
Abstract
The cerebrospinal fluid (CSF) system provides nutrients to and removes waste products from the brain. Recent findings suggest, however, that in addition, the CSF contains message molecules in the form of actively released neuroactive substances. The concentrations of these vary between locations, suggesting they are important for the changes in brain activity that underlie different brain states, and induce different sensory input and behavioral output relationships.
The cranial CSF displays a rapid caudally-directed ventricular flow followed by a slower rostrally-directed subarachnoid flow (mainly towards the cribriform plate and from there into the nasal lymphatics). Thus, many brain areas are exposed to and can be influenced by substances contained in the CSF. In this review we discuss the production and flow of the CSF, including the mechanisms involved in the regulation of its composition. In addition, the available evidence for the release of neuropeptides and other neuroactive substances into the CSF is reviewed, with particular attention to the selective effects of these on distant downstream receptive brain areas. As a conclusion we suggest that (1) the flowing CSF is involved in more than just nutrient and waste control, but is also used as a broadcasting system consisting of coordinated messages to a variety of nearby and distant brain areas; (2) this special form of volume transmission underlies changes in behavioral states.
Similar content being viewed by others
Review outline
Introduction
Production and circulation of CSF
Production and sources of CSF
Flow and destiny of the CSF
The CSF circulation during development and aging of the CNS
CSF composition: sources, targets and exchange with ECF
Sources of CSF contents
Flow-transport: speed and mechanisms
Exchange of substances between CSF and ECF; mechanisms and targets:
Neuropeptides form an integral part of the CSF contents
Behavioral effects of CSF contents
Conclusions
Introduction
Behavioral observations as well as modern imaging techniques show that brain activity is an input-output process that depends on the physiological or behavioral state of the animal or subject. A state is a well-known concept in mathematical system theory and in computer science. A machine, but for that matter also an organism or even a human, is at moment t1 in the same state as at moment t2, if for all possible input, the resulting internal and external reactions are the same. The states will be different if there is a particular input resulting in different reactions. Of course, the collection of all possible input (and also reactions) is huge and in a particular scientific discipline one restricts oneself to a set of relevant inputs and reactions. Many well-known examples of states come from animal observations, and have been described as motivational, emotional and mental states, or simply brain states in animal and human behavioral studies [1–5] and in a phenomenological study [6].
Changes in behavioral states, which induce adaptive physiological mechanisms, require a complex rearrangement of neural activity in a wide variety of brain areas. The hypothalamus, embedded in the limbic system is strongly involved when a motivational state changes [3–5, 7, 8]. Other brain areas, such as cortical regions or spinal cord, must be involved as well, in order to accomplish coordinated adaptations in the neural (sensory, motor and autonomic) as well as hormonal systems, that underlie goal-directed behavior.
Neural networks involved in the coordination of these changes are complex. Synaptic communication plays a role whenever rapid changes in neural activity are necessary. However, non-synaptic or paracrine communication has been proposed as a mechanism to coordinate changes in neuronal activity in multiple brain regions, for a prolonged period of time [5, 7–11]. These mechanisms, synaptic and non-synaptic, may serve the same goal, however, and are usually well coordinated, complementary and mutually supportive, as will be discussed later. All changes induced by these mechanisms can be viewed as basic requirements for motivational changes. Many neuropeptidergic fiber systems descend over long distances, are abundantly present in behavior-relevant brain areas, contain mostly unmyelinated varicose fibers and lack synaptic contacts [7–15]. This combination of characteristics puts these fibers in a most favourable position to participate in changing the weight factors in the neural network involved in maintaining or switching between behavioral states [4, 5]. In 1985, Nieuwenhuys [9] wrote (p 197): "I concur with the opinion of Chan-Palay that the periventricular part of the neural extracellular space presumably communicates via ependymal elements with the ventricular system, which in its turn may also represent an important communication channel, acting as a vehicle for widespread distribution of neuroactive substances". From 1986, the term volume transmission has been used for this type of non-synaptic communication [16–21].
