
Abstract
Inflammatory mediators can directly sensitize primary afferent nociceptors to mechanical and osmotic stimuli. Sensitized nociceptors have a lowered threshold of activation and increased spontaneous activity, which result in symptoms of hyperalgesia and pain, respectively. The transient receptor potential vanilloid 4 (TRPV4) ligand-gated ion channel has been implicated in the hyperalgesia for mechanical and osmotic stimuli associated with inflammatory states. To investigate whether TRPV4 directly contributes to the mechanisms of inflammatory mediator sensitization of C-fiber nociceptors, we compared the effect of the injection of simplified inflammatory soup (prostaglandin E2 and serotonin) into the mechanical receptive fields of C-fibers in TRPV4+/+ and TRPV4-/- mice in vivo. Following the injection of the soup, the percentage of C-fibers responding to a hypotonic stimulus and the magnitude of the response was significantly greater in TRPV4+/+ mice compared to TRPV4-/- mice. Moreover, in response to simplified inflammatory soup only C-fibers from TRPV4+/+ mice exhibited increased spontaneous activity and decreased mechanical threshold. These marked impairments in the response of C-fibers in TRPV4-/- mice demonstrate the importance of TRPV4 in nociceptor sensitization; we suggest that TRPV4, as TRPV1, underlies the nociceptive effects of multiple inflammatory mediators on primary afferent.
Similar content being viewed by others


Background
Transient receptor potential vanilloid 4 (TRPV4), a member of the vanilloid subfamily of transient receptor potential ligand-gated ion channels, cloned from hypothalamus using a functional assay screening for osmo-sensitivity [1] or kidney [2], is also present in sensory neurons that express properties of nociceptors [3, 4]. Accumulating data support a role of TRPV4 in nociception: 1) mice lacking a functional TRPV4 gene show impaired responses to intense noxious mechanical stimuli but normal responses to low-threshold mechanical stimuli [5, 6], 2) TRPV4 plays an important role in hyperalgesia to osmotic and mechanical stimuli generated by inflammatory mediators [7, 8], and 3) inflammatory mediators can engage TRPV4 in hyperalgesia to mechanical and osmotic stimuli [9]. While primary afferent nociceptors in the rat respond to hypotonic stimuli, an effect that is enhanced by prostaglandin E2 [7] on the role of TRPV4 is unknown. To establish the role of TRPV4, in vivo, in peripheral nociceptor sensitization, we performed a single fiber electrophysiology study of primary afferent nociceptors in TRPV4+/+ and TRPV4-/- mice.
Results
There were no significant differences in the average conduction velocity and baseline mechanical threshold for C-fibers from TRPV4+/+ and TRPV4-/- mice (unpaired t- and Mann Whitney test, respectively, both p > 0.05). The average conduction velocity of C-fibers from TRPV4+/+ and TRPV4-/- mice were 1.1 ± 0.1 and 1.0 ± 0.1 m/sec, respectively. And the average baseline mechanical threshold of C-fibers from TRPV4+/+ and TRPV4-/- mice were 23.7 ± 7.86 and 16.2 ± 5.73 mN, respectively. Their receptive fields were both approximately 2 mm across. However, in TRPV4-/- mice C-fiber spontaneous activity was 4.15 ± 1.61 spikes/min, which was significantly higher than in TRPV4+/+ controls (0.18 ± 0.18 spikes/min, unpaired t-test, p < 0.05). Of note, only one C-fiber from a TRPV4+/+ mouse had spontaneous activity, at a very low frequency (2 spikes/min), while 38.5% (5/13) of C-fibers from TRPV4-/- mice had low frequency spontaneous activity (average, 11 spikes/min, n = 5, p < 0.05).
Approximately half of C-fibers in both TRPV4+/+ and TRPV4-/- mice were excited by intradermal injection of simplified inflammatory soup, adjacent to their mechanical receptive fields. The magnitude of activity evoked by injection of simplified inflammatory soup was similar between TRPV4+/+ and TRPV4-/- mice (p = ns, Fig. 1).

