain behaviour and plays an important role in central sensitization. Minocycline attenuates neuropathic pain behaviour and reduces microglia activation in the spinal cord [17–20]. In vitro, microglia are capable of synthesising and catabolising endocannabinoids [21, 22], which play an important role in modulating nociceptive responses. Thus, we hypothesized that the presence of activated microglia in the spinal cord of neuropathic rats may contribute to the reported increased levels of the endocannabinoids anandamide (AEA) and 2-arachidonoylglycerol (2-AG) in the spinal cord [23]. Herein, we demonstrate that the inhibition of pain behaviour and microglia activation by chronic treatment with minocycline was associated with decreased spinal levels of 2-AG, but not AEA. By contrast, we report a significant elevation of PEA in the spinal cord of neuropathic rats treated with minocycline and demonstrate differing rates of hydrolysis of AEA and PEA by microglia BV-2 cells, which supports the notion that microglia differentially modulate levels of endocannabinoids in pathological states. Our data implicate a contribution of PEA, which has been shown to be neuroprotective in a model of focal cerebral ischemia [24], to the analgesic effects of minocycline in neuropathic rats.
Results
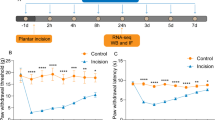
Minocycline attenuates mechanical allodynia induced by spinal nerve ligation
Chronic vehicle treatment did not alter the ipsilateral or contralateral paw withdrawal thresholds (PWTs) in sham-operated rats (data not shown). By contrast, SNL rats chronically treated with vehicle had a significantly (P < 0.001) lower ipsilateral PWT, compared to the PWT of the contralateral hindpaw of SNL rats and ipsilateral hindpaw of sham-operated rats (data not shown), consistent with the development of mechanical allodynia in these rats (Fig 1). Chronic daily treatment with minocycline (30 mg/kg, ip for 14 days) starting at 1 hr before SNL surgery significantly reduced the development of mechanical allodynia at days 5, 10 and 14 post-surgery compared to vehicle-treated SNL rats (Fig 1). Chronic minocycline treatment (30 mg/kg, ip) in sham-operated rats did not alter ipsilateral or contralateral PWT (data not shown).

Chronic treatment with minocycline significantly attenuates spinal nerve ligation-induced reduction in mechanical paw withdrawal threshold. Minocycline (Mino, 30 mg/kg, ip) or vehicle (Veh, sterile water) was injected daily for 14 days and paw withdrawal threshold of the ipsilateral (Ipsi) and contralateral (Contra) hindpaw were measured using von Frey filaments (sham data not shown for clarity of figure). Data were analysed using 2-way ANOVA followed by Bonferroni's post-hoc tests, and are expressed as mean (± SEM, n = 6 rats/group). ** P < 0.01, *** P < 0.001 vs contralateral paw; δ P < 0.05, δδ P < 0.01, δδδ P < 0.001 vs vehicle-ipsilateral paw.
Effects of chronic minocycline treatment on spinal glial cell proliferation
SNL surgery significantly increased OX-42 immunoreactivity in the L4-L6 sections of ipsilateral spinal cord of chronic vehicle-treated rats, compared to the contralateral spinal cord, indicative of spinal microglia activation (Fig 2, OX-42 immunoreactivity, ipsilateral-vehicle, 30.2 ± 0.5; contralateral-vehicle, 17.0 ± 0.3). Chronic minocycline treatment (30 mg/kg, ip) significantly attenuated OX-42 immunoreactivity in the ipsilateral (P < 0.001) and contralateral (P < 0.01) spinal cord of SNL rats, compared to vehicle controls (Fig 2). Thus, effects of minocycline treatment on neuropathic pain behaviour were associated with decreased levels of a marker for activated microglia in the ipsilateral spinal cord of SNL rats. Nevertheless, levels of OX-42 in the ipsilateral spinal cord of SNL rats treated with minocycline were still elevated compared to the contralateral spinal cord of minocycline-treated SNL rats (total: ipsilateral-minocycline, 14.9 ± 1.3; contralateral-minocycline, 6.5 ± 0.7).

Chronic minocycline treatment attenuates the spinal nerve ligation-induced increase in OX-42 immunoreactivity (ir) in the lumbar spinal cord. Vehicle (A, B, C, sterile water) or minocycline (D, E, F, 30 mg/kg, ip) was injected daily for 14 days and spinal cords dissected and processed for immunohistochemical detection of OX-42ir at the level of L4 (A-a" and D-d"), L5 (B-b" and E-e") and L6 (C-c" and F-f"). High magnification images of the dorsal area used for OX-42ir signal quantification are also shown. Histograms (G-I) showing effects of minocycline or vehicle in L4-L6 spinal cord OX-42ir (white bars: ipsilateral side; black bars: contralateral side) are shown at the bottom. Data are expressed as mean ± SEM (n = 12 sections from 4 rats per group). Data were analysed using Kruskal Wallis non-parametric test followed by Dunn's multiple comparison posthoc analysis, * P < 0.05, ** P < 0.01, *** P < 0.001 vs ipsilateral-veh, # P < 0.05, ## P < 0.01 vs contralateral-vehicle. Scale bars: F = 1 mm (applies to A-F); f' = 100 μm (applies to a'-f' and a"-f").