Many descending fibers showing paracrine characteristics are observed close to the ventricular system [12, 14, 15, 22], which raises the question as to how far the cerebrospinal fluid (CSF), flowing through the ventricular system and the subarachnoid space surrounding the brain, could be involved in behavioral changes. As early as 1910, Cushing and Goetsch [23] discussed the CSF circulation as a possible way to send messages to distant parts of the brain. Using CSF, obtained from patients with a brain tumour causing obstructive hydrops, strong physiological reactions were observed in rabbits after CSF injections. Later on, Cushing coined the term "third circulation" for the CSF flowing caudally through, as well as around, the CNS. In 1932 Friedman and Friedman investigated the effects of CSF oxytocin on rabbit uterus contractions and noted that the concentrations were effective [24]. Many years later, Borison et al. noted that "the simple concept that the subarachnoid space constitutes a catch basin for interstitial fluid is no longer tenable. Rather, the CSF of that space must be considered as being in dynamic chemical communication with all parts of the central nervous system" [25]. In 1998 a special meeting was held in Los Angeles about a possible role of CSF in spreading the messages and Nicholson reported on signals that go with the flow [26]. In the present review we explore the hypothesis that neuropeptides can be released into the CSF as messages to flow with the fluid, in order to induce changes in motivational states, by influencing appropriate receptors in distant brain areas.
The reviewed data concerning the CSF flow and the distribution of messages support earlier proposals by Sewards and Sewards [27, 28] concerning vasopressin and corticotrophin releasing factor (CRF). In a recent paper, the presence of neuroendocrine signalling molecules in the CSF, led to the conclusion that they play an active role in the function of the nervous system [29], in full agreement with our proposal.
Production and circulation of CSF
Anatomically, the ventricular system of the vertebrate brain develops from a simple elongated cavity inside the neural tube, into a highly complex structure, filled with CSF (Fig 1) [10, 30]. Vigh and others have extensively studied the specialisations and CSF-contacting neurons composing and surrounding the ventricular walls [31–35].

A schematic diagram of structures and specialized cell types bordering the different parts of the mammalian ventricular system, and in contact with the cerebrospinal fluid (CSF). The complexity of the system suggests that CSF functions are not limited to metabolic support of the brain and the release of waste products. Abbreviations: CO: caudal opening of the central canal of the spinal cord;H: hypothalamic CSF-contacting neurons; HY: Hypophysis; LV: lateral ventricle; ME: median eminence; O: vascular organ of the terminal lamina; PIN: pineal organ; R: raphe nuclei; RET: retina; RF: Reissner's fiber; SE: septal region; SCO: subcommissural organ; SP: medullo-spinal CSF-contacting neurons; TEL: telencephalon; TF: terminal filum; (Fig. 1 was kindly provided by Prof. B. Vigh. For details about specific cell types, the reader is referred to: Vigh and Vigh-Teichmann, [31], and to Vigh et al, [32]).
The CSF has an essentially unidirectional flow [36]; caudal through the ventricular system and in different directions through the cisternal subarachnoid spaces surrounding the brain, providing the brain with a protective environment. The CSF can be characterized by its open communication with the extracellular fluid (ECF) of the neuropil, to such an extent that CSF has been described as a reservoir of cerebral ECF [37]. The intercellular junctions between the ependymal cells determine the extent of this communication, and there are many regional differences [32]. In addition, recent investigations suggest that this open communication is biased in the sense that CSF has rapid and widespread access to the ECF, causing local interstitial oedema when CSF-pressure is rising, like after acute hydrocephalus [38, 39]. In the opposite direction, however, only about 10 to 15% of the ECF drains into the CSF [40, 41]. The intercellular junctions in the superficial and perivascular glial limiting membranes may play a role here by providing a separation between subarachnoid and perivascular CSF space and brain ECF.
Therefore, CSF-ECF exchange of neuroactive substances is a variable and complicated process. Concentrations vary over time and in addition there may be large regional concentration differences of substances. Such differences arise following massive local release of substances in the brain, but exist in the CSF as well, where concentrations can vary by a factor of 10 in different regions of the ventricular system [36, 42].
CSF has a high salt concentration (>150 mmol/l) and low protein concentration (ca. 200-700 μg protein/ml). It serves as a transport medium, carrying nutrients for cells and removing products of their metabolism. However, CSF not only contains polypeptides which pass through the blood-brain barrier but also harbours peptides and proteins manufactured locally. The CSF protein population differs in composition from that of plasma due to inherent CSF functions, such as ongoing proteolytic processes involved in cell surface remodeling, protein shedding, and synthesis of regulatory peptides [36, 43, 44]. Blood-derived labelled proteins, administered peripherally, appear first in the lumbar CSF and only later in the cisternal or ventricular CSF [36], while their concentration remains highest in the lumbar subarachnoid space. The additional 20% of the CSF peptides are brain-derived, however, and have the highest concentration in ventricular CSF steadily decreasing along the flow pathway through the subarachnoid space [36]. Apparently, gradients in CSF levels exist and concentration differences of such peptides along the rostrocaudal axis of the central nervous system tell us something about their sources. In summary, the CSF shows an essentially unidirectional flow and may contain brain-derived peptides moving with the flow.