The average time course of the response of C-fibers during the 60 sec period after injection of simplified inflammatory soup was not significantly different in TRPV4+/+ and TRPV4-/- mice (unpaired t-test, p > 0.05). The bin width is 1 sec. A. The open bars represent activity evoked by simplified inflammatory soup in TRPV4+/+ C-fibers (n = 11), and B. The filled bars represent action potentials evoked in TRPV4-/- C-fibers (n = 13).
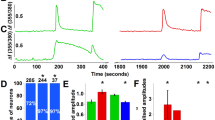
Almost all C-fibers from TRPV4+/+ mice had no spontaneous activity before applying stimuli on the receptive fields, except one fiber with very low frequency of on-going activity (2 spikes/min). While baseline spontaneous activity was greater in TRPV4-/- mice, on-going activity after injection of simplified inflammatory soup was significantly increased in TRPV4+/+ C-fibers (p < 0.05), while it was unchanged in the TRPV4-/- C-fibers (p = ns, Fig. 2A). Also, while there was no difference in baseline threshold between C-fibers in TRPV4+/+ and TRPV4-/- mice, after injection of simplified inflammatory soup, the mechanical threshold of C-fibers in TRPV4+/+ but not TRPV4-/- mice significantly decreased (p < 0.05, Fig. 2B).

A. On-going activity in C-fibers before (open bar) and after (filled bar) injection of simplified inflammatory soup into each fiber's mechanical receptive field was significantly increased in TRPV4+/+ (n = 11, paired t-test, * p < 0.05) but not TRPV4-/- mice (n = 13, paired t-test, p > 0.05). B. The mechanical threshold of C-fibers in TRPV4+/+ (open bar, n = 11) produced by intradermal injection of simplified inflammatory soup was statistically significant (Wilcoxon matched test, * p < 0.05). However, simplified inflammatory soup did not significantly change mechanical threshold of C-fibers in TRPV4-/- mice (filled bars, n = 13, Wilcoxon matched test, p > 0.05). The change in mechanical threshold of C-fibers after simplified inflammatory soup was significantly greater in TRPV4+/+ than TRPV4-/- C-fibers (χ2 test, * p < 0.05).
Finally, in TRPV4+/+ mice, 81.2% (9/11) of C-fibers responded to intradermal hypotonic solution injected 15 minutes after intradermal injection of simplified inflammatory soup; only 33.3% (3/9) of C-fibers responded in TRPV4-/- mice (Fig. 3, p < 0.05).

Upper panel, response of a TRPV4+/+ C-fiber to hypotonic solution 15 min after injection of simplified inflammatory soup. Arrow indicates the time of injection of hypotonic solution. Lower panel, the average time course of the response of C-fibers during the first 60 sec after injection of hypotonic solution in TRPV4+/+ mice (open bar, n = 11). The bin width is 1 sec. B. Upper panel, response of a TRPV4-/- C-fiber to hypotonic solution after injection of simplified inflammatory soup. Lower panel, the average time course of the response of C-fibers during the first 60 sec after injection of hypotonic solution in TRPV4-/- mice (filled bars, n = 9).
Discussion
Recent studies support a role of TRPV4 in inflammatory mediator-induced sensitization to osmotic and mechanical stimuli: 1) TRPV4 contributes to the enhanced nociceptive behaviors in response to mechanical and osmotic stimuli, produced by inflammation and peripheral neuropathy [4, 8], 2) inflammatory mediators can engage TRPV4 in DRG neurons in vitro [9] and 3) hypotonic stimuli elicit action potentials in C-fibers, and in presence of inflammatory mediators, the number of action potentials significantly increases, as does the percentage of responsive fibers [7]. To establish the contribution of TRPV4 in inflammatory mediator-induced sensitization of C-fiber nociceptors, in vivo, we compared the effect of inflammatory mediators that engage TRPV4 function in behavioral studies, on the response of C-fibers to osmotic and mechanical stimulation in TRPV4+/+ and TRPV4-/- mice. The percentage of C-fibers responding to hypotonicity as well as the magnitude of the response, is significantly greater in TRPV4+/+ but not TRPV4-/- mice (Fig. 3). This finding demonstrates a contribution of TRPV4 to inflammatory mediator-induced sensitization of C-fibers to hypotonic stimuli. This finding also has clinical relevance since water is an irritant, exacerbating painful conditions [10–12], and changes in tissue osmolarity have been reported in diseases that are associated with painful peripheral neuropathy such as diabetes [13] and alcoholism [14].