Minocycline treatment alters spinal levels of endocannabinoids and related fatty acid ethanolamides in spinal nerve ligated rats
In SNL rats with chronic vehicle treatment, levels of AEA were significantly increased (P < 0.01) in the ipsilateral spinal cord, compared to the contralateral spinal cord (Table 1). Following chronic minocycline treatment in SNL rats, levels of AEA in the ipsilateral spinal cord were also elevated, compared to the contralateral spinal cord (Fig 3). The elevated ipsilateral levels of AEA in minocycline-treated rats were not different from the levels in vehicle-treated SNL rats.

Effects of chronic intraperitoneal dosing of minocycline (Mino, 30 mg/kg) or vehicle (Veh, sterile water) on the levels of endocannabinoids (A, B) and related fatty acid ethanolamides (C, D) in the ipsilateral spinal cord of spinal nerve ligated (SNL) or sham-operated rats. Data were analysed using Mann-Whitney nonparametric test and are expressed as a mean percentage of contralateral levels ± SEM (n = 5 – 6 rats/group). * P < 0.05, ** P < 0.01 vs contralateral paw. AEA, anandamide; 2-AG, 2-arachidonoylglycerol; PEA, N-palmitoylethanolamine; OEA, N-oleoylethanolamine.
There were no significant differences in levels of OEA in the ipsilateral and contralateral spinal cord of vehicle-treated SNL rats (Table 1). Levels of OEA in ipsilateral and contralateral spinal cord of SNL rats were unaltered by minocycline treatment (Fig 3).
By contrast, chronic minocycline treatment of SNL rats had a marked impact on levels of 2-AG and PEA in the ipsilateral spinal cord, compared to the contralateral spinal cord (Fig. 3) and the ipsilateral spinal cord from vehicle-treated animals. In vehicle-treated SNL rats, levels of PEA were decreased (P < 0.01) in the ipsilateral spinal cord, compared to contralateral spinal cord (Table 1). However levels of PEA were increased (P < 0.01) in the ipsilateral spinal cord (31.59 ± 5.61 nmol/g) of minocycline-treated SNL rats compared to their contralateral spinal cord (5.01 ± 0.48 nmol/g) (Fig 3). Although there were no significant differences in levels of 2-AG in the ipsilateral and contralateral spinal cord of vehicle-treated SNL rats (Table 1), levels of 2-AG were significantly decreased (P < 0.01) in the ipsilateral spinal cord (4.53 ± 0.93 nmol/g) compared to contralateral spinal cord (47.66 ± 4.38 nmol/g) of minocycline treated SNL rats (Fig 3).
Chronic minocycline treatment of sham-operated rats did not significantly alter levels of AEA, OEA or PEA in the ipsilateral spinal cord, compared to the contralateral spinal cord (Fig 3). By contrast, levels of 2-AG in the ipsilateral spinal cord of minocycline treated sham-operated rats were significantly (P < 0.05) lower than levels in the contralateral spinal cord (Fig 3; ipsilateral: 39.79 ± 4.38 nmol/g, contralateral: 62.61 ± 7.91 nmol/g).
Levels of ECs and related compounds in the contralateral spinal cord of SNL rats treated with minocycline were compared to levels in the contralateral spinal cord of SNL rats treated with vehicle. Although levels of AEA in the contralateral spinal cord of minocycline treated SNL rats were not significantly different to levels in the contralateral spinal cord of vehicle-treated SNL rats, there was a trend towards an increase in the minocycline treated group (53.72 ± 2.33 pmol/g vs 28.42 ± 0.86 pmol/g respectively). Levels of OEA in the contralateral spinal cord of minocycline-treated rats were comparable to levels in the contralateral spinal cord of vehicle-treated rats (1.51 ± 0.13 nmol/g vs 1.43 ± 0.08 nmol/g). In contrast to the marked effects of minocycline treatment on levels of PEA and 2-AG in the ipsilateral spinal cord of SNL rats, this treatment did not significantly influence levels of PEA and 2AG in the contralateral spinal cord. There were no significant differences between levels of 2-AG or PEA in contralateral spinal cord of minocycline-treated SNL rats compared to vehicle-treated SNL rats (2-AG, 47.66 ± 4.38 nmol/g vs 54.40 ± 4.02 nmol/g; PEA, 5.01 ± 0.48 nmol/g vs 8.80 ± 0.97 nmol/g).
Microglial differentially hydrolyse AEA and PEA
Our in vivo data demonstrate that neuropathic pain has a differential effect on the levels of AEA and PEA in the spinal cord, which may reflect the presence of microglia in the spinal cord. Here, we investigated the hypothesis that microglia have a differential effect on the hydrolysis of AEA and PEA, which may account for our in vivo observations in neuropathic rats. We report marked differences in the time courses for PEA and AEA hydrolysis in intact BV-2 microglial cells (Fig 4.) Specifically, the capacity for hydrolysis of AEA by the BV-2 cells was limited, in that, after 10 minutes, there was no further metabolism of AEA, as shown by the plateau in accumulation of water-soluble product. In marked contrast, the hydrolysis of PEA continued for at least 30 minutes (Fig 4).

Time course of hydrolysis of AEA (anandamide) and PEA (N-palmitoylethanolamine) in BV2 microglial cells. Data are means ± SEM of six independent experiments conducted in triplicate.