Production and sources of CSF
In the human brain, about 75% of CSF is secreted by the choroid plexuses inside the lateral ventricles [45]. The additional 25% is contributed by the ECF across the ependymal layer [46]. CSF production in the young human brain is about 500 ml/day or about 340 μl/min [40, 45]. This reduces to about 250 ml/day for the elderly, [47]. The adult brain contains a constant volume of about 140 ml of CSF, of which 30 ml occurs in the ventricles and about 110 ml in the subarachnoid spaces [40]. Therefore, human CSF is completely replaced about three times every 24 hours. In the young adult rat, which has about 200 μl CSF, it takes about 2 hours to replace the CSF whereas in 30 month-old rats with 300 μl, it may take up to 8 hours for complete replacement [48].
The production of CSF is under control of factors working on both sides of the choroid plexus [29]. On the vascular or basal side of the epithelium, noradrenergic and cholinergic nerves may exert direct control, diminishing or increasing production, respectively, as reviewed by Perez-Figares et al [46]. On the ventricular side substances present in the CSF may influence receptors on the apical surface of the choroid plexus epithelium. Serotonin, for example, is probably released into the CSF by the supraependymal plexus (see below) and regulates CSF production via binding to 5-HT2C receptors, thereby reducing CSF secretion. Similar mechanisms have been proposed to explain the effects of dopamine, norepinephrine, melatonin and several peptides, among them vasopressin (See [46] for review).
In addition, the composition of CSF is dependent on the activity of surrounding brain tissue [29, 36] and on the activity of a specific circumventricular (CVO) organ, the subcommissural organ and Reissner's fiber complex (Fig 1) [49, 50]. Reissner's fiber develops and extends from the subcommissural organ in a caudal direction, eventually reaching and even extending from the caudal tip of the central canal of the spinal cord (Fig 1) [51, 52]. In the adult human brain, the subcommissural organ and Reissner's fiber can no longer be detected [52], suggesting that the fiber plays a special role during the development of the mammalian CNS [50]. This fiber is composed of aggregated glycoproteins and has the capacity to bind substances present in the CSF and most probably affects the clearance of monoamines from the CSF [53, 54].
The subcommissural organ itself is under serotonergic control [55], while the choroid plexus is under control of the autonomic nervous system, which in turn is controlled by a number of hypothalamic regions. These circumstances make it clear that CSF production and composition is under the influence of an extensive set of regulatory brain mechanisms. In addition, changes in CSF- and intracranial pressure or a developing hydrocephalus seem to be involved in regulatory feedback control mechanisms [56]. In summary, both the production and the composition of the CSF are controlled by a variety of mechanisms.
Flow and destiny of the CSF
CSF is constantly flowing inside the central nervous system, driven by the arterio-venous pressure gradient and the secretory processes at the choroid plexuses [36]. Two different kinds of flow, bulk flow and laminar flow, have to be distinguished, since the mechanisms are very different and probably subservient to very different functions.
The bulk of the CSF flows in a caudal direction via the third ventricle and the cerebral aqueduct to the fourth ventricle. From there it flows either via the lateral apertures of Luschka in all sub-primate vertebrates, and via the additional median aperture of Magendie in primates only [57] into the subarachnoid space including cisternae magna and pontis, surrounding the brainstem [58, 59]. Some flow may continue through the central canal of the spinal cord but most of the subarachnoid CSF flow apparently divides in the cisterna magna into a cortical, rostrally directed, and a lumbar, caudally directed, component [36]. Inside the ventricular system, turbulences can occur at the entrance to the narrowest parts such as the cerebral aqueduct and central canal, which may hinder the flow of the CSF. Such turbulences are probably reduced by the presence of Reissner's fiber [60]. In the central canal of the rat spinal cord, CSF has been observed to continue flowing caudally at a speed of about 1 cm/min [60]. At the caudal tip of the spinal cord in the filum terminale, one opening (rat) or several openings (guinea pig, rabbit) (Fig 1) allow the CSF to access the subarachnoid space [61]. The CSF surrounding the spinal cord may either rapidly re-enter the spinal cord and central canal via perivascular spaces [62], or leave perispinal spaces to be released into lymphatic or venous vessels [63]. Recent human studies suggest that spinal CSF absorption via perispinal spaces alongside the exiting spinal nerves may account for one third of total CSF absorbed in the resting state and considerably more in active individuals [45].