However, recent reports suggest that TRPV4 may also act as a mechano-transducer in primary afferent nociceptors; mice lacking a functional TRPV4 gene have impaired behavioral responses to intense noxious mechanical stimuli but normal response to low-threshold mechanical stimuli [5, 6], and spinal intrathecal administration of oligodeoxynucleotides antisense to TRPV4 reverses mechanical hyperalgesia in a rat model of small-fiber painful peripheral neuropathy induced by the cancer chemotherapy agent Taxol® [4]. In addition, while the baseline mechanical paw-withdrawal threshold is not significantly different between TRPV4+/+ and TRPV4-/- mice, after intraplantar injection of simplified inflammatory soup, mechanical hyperalgesia only occurred in TRPV4+/+ mice [9]. Similarly, mechanical hyperalgesia induced by simplified inflammatory soup, in the rat, is prevented by spinal intrathecal treatment with TRPV4 antisense [9]. These findings suggested a role for TRPV4 in inflammatory mediator-induced sensitization of nociceptors to mechanical stimuli. Our present study actually demonstrated, in vivo, the role of TRPV4 in nociceptor sensitization, the mechanism underlying primary mechanical hyperalgesia. In agreement with behavioral studies demonstrating similar mechanical nociceptive thresholds in TRPV4+/+ and TRPV4-/- mice [5, 6, 9], the mechanical thresholds of C-fibers from TRPV4+/+ and TRPV4-/- mice were not significantly different. However, intradermal injection of simplified inflammatory soup lowered mechanical threshold in TRPV4+/+ but not TRPV4-/- C-fibers, supporting the idea of an in vivo contribution of TRPV4 to inflammatory mediator-induced sensitization of primary afferent nociceptors to mechanical stimuli.
Inflammatory mediators, such as PGE2 and 5-HT act directly on primary afferent nociceptors, to decrease their threshold of activation, increase spontaneous activity and increase response to supra-threshold stimuli (for review) [15]; whereas spontaneous activity of peripheral nociceptors may contribute to ongoing pain [16]. In this regard, it is noteworthy that simplified inflammatory soup enhanced spontaneous activity in C-fibers from TRPV4+/+ but not TRPV4-/- mice. This result is also compatible with the recent suggestion that depending on the cellular context some TRP channels may mediate increases in neuronal excitability through activation of intracellular signaling pathways rather than through the binding of a specific ligand [17]. We recently demonstrated that the simplified inflammatory soup constituting two inflammatory mediators (PGE2 and 5-HT) can act synergistically through cAMP to engage TRPV4 in mechanical hyperalgesia [9]. The role of TRPV4 not only in mechanical and osmotic hyperalgesia but also in the inflammatory mediator-induced increase in spontaneous activity in primary afferent nociceptors suggests that TRPV4 contributes to peripheral sensitization via the nociceptive effects of multiple inflammatory mediators. Moreover, to our knowledge this is the first report suggesting a role for TRPV4 in any form of spontaneous pain.
Not previously reported, TRPV4 may also play a role in the regulation of spontaneous activity of nociceptors, even in the absence of inflammation; spontaneous activity of C-fibers is about 20-fold higher in TRPV4-/- mice than in TRPV4+/+ mice. Thus, low level of on-going activity in primary afferents may not always result in spontaneous pain. The significant change of primary afferent activities can be more important as we observed in TRPV4+/+ mice after simplified inflammatory soup. Of note, C-fiber conduction velocity is similar in TRPV4+/+ and TRPV4-/- mice. Since nerve conduction velocity is a measure of the excitability of voltage-sensitive channels in neurons, a variation in its value would reflect compensatory changes in voltage-gated ion channels in the axon [18–20], the difference in spontaneous activity is less likely to result from compensatory changes in most of the voltage-gated channels found in axons, but may reflect a contribution of TRPV4 in the modulation of membrane potential and the generation of action potentials in primary afferent nociceptors. The contribution of TRPV4 to spontaneous activity in primary afferent nociceptors, both in physiological and inflammatory states, support findings in other cell types where TRPV4 alters cell function by modulating complex calcium events [21–23]. Of course, we cannot rule out the possibility that subtle compensatory abnormalities in ion channel function in the TRPV4-/- mouse are responsible for the change of the spontaneous activity. Anyway, the actual mechanism of spontaneous activity of primary afferents in TRPV4-/- mice remains to be further elucidated.