Discussion
Two weeks following peripheral nerve injury, levels of the endocannabinoid AEA are markedly elevated in the ipsilateral spinal cord of neuropathic rats, whereas levels of the related anti-inflammatory compound PEA are significantly decreased. By contrast levels of 2-AG and OEA in the ipsilateral spinal cord of neuropathic rats are unaltered compared to the contralateral spinal cord. Inhibition of microglia activation, by chronic minocycline treatment, significantly altered pain behaviour and levels of 2-AG and PEA in the ipsilateral spinal cord of neuropathic rats without affecting levels of AEA or OEA.
Two weeks following peripheral nerve injury we observed a significant elevation in levels of activated microglia (increased OX-42 labelling) and AEA in the ipsilateral spinal cord, compared to the contralateral spinal cord. By contrast, levels of AEA in the ipsilateral spinal cord of sham operated rats were unaltered compared to the contralateral spinal cord. In the same group of neuropathic rats, levels of PEA were significantly decreased in the ipsilateral spinal cord, compared to the contralateral spinal cord. Levels of PEA were unaltered in the ipsilateral spinal cord of sham operated rats, compared to the contralateral spinal cord. The elevation in levels of AEA in neuropathic rats is consistent with an earlier report that levels of AEA are elevated 3 fold in the whole lumbar spinal cord at 3 and 7 days following chronic constriction injury of the sciatic nerve [23]. This previous study also reported decreased levels of PEA in the whole lumbar spinal cord at 3 days, but not 7 days, following chronic constriction injury of the sciatic nerve. Although our findings are consistent with the previous report of increased levels of AEA in the lumbar spinal cord, our data highlight the importance of selective ipsilateral changes in levels of ECs and related compounds which is not unexpected given the unilateral nature of the pain behaviour.
Our data demonstrating a differential effect of peripheral nerve injury on levels of AEA versus PEA in the ipsilateral spinal cord suggest that their metabolism can be regulated independently in vivo, in keeping with the discovery of different biosynthetic pathways for AEA and PEA in the brain [25] and evidence of independent signalling pathways in a microglia cell line (BV-2 cells) [26].
Despite the increased levels of activated microglia in the ipsilateral spinal cord, compared to the contralateral spinal cord, of neuropathic rats we did not observe any changes in levels of 2-AG in the ipsilateral spinal cord of neuropathic rats. In vitro studies have shown a role of 2-AG in CB2 receptor mediated migration of microglia [27] and proliferation of microglia [21]. In the present study we did not determine whether levels of 2-AG were elevated at earlier timepoints, which may have contributed to the recruitment of microglia to the site of spinal injury. This does, however, seem unlikely as levels of 2-AG were unaltered in the whole lumbar spinal cord at 3 days following peripheral nerve injury [23].
Chronic daily treatment with minocycline significantly attenuated the development of mechanical allodynia and the associated increase in activated microglia, in the L4 - L6 region of the ipsilateral spinal cord in neuropathic rats. These data are consistent with previous studies of the effects of minocycline on microglia activation and neuropathic pain behaviour [17, 28]. Levels of OX-42 labelling in the contralateral spinal cord of vehicle-treated SNL rats were higher than those in the contralateral spinal cord of minocycline-treated SNL rats, consistent with the bilateral activation of microglia in the spinal cord following peripheral nerve injury [29] and indicates that minocycline treatment modulates this bilateral activation of microglia. Nevertheless, we did not observe any changes in paw withdrawal threshold of the contralateral hindpaw which indicates the absence of mechanical allodynia. Minocycline treatment did not significantly alter mechanical allodynia until day 5 post SNL surgery, there was however a trend towards an effect on day 3. These data corroborate previous studies reporting that the proliferation of microglia peaks at around 3 days following nerve injury, however this does vary slightly depending on the model studied [30]. The lack of effect of minocycline on mechanical allodynia on the first day after SNL surgery may reflect a delayed onset of action of minocycline [31]. Alternatively, microglia may not make a substantial contribution to the mechanical allodynia at this very early timepoint and, therefore, this treatment is unable to alter responses. It is feasible that the mechanical allodynia observed at day 1, and to some extent day 3, following SNL surgery is mediated, at least in part, by a transient post-surgical pain.
The effects of minocycline have been largely attributed to the inhibition of microglia [32], but effects on neuronal function in vitro should be borne in mind [31]. The integrated nature of in vivo studies makes it difficult to dissociate direct, versus indirect, effects of minocycline on microglia function and how this relates to levels of ECs and related compounds in neuropathic rats. It is estimated that, in vitro at least, microglia produce 20-fold more endocannabinoids than neurones or astrocytes [27, 33] and primary microglia inactivate both AEA and 2-AG [34], suggesting that changes in the levels of activated microglia in the spinal cord, despite their lower abundance compared with neurones and other glia, could have a significant effect on whole tissue endocannabinoid content. Our data are consistent with this notion, indeed inhibition of microglia activation was associated with a significant decrease in levels of 2-AG in the ipsilateral spinal cord of neuropathic rats, compared to the contralateral spinal cord. These data suggest that activated microglia may contribute to the biosynthesis of 2-AG in neuropathic rats, which is consistent with previously described production of 2-AG by microglia in vitro [21, 27, 34], and provide indirect evidence for microglia modulation of 2-AG turnover in vivo.