Concerning the cranial CSF surrounding the brain, most textbooks tell us that arachnoid granulations, penetrating the dura and extending into the superior sagittal sinus, compose the main outlet system for CSF absorption in the human. Recent animal investigations, however, have shown that at least 50% and perhaps up to 80% or more of the CSF leaves the cranial cavity through the cribriform plate, via the perineuronal spaces surrounding the olfactory nerves, to drain into the nasal and cervical lymphatics (Fig 2) [64–70]. This rostral outflow of CSF contains considerable amounts of interstitial fluid, especially from midbrain areas [71, 72], and plays an important role in CNS immune system interactions [68, 73].

A schematic diagram showing the direction of the CSF bulk flow in the mammalian brain based on a midsagittal section of the rat brain, kindly provided by Prof. L.W. Swanson. The following regions are indicated: telencephalon (pink shades); diencephalon (interbrain) and brainstem areas (yellow shades); cerebellum dorsal to the brainstem (light blue); sectioned fiber tracts (black). CSF (blue arrows) flows from the lateral ventricles to the third ventricle via the interventricular foramen (IVF), and flows caudally along the dorsal and ventral side of thalamic adhesion, to the cerebral aqueduct (AQ) and fourth ventricle (V4). Some CSF may continue flowing caudally through the central canal of the spinal cord (CC), but most leaves the ventricular system via the lateral apertures and flows through the subarachnoid space, surrounding the brain. This external flow is indicated here along the dorsal and ventral side of the brain but occurs also along all other external brain surfaces. The destination of the subarachnoid flow is the cribriform plate of the ethmoidal bone, containing the penetrating olfactory fibers, where CSF is released in small lymphatic vessels. Additional abbreviations: V3(p, h, m, t, pi): regions and recesses of the third ventricle; Circumventricular organs: 1: subfornical organ; 2: organum vasculosum of the lamina terminalis; 3: median eminence; 4: subcommissural organ; 5: pineal organ; 6: area postrema; AL, IL and NL: different lobes of pituitary; Fiber bundles crossing the midline, coloured black, are not relevant for the present review.
The flow of CSF is indicated in Fig 2. Additional CSF absorption pathways may exist alongside the vessels of the cavernous sinus and via small lymphatic vessels emerging from the perineuronal space of other cranial nerves, like the trigeminal nerve [69]. These cranial CSF exits operate in parallel to the spinal CSF exits to the lymphatic system, mentioned before [63]. Bringing these data together, it seems to be reasonable to hypothesize that of all CSF produced continuously in the human brain, about one third leaves via the spinal vessels and one third via the nasal lymphatics, which leaves about one third for absorption through the arachnoid granulations into the superior sagittal sinus. The balance between these absorption pathways may vary considerably, however, depending on body position, physical activity and other possible factors.
The driving force of the CSF bulk flow is actively supported by arterial pulsations as proposed by Bering [74], in the choroid plexus or by arterial and brain expansion mechanisms [75–78]. A decade ago, these temporarily neglected expansion mechanisms started to receive attention again because of potential clinical relevance [46, 79–83] and age related changes [84].
The laminar flow of CSF occurs in a thin layer along the ventricular walls and is not necessarily restricted to directions from rostral to caudal. Directional beating of ependymal cell cilia lining the ventricular wall, seems to be the most important factor for this flow [60, 85, 86]. The ciliary movements may contribute to the mixing of the CSF, are supported by a sialic acid-induced hydration mantle on the ventricular surfaces and are stimulated by release of serotonin from the supraependymal serotonergic terminals. In addition to transport of neuroactive substances, a rostrally-directed laminar flow may be crucial for newborn neurons to reach their final location (see below). Both bulk and laminar flow seem to be prerequisites for normal CSF functioning, since disturbance of either the ciliary movements or Reissner's fiber both perturb CSF flow to such an extent that hydrocephalus develops [87, 88].