Conclusion
Using in vivo single fiber electrophysiology, we demonstrate a role of TRPV4 in inflammatory mediator-induced sensitization of C-fibers to hypotonic and mechanical stimuli. Moreover, TRPV4 may play a broader role in inflammatory pain, also contributing to the spontaneous pain induced by the action of inflammatory mediators. Therefore, we suggest that TRPV4 may play a similar role as TRPV1 which underlines mechanical and thermal sensitivities in the sensitization of primary nociceptive afferent, acting as a final pathway for the nociceptive effects of multiple inflammatory mediators and second messenger signaling to mediate primary afferent nociceptor sensitization to mechanical stimuli and, therefore, mechanical hyperalgesia.
Methods
Animal model
Experiments were performed on 24–34 g adult male C57BL6 mice lacking a functional TRPV4 gene (TRPV4-/-) and TRPV4 wild-type (TRPV4+/+) littermates [5]. A controlled heater (TR-200, FST, Foster City, CA) was used to maintain body temperature (≥ 36.5°C). Animal care and use conformed to National Institutes of Health (NIH) guidelines and was approved by the UCSF Committee on Animal Research.
Electrophysiology
In vivo single fiber electrophysiology experiments were performed as described previously in rats [24, 25] and mice [26]. Briefly, mice were anesthetized with sodium pentobarbital (initially 50 mg/kg, i.p., with additional doses given throughout the experiment to maintain areflexia). Recordings were made from C-fibers in the saphenous nerve, which innervates part of the dorsal surface of the hind paw. Bipolar stimulating electrodes were placed under the nerve, distal to the recording site. The nerve was cut proximal to the recording site and put on a platform for further dissection. Fine fascicles of axons were teased from the nerve and placed on a recording electrode. There usually are more than one fiber in the fasicle from which our recordings are made (often two or three). Single units were first detected by electrical stimulation of the nerve. Each fiber's conduction velocity was calculated by dividing the distance between the stimulating and recording electrodes by the latency of the electrically-evoked action potential. Fibers that conducted slower than 2 m/s were classified as C-fibers [27, 28]. C-fiber receptive fields were located using a mechanical search stimulus, either a blunt probe with smooth tip or a 60 g von Frey hair (VFH). The electrically evoked action potential corresponding to the C-fiber whose mechanical receptive field had been identified was verified by the latency delay technique, in which electrically evoked spikes resulted in longer latency when the receptive field of the same fiber was stimulated mechanically [29]. Mechanical threshold was determined with calibrated VFH and defined as the lowest force that elicited ≥ 2 spikes within 1 s, in at least 50% of trials.
The spontaneous activity was expressed in spikes/min and measured one minute before and after injection of soup, respectively. The neural activity of C-fibers was captured and stored by using an IBM-compatible computer with MICRO 1401 interface (CED, Cambridge, UK) and further analyzed with Spike2 software (CED). Generally, the studied unit is the largest unit which can be easily captured by a window discriminator (Winston Electronics, San Francisco, CA) or by Spike2 software according to the amplitude and duration of action potentials. At the end of the experiment the mouse was euthanized by sodium pentobarbital followed by bilateral thoracotomy.