We have identified major differences in the effects of microglial inhibition on levels of AEA and PEA in neuropathic rats, which suggest that there is divergence in the biosynthesis/catabolism of these N-acylethanolamines by microglia. Our data suggest that activated microglia play a major role in modulating levels of PEA, but not AEA or OEA, in the spinal cord of neuropathic rats. Given that in vitro studies using BV-2 cells suggest that microglia produce 1.5 fold more PEA than 2-AG [26], we hypothesised that the effects of inhibiting activated microglia on levels of PEA reflected a role of microglia in the hydrolysis of PEA.
To investigate whether differential catabolism of PEA and AEA by microglia may account for our in vivo data, we studied their rates of hydrolysis in intact BV-2 microglia cells. We demonstrate for the first time that the hydrolysis of PEA is sustained, whereas hydrolysis of AEA is transient, in BV-2 cells. These in vitro findings are consistent with our in vivo data, and suggest that the presence of activated microglia in the spinal cord of neuropathic rats contributes to the decreased spinal levels of PEA as a result of the sustained hydrolysis of PEA versus AEA. Conversely, inactivation of spinal microglia by minocycline would result in a decreased microglial component of PEA hydrolysis and the observed increase in levels of PEA in the spinal cord of neuropathic rats treated with minocycline. The differential hydrolysis of PEA and AEA by BV-2 cells suggests that distinct enzymatic activities may be responsible for AEA and PEA catabolism in microglia. Indeed, microglial cells express both fatty acid amide hydrolase and monoacylglycerol lipase, as well as other activities including a PEA hydrolase with a distinct pharmacological profile [22].
When considering the behavioural phenotype of minocycline-treated neuropathic rats versus vehicle-treated rats, the analgesia seen in the minocycline-treated rats may be mediated by the increased levels of AEA observed in neuropathic rats. It is unlikely that 2-AG contributes to the changes in behavioural allodynia, as the marked loss of 2-AG could be expected to exacerbate allodynia. The most prominent effect of minocycline was the significant elevation of ipsilateral PEA in minocycline-treated neuropathic rats, compared to vehicle treated neuropathic rats. PEA has been shown to attenuate neuropathic pain behaviour [35, 36]. Although a role of cannabinoid CB2 receptors in the effects of PEA was initially proposed [37, 38] this was then discounted [39]. Furthermore, the antinociceptive profile of PEA differs from the broad spectrum analgesia produced by systemically administered CB2 receptor agonists [40]. PEA has well described anti-inflammatory effects and has been shown to be neuroprotective [24]. PEA activates the nuclear receptor peroxisome proliferator-activated receptor-α, which mediates the analgesic effects of PEA [41]. Inhibitory effects of PEA in neuropathic rats are also mediated by PPARγ receptors [36]. Our data suggest that the contribution of PEA to the neuroprotective effects of minocycline warrants further investigation.
In conclusion, we have shown that the analgesic effects produced by minocycline treatment are associated with marked changes in levels of 2-AG and PEA and activated microglia in the spinal cord of neuropathic rats. Our data provide evidence for a role of activated microglia in the control of levels of endocannabinoids and related compounds in vivo.
Methods
All experiments were carried out in accordance with the UK Home Office Animals (Scientific Procedures) Act (1986). Experiments were performed on 32 male Sprague Dawley rats (CRUK) in the light period of a 12 hr light/dark cycle. Animals had free access to food and water and were group-housed throughout the experiments. Rats were divided into 4 experimental groups comprising of sham-operated and SNL rats, treated with either vehicle or minocycline.
Model of neuropathic pain: Spinal nerve ligation
The spinal nerve ligation model of neuropathic pain was used in this study. Spinal nerves L5-L6 were ligated according to the procedures described by [42]. Male Sprague Dawley rats (101 – 135 g) were anaesthetised using isoflurane (3% induction, 1–1.5% maintenance in 33% O2/67% N2O) and placed in a prone position. A midline incision was made at the L3-S2 level and the left paraspinal muscles at L4-S2 level were separated from spinal processes. Part of the L6 transverse process was removed with fine rongeurs and the L4-L6 nerves identified. The L5-L6 spinal nerves were isolated and tightly ligated (one suture per nerve) distal to the dorsal root ganglia and proximal to the sciatic nerve formation with 6-0 silk. The wound was closed in two layers using absorbable sutures and wound clips, following complete haemostasis. A similar procedure was performed for the sham surgery, except spinal nerves were not ligated. Post-surgery, the sham-operated and the SNL rats were group-housed (5 rats per cage) and their posture and behaviour were closely monitored for 48 hours.
Drug treatment protocol
Effects of chronic intraperitoneal (ip) injections of minocycline (Sigma, UK, 30 mg/kg, n = 16) or vehicle (2 ml/kg sterile water, n = 16) on mechanical allodynia were measured over 14 days post-surgery in sham-operated and SNL rats. Minocycline (30 mg/kg, ip) or vehicle was injected into rats 1 hour before sham- or SNL surgery and subsequently daily for 14 days, approximately 15 hours prior to behavioural testing.