In summary, the flow of the CSF comprises two different mechanisms. Bulk flow is driven by arterio-venous pressure gradients and arterial pulsations, traversing the ventricular system in a caudal direction towards the brainstem apertures. From there the subarachnoid CSF flow can be described as bidirectional: either descending to the spinal cord or ascending to the dorsal and rostral parts of the brain. Since in the latter, cranial, compartment the main exit of the CSF appears to be along the olfactory nerves through the cribriform plate, there must be an important rostrally directed vector in this subarachnoid CSF flow. The laminar flow occurs only inside the ventricular system in a very thin layer along the walls in a variety of directions and driven by the beating of the cilia of local ependymal cells. Interruption of either bulk or laminar flow results in hydrocephalus.
The CSF circulation during development and aging of the CNS
Recently, it was shown in mice that ependymal ciliary beating, inducing laminar flow, is a prerequisite for the migration of new neurons from the subventricular zone to the anteriorly located olfactory bulb [89]. Even in the adult human brain, newly formed neuroblasts follow this path between the subventricular zone and the olfactory bulb. This path is referred to as the rostral migratory stream [90]. Apparently, supported by gradients of brain-derived neurotrophic factor (BDNF) levels in the CSF [91], the neuroblasts know how to go with the laminar flow.
Experimental evidence suggests that during development of the brain, messages from specific parts of the ventricular system are essential for the formation of the layered structure of the cortex [92]. This suggests that specific messages released into the CSF at a specific time point, influence not only functional but also important structural aspects of distant target areas. In addition, such findings suggest that the targets for CSF messages may not be limited to the periventricular areas inside the brain but may be present at the outer surface of the brain as well.
The features of the walls lining the ventricular system also change over time. In the rat brain, ependymal cells appear from gestational day 16 [93]. The function of the ependymocytes as well as the permeability of the ependyma is age-dependent [94–96] and contributes to the changes with age in composition of the CSF [48, 96]. In elderly humans CSF production and flow rate decrease, affecting protein concentrations [36] demonstrating that clear functional changes occur in the third circulation with increasing age.
Clinical evidence shows that changes occurring in the composition of the CSF and in the homeostatic balance of freely exchanging CSF and ECF compartments with age [36], are possibly involved in brain diseases such as Alzheimer's [48, 97]. On the one hand, such changes in the composition of the CSF may profoundly affect the paracrine messages transmitted via the CSF. On the other hand, it may facilitate new opportunities for the treatment of brain diseases as prospects are improved for drug delivery and brain targeting via the CSF for multiple brain diseases [98]. In summary, CSF flow plays an important role in the structural development of the CNS. The laminar flow is involved in the guidance of neuroblasts and the bulk flow in delivering messages obtained from the brainstem to guide cortical development. The composition of CSF and CSF-ECF exchange are profoundly affected by the aging process and may be related to brain diseases, suggesting an opportunity for treatment by targeting drugs via the CSF.
CSF composition, sources, targets and exchange with ECF
Sources of CSF contents
In addition to blood-derived proteins in the CSF (approximately 80%), the other proteins (about 20%) originate from within the CNS, from neurons, glial and leptomeningeal cells [36]. The choroid plexus itself synthesizes proteins, some of them abundantly, such as transthyretin, a carrier for thyroid hormones. Once secreted into the CSF, such proteins are thought to influence distant parts of the brain [99]. The direct or indirect activation of brain areas bordering the ventricular system or subarachnoid space has been shown to result in increased levels of neuropeptides released into the CSF [100–102]. The circadian variations in CSF concentrations without concomitant changes in peripheral concentrations provide additional evidence for selective release of neuropeptides such as oxytocin into the CSF [103–106]. Specific dendritic release mechanisms occur in brain areas such as the paraventricular and supraoptic hypothalamic nuclei, leading to as much as a thousand fold increase in the local ECF peptide concentrations [107]. Experiments involving ablation and transplantation of the suprachiasmatic nucleus in the golden hamster showed that the release of a diffusible signal was most likely carried by the flow of CSF [108, 109]. Recently, it has been shown that in the diurnal grass rat, the suprachiasmatic nucleus releases at least some of its contents into the CSF in order to reach specific target areas [110]. Taking together the available evidence, we suggest that the active release of neuropeptides into the CSF has to be considered as a probable mechanism for message transmission via the CSF to distant brain areas, as was previously discussed during a special conference in 1998 [26].