Injections of simplified inflammatory soup and hypotonic solution
The simplified inflammatory soup used in these experiments was composed of the combination of prostaglandin E2 (PGE2) and serotonin (5-HT), both purchased from Sigma (St. Louis, MO), dissolved in 0.9% saline. A 2.5 μl volume of simplified inflammatory soup containing 100 ng each of PGE2 and 5-HT, was injected adjacent to a C-fiber's mechanical receptive field, which produced mechanical hyperalgesia in behavioral studies [9]. Hypotonic solution (deionized distilled water, 2.5 μl) was injected into the receptive field of an identified C-fiber, 15 min after the simplified inflammatory soup; after the simplified inflammatory soup was injected, mechanical threshold was re-measured, before recording the response to re-injection of hypotonic solution.
Statistics
Group data are expressed as mean ± SEM. Statistical analyses were done using paired or unpaired t-test, Wilcoxon matched or Mann Whitney and χ2 test, as appropriate. Differences were considered significant at a p-value of <0.05.
References
Liedtke W, Choe Y, Marti-Renom MA, Bell AM, Denis CS, Sali A, Hudspeth AJ, Friedman JM, Heller S: Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell 2000, 103: 525–535. 10.1016/S0092-8674(00)00143-4
Strotmann R, Harteneck C, Nunnenmacher K, Schultz G, Plant TD: OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat Cell Biol 2000, 2: 695–702. 10.1038/35036318
Suzuki M, Watanabe Y, Oyama Y, Mizuno A, Kusano E, Hirao A, Ookawara S: Localization of mechanosensitive channel TRPV4 in mouse skin. Neurosci Lett 2003, 353: 189–192. 10.1016/j.neulet.2003.09.041
Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD: Transient receptor potential vanilloid 4 is essential in chemotherapy-induced neuropathic pain in the rat. J Neurosci 2004, 24: 4444–4452. 10.1523/JNEUROSCI.0242-04.2004
Liedtke W, Friedman JM: Abnormal osmotic regulation in trpv4-/- mice. Proc Natl Acad Sci U S A 2003, 100: 13698–13703. 10.1073/pnas.1735416100
Suzuki M, Mizuno A, Kodaira K, Imai M: Impaired pressure sensation in mice lacking TRPV4. J Biol Chem 2003, 278: 22664–22668. 10.1074/jbc.M302561200
Alessandri-Haber N, Yeh JJ, Boyd AE, Parada CA, Chen X, Reichling DB, Levine JD: Hypotonicity induces TRPV4-mediated nociception in rat. Neuron 2003, 39: 497–511. 10.1016/S0896-6273(03)00462-8
Alessandri-Haber N, Joseph E, Dina OA, Liedtke W, Levine JD: TRPV4 mediates pain-related behavior induced by mild hypertonic stimuli in the presence of inflammatory mediator. Pain 2005, 118: 70–79. 10.1016/j.pain.2005.07.016
Alessandri-Haber N, Dina OA, Joseph EK, Reichling D, Levine JD: A transient receptor potential vanilloid 4-dependent mechanism of hyperalgesia is engaged by concerted action of inflammatory mediators. J Neurosci 2006, 26: 3864–3874. 10.1523/JNEUROSCI.5385-05.2006
Tsai TF, Maibach HI: How irritant is water? An overview. Contact Dermatitis 1999, 41: 311–314.
Shelley WB, Shelley ED: Aquadynia: noradrenergic pain induced by bathing and responsive to clonidine. J Am Acad Dermatol 1998, 38: 357–358. 10.1016/S0190-9622(98)70583-3
Fujimura M, Amemiya T, Myou S, Ishiura Y, Sasaki S, Matsuda T: A partial involvement of histamine in ultrasonically nebulized distilled water-induced bronchoconstriction in guinea-pigs. Br J Pharmacol 1998, 125: 902–908. 10.1038/sj.bjp.0702135
Puliyel JM, Bhambhani V: Ketoacid levels may alter osmotonicity in diabetic ketoacidosis and precipitate cerebral edema. Arch Dis Child 2003, 88: 366. 10.1136/adc.88.4.366-a
Vamvakas S, Teschner M, Bahner U, Heidland A: Alcohol abuse: potential role in electrolyte disturbances and kidney diseases. Clin Nephrol 1998, 49: 205–213.