Behavioural testing
From post-operative day 1 onwards (days 1, 3, 5, 7, 10 and 14), behavioural testing was performed to assess the development of mechanical allodynia, all testing was carried out between 8 and 11 am. Rats were placed in perspex cubicles with wire mesh grid floors and allowed to acclimatise prior to behavioural testing. Mechanical sensitivity of the ipsilateral and contralateral hind paw was assessed by measuring the paw withdrawal threshold (PWT) at which foot withdrawal to normally innocuous mechanical punctate stimuli was observed. Stimuli were delivered, from below, to the plantar surface of the foot using 0.2, 2, 4, 6, 8, 10 and 15 g von Frey hair stimuli for a maximum of 5 seconds per application. Each trial consisted of the application of a single von Frey hair, starting with a 4 g stimulus, and either increasing or decreasing in intensity until the PWT was observed, with the stimulus either side of the withdrawal weight subsequently repeated. The paw withdrawal threshold was recorded as the lowest von Frey stimulus to elicit a response.
Measurement of endocannabinoids and related compounds
On post-operative day 15, approximately 15 hours after the last injection of minocycline (30 mg/kg, ip) or vehicle, rats were decapitated prior to a standardised and rapid dissection of the ipsilateral and contralateral lumbar (L4-L6) spinal cord. The whole process was complete within 3 minutes and samples were placed immediately on dry ice and then stored at -80°C to minimise post-mortem changes in levels of endocannabinoids (ECs) and related compounds. A validated lipid extraction method [43] was employed for measurement of AEA and 2-AG and related compounds, PEA and N-oleoylethanolamine (OEA). In brief, tissue was homogenised in an ethyl acetate/hexane mixture with internal standards (0.42 nmol d8-AEA, 1 nmol d8-2-AG), followed by repeated centrifugation and supernatant layer collection stages. Solid phase extraction was subsequently performed to purify samples. Simultaneous measurement of ECs and related compounds was then performed using liquid chromatography-tandem mass spectrometry (LC-MS/MS). Analysis was carried out on an Agilent 1100 system (Agilent Technologies, Waldbrunn, Germany) coupled to a triple quadrupole Quattro Ultima MS (Waters, Manchester, UK) recording in electrospray positive mode. Analytes were separated chromatographically on a HyPurity Advance C8 column and pre-column (100 × 2.1 mm internal diameter, 3 μm particle size; Thermo Fisher Scientific, Runcorn, UK) with a mobile phase flow rate of 0.3 ml/min. A gradient elution was used, with mobile phases consisting of A (water, 1 g/L ammonium acetate, 0.1% formic acid) and B (acetonitrile, 1 g/L ammonium acetate, 0.1% formic acid). Samples were injected from a cooled auto sampler maintained at 4°C. Multiple reaction monitoring of individual compounds, using specific precursor and product mass to charge (m/z) ratios allowed simultaneous measurement of AEA, 2-AG, PEA and OEA.
Quantification of microglia
In a separate series of experiments, groups of vehicle-treated SNL rats or minocycline-treated SNL rats (n = 4 per group) were prepared for immunohistochemical studies of microglia activation. The development of mechanical allodynia in vehicle-treated SNL rats and the effect of minocycline treatment on mechanical allodynia were consistent with behavioural changes observed in vehicle-treated SNL and minocycline-treated SNL rats employed for measurement of levels of ECs. On post-operative day 15, rats were transcardially perfused with saline (0.9% w/v sodium chloride) followed by paraformaldehyde (PFA, Sigma, UK, 4% w/v) in phosphate-buffered saline (PBS, 3.2 mM Na2HPO4, 0.5 mM KH2PO4, 1.3 mM KCl, 135 mM NaCl, pH 7.4) under terminal anaesthesia induced by pentobarbital sodium (Euthetal®, Merial Animal Health Ltd, UK, 800 mg/kg, ip). The spinal cord was dissected and stored in PFA for approximately 2 hrs at 4°C, and then transferred to a cryoprotectant solution (30% w/v sucrose, 0.1% w/v sodium azide in PBS).
Spinal cords were frozen in O.C.T. embedding medium (VWR, UK) and 14 μm serial sections were cut onto Superfrost Plus slides (VWR) using a cryostat (CM3050S, Leica Microsystems, UK Ltd). Sections were air dried and then treated with 3% hydrogen peroxide in PBS for 30 minutes at room temperature, washed with PBS and then blocked with 5% normal goat serum (Sigma, UK) in PBS/0.2% Triton X-100. They were then incubated overnight with mouse monoclonal anti-OX-42 (AbD Serotec, UK), diluted 1:1000 in PBS/0.2% Triton X-100. This antibody recognizes the complement receptor type 3(CR3) and specifically labels the plasma membrane of the microglial cells. Sections were washed in PBS and incubated with a biotinylated goat anti-mouse IgG secondary (Vector Laboratories, UK) diluted 1:500 in PBS/0.2% Triton X-100 for 2 hours at RT. They were washed again, treated with avidin-biotin complex (Vector Laboratories, UK) according to the manufacturer's instructions, rinsed again and incubated with diaminobenzidine/nickel (Vector Laboratories, UK). The reaction was stopped with PBS and slides were dehydrated and coverslipped. Sections obtained from the eight rats were processed at the same time, and the experiments were repeated three times. Images were acquired using a Leica DMRA2 microscope (Leica, Nussloch, Germany), and digital images were captured using a Retiga 1300 12-bit camera (QImaging, Burnaby, BC, Canada) and QCapture 1.1.6 software (QImaging) running on a Macintosh G4 computer. Images were then assembled into figures using Photoshop 7.0 (Adobe Systems, San Jose, CA). OX-42 optical density was analysed at the level of L4, L5 and L6. To quantify, we applied Image J Software based analysis: nine equivalent boxes (areas of interest, measuring 180 × 180 squared μm) were placed within each dorsal horn covering the medial to lateral extent of Rexed laminae I-V, and the mean grey saturation measured within each area of interest. The mean value of three equivalent boxes outside each section was then subtracted to normalize background. Three sections were analysed at each level. Results (mean ± SEM) are expressed as arbitrary units.
AEA and PEA hydrolysis in microglial cells in vitro
The AEA/PEA hydrolysis assay was based on Paylor et al. [44]. In brief, BV2 mouse microglial cells (a generous gift from N. Stella, University of Washington), passage number <8, grown to confluency in 24 well plates were pre-incubated for 10 min at 37°C in 400 μl HEPES-containing physiological buffer, pH 8.0, containing 0.1% fatty acid-free bovine serum albumin. The reaction was initiated by the addition of 250 nM AEA or PEA containing [3H]-AEA or [3H]-PEA (approx 60,000 dpm/well), respectively. After varying periods of incubation (2 to 30 minutes) at 37°C, the buffer was rapidly removed by aspiration and replaced with 400 μl ice-cold methanol. Wells were scraped and the entire contents of each well collected. Following additions of 400 μl chloroform and 200 μl H2O, the phases were separated by centrifugation at 5,000 g for 2 minutes. Radioactivity in an aliquot of the aqueous, upper phase containing [3H]-ethanolamine was quantified by liquid scintillation spectroscopy.
Data analysis
Behavioural data were statistically analysed using two-way ANOVA with treatment and time as factors, followed by Bonferroni's post-hoc test, and are presented as mean ± SEM of mechanical PWT in grams. Levels of ECs and related compounds were calculated as the ratio of the EC peak area with its internal standard on the chromatogram and are expressed as mol/g wet weight of tissue. Levels of ECs and related compounds were excluded from analysis if the value was greater than 2 standard deviations from the mean, giving n = 6 for all groups except: n = 5 for contralateral vehicle-saline and ipsilateral vehicle-sham. Ipsilateral and contralateral spinal cord levels of AEA, 2-AG, OEA and PEA were statistically compared using a Mann-Whitney test and are presented as mean % of contralateral spinal cord values, or raw mean ± SEM). For comparison of levels of AEA, 2-AG, OEA and PEA between treatment groups, data were statistically compared using Kruskal-Wallis test. Immunohistochemical data were analysed using the Kruskal-Wallis non-parametric test and are presented as mean ± SEM of density of OX-42 staining. Hydrolysis of AEA & PEA was expressed as pmoles/well and was analysed as means ± SEM from six different cell passages investigated in triplicate.
Abbreviations
- AEA:
-
anandamide
- 2-AG:
-
2-arachidonoylglycerol
- CB1 :
-
cannabinoid 1 receptor
- CB2 :
-
cannabinoid 2 receptor
- CNS:
-
central nervous system
- EC:
-
endocannabinoid
- MAPK:
-
mitogen activated protein kinase
- OEA:
-
N-oleoylethanolamine
- PEA:
-
N-palmitoylethanolamine
- PFA:
-
paraformaldehyde
- PWT:
-
paw withdrawal threshold
- SNL:
-
spinal nerve ligation.
References
Chapman V, Finn DP: Analgesic effects of cannabinoids: sites and mechanisms of action. Reviews in Analgesia 2003, 7: 25–41.
Williamson EM, Evans FJ: Cannabinoids in clinical practice. Drugs 2000,60(6):1303–14.
Jhaveri MD, Sagar DR, Elmes SJR, Kendall DA, Chapman V: Cannabinoid CB2 receptor-mediated anti-nociception in models of acute and chronic pain. Mol Neurobiol 2007,36(1):26–35.
Yamamoto W, Mikami T, Iwamura H: Involvement of central cannabinoid CB2 receptor in reducing mechanical allodynia in a mouse model of neuropathic pain. Eur J Pharmacol 2008,583(1):56–61.
Yao BB, Hsieh GC, Frost JM, Fan Y, Garrison TR, Daza AV, Grayson GK, Zhu CZ, Pai M, Chandran P, Salyers AK, Wensink EJ, Honore P, Sullivan JP, Dart MJ, Meyer MD: In vitro and in vivo characterization of A-796260: a selective cannabinoid CB2 receptor agonist exhibiting analgesic activity in rodent pain models. Br J Pharmacol 2008,153(2):390–401.
Sagar DR, Kelly S, Millns PJ, O'Shaughnessey CT, Kendall DA, Chapman V: Inhibitory effects of CB1 and CB2 receptor agonists on responses of DRG neurons and dorsal horn neurons in neuropathic rats. Eur J Neurosci 2005,22(2):371–9.