Millan MJ: The induction of pain: an integrative review. Prog Neurobiol 1999, 57: 1–164. 10.1016/S0301-0082(98)00048-3
Djouhri L, Koutsikou S, Fang X, McMullan S, Lawson SN: Spontaneous pain, both neuropathic and inflammatory, is related to frequency of spontaneous firing in intact C-fiber nociceptors. J Neurosci 2006, 26: 1281–1292. 10.1523/JNEUROSCI.3388-05.2006
Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J, Yamoah EN, Basbaum AI, Julius D: TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124: 1269–1282. 10.1016/j.cell.2006.02.023
Brown AM: Simulation of axonal excitability using a Spreadsheet template created in Microsoft Excel. Comput Methods Programs Biomed 2000, 63: 47–54. 10.1016/S0169-2607(00)00076-6
Flake NM, Lancaster E, Weinreich D, Gold MS: Absence of an association between axotomy-induced changes in sodium currents and excitability in DRG neurons from the adult rat. Pain 2004, 109: 471–480. 10.1016/j.pain.2004.02.024
Dawson GD: The relative excitability and conduction velocity of sensory and motor nerve fibres in man. J Physiol 1956, 131: 436–451.
Andrade YN, Fernandes J, Vazquez E, Fernandez-Fernandez JM, Arniges M, Sanchez TM, Villalon M, Valverde MA: TRPV4 channel is involved in the coupling of fluid viscosity changes to epithelial ciliary activity. J Cell Biol 2005, 168: 869–874. 10.1083/jcb.200409070
Earley S, Heppner TJ, Nelson MT, Brayden JE: TRPV4 forms a novel Ca2+ signaling complex with ryanodine receptors and BKCa channels. Circ Res 2005, 97: 1270–1279. 10.1161/01.RES.0000194321.60300.d6
Reiter B, Kraft R, Gunzel D, Zeissig S, Schulzke JD, Fromm M, Harteneck C: TRPV4-mediated regulation of epithelial permeability. Faseb J 2006, 20: 1802–1812. 10.1096/fj.06-5772com
Ahlgren SC, Wang JF, Levine JD: C-fiber mechanical stimulus-response functions are different in inflammatory versus neuropathic hyperalgesia in the rat. Neuroscience 1997, 76: 285–290. 10.1016/S0306-4522(96)00290-4
Chen X, Levine JD: Hyper-responsivity in a subset of C-fiber nociceptors in a model of painful diabetic neuropathy in the rat. Neuroscience 2001, 102: 185–192. 10.1016/S0306-4522(00)00454-1
Cain DM, Khasabov SG, Simone DA: Response properties of mechanoreceptors and nociceptors in mouse glabrous skin: an in vivo study. J Neurophysiol 2001, 85: 1561–1574.
Lynn B, Carpenter SE: Primary afferent units from the hairy skin of the rat hind limb. Brain Res 1982, 238: 29–43. 10.1016/0006-8993(82)90768-5
Willis WD: The pain system: the neural basis of nociceptive transmission in the mammalian nervous system. In Pain and headache. Volume 8. Basel; New York, Karger; 1985:346.
Iggo A: The electrophysiological identification of single nerve fibers, with particular reference to the slowest-conduction vagal afferent fibers in the cat. J Physiol (Lond) 1958, 142: 110–126.
Acknowledgements
We thank Drs. Wolfgang Liedtke (Duke University) and Jeffery Friedman (Rockefeller University) for kindly providing the TRPV4 knockout mice. This research was supported by NIH grant NS21647.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
XC participated in the design of the study, carried out all the experiment, performed the statistical analysis and drafted the manuscript. NA bred the mice and joined drafting the manuscript. JDL participated in the design of the study and drafted the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Chen, X., Alessandri-Haber, N. & Levine, J.D. Marked attenuation of inflammatory mediator-induced C-fiber sensitization for mechanical and hypotonic stimuli in TRPV4-/- mice. Mol Pain 3, 31 (2007). https://doi.org/10.1186/1744-8069-3-31
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1744-8069-3-31