Zhang J, Hoffert C, Vu HK, Groblewski T, Ahmad S, O'Donnell D: Induction of CB2 receptor expression in the rat spinal cord of neuropathic but not inflammatory chronic pain models. Eur J Neurosci 2003,17(12):2750–4.
Beltramo M, Bernardini N, Bertorelli R, Campanella M, Nicolussi E, Fredduzzi S, Reggiani A: CB2 receptor mediated anti-hyperalgesia: possible direct involvement of neural mechanisms. Eur J Neurosci 2006, 23: 1530–1538.
Romero-Sandoval A, Nutile-McMenemy N, DeLeo JA: Spinal microglial and perivascular cell cannabinoid receptor type 2 activation reduces behavioral hypersensitivity without tolerance after peripheral nerve injury. Anesthesiology 2008,108(4):722–34.
Wotherspoon G, Fox A, McIntyre P, Colley S, Bevan S, Winter J: Peripheral nerve injury induces cannabinoid receptor 2 protein expression in rat sensory neurons. Neuroscience 2005,135(1):235–45.
Cabral GA, Harmon KN, Carlisle SJ: Cannabinoid-mediated inhibition of inducible nitric oxide production by rat microglial cells: evidence for CB1 receptor participation. Adv Exp Med Biol 2001, 493: 207–14.
Cabral GA, Raborn ES, Griffin L, Dennis J, Marciano-Cabral F: CB2 receptors in the brain: role in central immune function. Br J Pharmacol 2008, 153: 240–51.
Inoue K, Tsuda M, Tozaki-Saitoh H: Modification of neuropathic pain sensation through microglial ATP receptors. Purinergic Signal 2007,3(4):311–316.
Narita M, Yoshida T, Nakajima M, Narita M, Miyatake M, Takagi T, Yajima Y, Suzuki Tyr2006: Direct evidence for spinal cord microglia in the development of a neuropathic pain-like state in mice. J Neurochem 2006, 97: 1337–1348.
Bye N, Habgood MD, Callaway JK, Malakooti N, Potter A, Kossmann T, Morganti-Kossmann MC: Transient neuroprotection by minocycline following traumatic brain injury is associated with attenuated microglial activation but no changes in cell apoptosis or neutrophil infiltration. Exp Neurol 2007, 204: 220–233.
Choi Y, Kim H-S, Shin KY, Kim EM, Kim M, Kim HS, Park CH, Jeong YH, Yoo J, Lee JP, Chang KA, Kim S, Suh YH: Minocycline attenuates neuronal cell death and improves cognitive impairment in Alzheimer's disease models. Neuropsychopharmacology. 2007,32(11):2393–2404.
Raghavendra V, Tanga F, DeLeo JA: Inhibition of microglial activation attenuates the development but not existing hypersensitivity in a rat model of neuropathy. J Pharmacol Exp Ther 2003,306(2):624–30.
Ledeboer A, Sloane EM, Milligan ED, Frank MG, Mahony JH, Maier SF, Watkins LR: Minocycline attenuates mechanical allodynia and proinflammatory cytokine expression in rat models of pain facilitation. Pain 2005,115(1–2):71–83.
Piao ZG, Cho IH, Park CK, Hong JP, Choi SY, Lee SJ, Lee S, Park K, Kim JS, Oh SB: Activation of glia and microglial p38 MAPK in medullary dorsal horn contributes to tactile hypersensitivity following trigeminal sensory nerve injury. Pain 2006,121(3):219–31.
Wallace VC, Blackbeard J, Segerdahl AR, Hasnie F, Pheby T, McMahon SB, Rice AS: Characterization of rodent models of HIV-gp120 and anti-retroviral-associated neuropathic pain. Brain 2007,130(Pt 10):2688–702.
Carrier EJ, Kearn CS, Barkmeier AJ, Breese NM, Yang W, Nithipatikom K, Pfister SL, Campbell WB, Hillard CJ: Cultured rat microglial cells synthesize the endocannabinoid 2-arachidonylglycerol, which increases proliferation via a CB2 receptor-dependent mechanism. Mol Pharmacol 2004,65(4):999–1007.
Muccioli GG, Xu C, Odah E, Cudaback E, Cisneros JA, Lambert DM, López Rodríguez ML, Bajjalieh S, Stella N: Identification of a novel endocannabinoid-hydrolyzing enzyme expressed by microglial cells. J Neurosci 2007,27(11):2883–9.
Petrosino S, Palazzo E, de Novellis V, Bisogno T, Rossi F, Maione S, Di Marzo V: Changes in spinal and supraspinal endocananbinoid levels in neuropathic rats. Neuropharmacology. 2007,52(2):415–422.
Schomacher M, Muller HD, Sommer C, Schwab S, Schabitz WR: Endocannabinoids mediate neuroprotection after transient focal cerebral ischemia. Brain Res 2008, 1240: 213–220.
Leung D, Saghatelian A, Simon GM, Cravatt BF: Inactivation of N-acyl phosphatidylethanolamine phospholipase D reveals multiple mechanisms for the biosynthesis of endocannabinoids. Biochemistry 2006,45(15):4720–4726.
Muccioli GG, Stella N: Microglia produce and hydrolyze palmitoylethanolamide. Neuropharmacology 2008,54(1):16–22.
Walter L, Franklin A, Witting A, Wade C, Xie Y, Kunos G, Mackie K, Stella N: Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J Neurosci 2003,23(4):1398–405.
Blackbeard J, O'Dea KP, Wallace VC, Segerdahl A, Pheby T, Takata M, Field MJ, Rice AS: Quantification of the rat spinal microglial response to peripheral nerve injury as revealed by immunohistochemical image analysis and flow cytometry. J Neurosci Methods 2007,164(2):207–17.
Clark AK, Gentry C, Bradbury EJ, McMahon SB, Malcangio M: Role of spinal microglia in rat models of peripheral nerve injury and inflammation. Eur J Pain 2007, 11: 223–230.
Tsuda M, Inoue K, Salter MW: Neuropathic pain and spinal microglia: a big problem from molecules in small glia. Trends Neurosci. 2005,28(2):101–107.
Gonzalez JC, Egea J, Del Carmen Godino M, Fernandez-Gomez FJ, Sanchez-Prieto J, Gandia L, Garcia AG, Jordan J, Hernandez-Guijo JM: Neuroprotectant minocycline depresses gluatamatergic neurotransmission and Ca2+ signalling in hippocampal neurones. Eur J Neurosci 2007, 26: 2481–2495.
Hailer NP: Immunosuppression after traumatic or ischemic CNS damage: it is neuroprotective and illuminates the role of microglial cells. Progress in Neurobiology 2008, 84: 211–233.
Walter L, Franklin A, Witting A, Möller T, Stella N: Astrocytes in culture produce anandamide and other acylethanolamides. J Biol Chem 2002, 277: 20869–20876.
Witting A, Walter L, Wacker J, Möller T, Stella N: P2X7 receptors control 2-arachidonoylglycerol production by microglial cells. Proc Natl Acad Sci USA 2004, 101: 3214–9.
Lo Verme J, Russo R, La Rana G, Fu J, Farthing J, Mattace-Raso G, Meli R, Hohmann A, Calignano A, Piomelli D: Rapid broad-spectrum analgesia through activation of peroxisome proliferator-activated receptor-α. J Pharmacol Exp Ther. 2006,319(3):1051–1061.
Costa B, Comelli F, Bettoni I, Colleoni M, Giagnoni G: The endogenous fatty acid amide, palmitoylethanolamide, has anti-allodynic and anti-hyperalgesic effects in a murine model of neuropathic pain: involvement of CB(1), TRPV1 and PPARgamma receptors and neurotrophic factors. Pain 2008, 139: 541–550.
Calignano A, La Rana G, Giuffrida A, Piomelli D: Control of pain initiation by endogenous cannabinoids. Nature 1998,394(6690):277–81.
Facci L, Dal Toso R, Romanello S, Buriani A, Skaper SD, Leon A: Mast cells express a peripheral cannabinoid receptor with differential sensitivity to anandamide and palmitoylethanolamide. Proc Natl Acad Sci 1995,92(8):3376–80.
Griffin G, Tao Q, Abood ME: Cloning and pharmacological characterization of the rat CB(2) cannabinoid receptor. J Pharmacol Exp Ther 2000,292(3):886–94.
Walker JM, Hohmann AG: Cannabinoid mechanisms of pain suppression. Handb Exp Pharmacol 2005, 168: 509–54.
Lo Verme J, Fu J, Astarita G, La Rana G, Russo R, Calignano A, Piomelli D: The nuclear receptor peroxisome proliferator-activated receptor-alpha mediates the anti-inflammatory actions of palmitoylethanolamide. Mol Pharmacol 2005,67(1):15–9.
Kim SH, Chung JM: An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992,50(3):355–63.
Richardson D, Ortori CA, Chapman V, Kendall DA, Barrett DA: Quantitative profiling of endocannabinoids and related compounds in rat brain using liquid chromatography-tandem electrospray ionization mass spectrometry. Anal Biochem 2007,360(2):216–26.
Paylor B, Holt S, Fowler CJ: The potency of the fatty acid amide hydrolase inhibitor URB597 is dependent upon the assay pH. Pharmacological Research 2006, 54: 481–485.
Acknowledgements
This research was supported by the Wellcome Trust (grant: 066203/Z/01/Z) and the Medical Research Council (grant: G0401647). Khalil Eldeeb's studentship was supported by the Faculty of Medicine, Al-Azhar University, Dammietta, Egypt.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
LG carried out the quantification of microglia and statistical analysis. DR carried out the drug dosing, behavioural testing, quantification of endocannabinoids and related compounds and statistical analysis. MJ generated neuropathic rats and carried out behavioural testing. KE carried out hydrolysis assays in BV2 cells and statistical analysis. DAB participated in study design. MRE participated in study design. SPHA participated in study design. DAK participated in study design. GJM participated in study design. VC generated neuropathic rats, conceived the study, and participated in its design and coordination. All authors read and approved the final manuscript.
Leonardo Guasti, Denise Richardson contributed equally to this work.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Guasti, L., Richardson, D., Jhaveri, M. et al. Minocycline treatment inhibits microglial activation and alters spinal levels of endocannabinoids in a rat model of neuropathic pain. Mol Pain 5, 35 (2009). https://doi.org/10.1186/1744-8069-5-35
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1744-8069-5-35