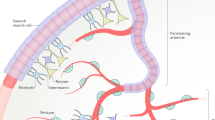
In addition to the neuronal release from specific brain areas, the presence of widespread supraependymal cell clusters, fibers and plexuses of neuronal and other cellular elements, including macrophages, have been described since the early seventies [32, 111]. Their abundant presence near the median eminence suggests that these supraependymal elements are involved in a variety of functions [112–117] (Fig 3A, B). Among these, serotonergic fibers have been described as originating from the raphe nuclei with nerve terminals located in the CSF. These fibers are thought to release their content differentially from several parts of the ventricular walls of the mammalian ventricular system into the flowing CSF [111, 112, 117–121] (Fig 3C).

(A and B) show scanning electron microscopy of supraependymal structures in the rat brain. A: Partially ensheathed fibers and terminal arborizations; B: cellular structures with short fiber extensions, on the ventricular surface of the ependymal cells; Fig 3C: a schematic diagram of the ependymal layer with supra- and subependymal cellular structures contacting the CSF at the ventricular side of the layer; the * indicates a supraependymal terminal extending into the CSF, as described for serotonergic fibers. Figs 3A and 3B were reprinted from [121], with permission of Springer-Verlag. Fig 3C was kindly provided by Prof. B. Vigh.
In addition to the supraependymal elements, many fibers containing specific neuroactive substances can be observed in the subependymal layer. Many of these are of hypothalamic origin and may contain peptides like luteinising hormone releasing hormone (LHRH), CRF or adreno-corticotropic hormone (ACTH) [12, 14, 122]. Others originate in the dorsal periaqueductal gray to descend towards the cervical and upper thoracic spinal cord, many of them contacting the perivascular spaces [22]. Many of these descending fibers are characterized by large numbers of varicosities or local swellings containing numerous vesicles but without synaptic specialisations. The presence of such varicosities suggests that a single fiber may release its contents over large distances, by non-synaptic release or exocytosis, either into the CSF or for diffusion into the bordering ECF to influence a population of receptive neurons [12, 14, 31, 32].
From the available data, no clear general function can be derived for these supra- and sub-ependymal elements. However, their differential distribution over specific regions of the ventricular walls strongly suggests two possibilities: local effects and/or specific effects on receptive distant target areas. Two interesting cases of substances released into the third ventricle, going with the flow to reach their target areas, have been studied in detail so far: melatonin and the LHRH system, also known as the gonadotropin-releasing hormone (GnRH) system.
The pineal gland produces the hormone melatonin at night. The gland is located dorsal to the third ventricle and has access to the CSF via the pineal recess [123]. In a series of experiments in sheep, it was shown that the main effects of the pineal gland occurred through release of melatonin into the dorsal tip of the third ventricle. At the exit of the pineal gland, concentrations were about 10 times higher than in the rest of the ventricular system and about 100 times higher than in peripheral blood [123, 124]. Melatonin is not only secreted into the CSF but also into the general circulation [124, 125], which makes the situation more complex. Melatonin also regulates the CSF levels of another hormone, estradiol [126], which may result in additional indirect effects. However, its main site of action to control reproduction is located in the premammillary hypothalamic nucleus, where it stimulates luteinizing hormone secretion in the ewe [127]. Apparently, transport mechanisms from the pineal gland via the CSF to the premammillary hypothalamus are involved.
Another example is posed by the neuropeptide: LHRH also known as GnRH. This neuropeptide is released in a pulsatile manner into the portal system and stimulates the pulsatile gonadal hormone production of the pituitary [128, 129]. However, GnRH is released into the CSF as well, and this CSF GnRH neither functions as a feedback system for controlling GnRH production, nor does it directly affect the pituitary gland [130]. The influx of peripheral GnRH across the blood-brain barrier into the CSF is so limited that it cannot explain the observed levels of CSF GnRH which are comparable to those in the portal blood stream [131]. These findings, combined with detailed measurement of GnRH concentrations at different sites in the ventricular system suggest that GnRH is released directly into the CSF and the third ventricle. This occurs at the level of the median eminence, and possibly the organum vasculosum of the lamina terminalis (OVLT) (indicated as ME and O in Fig 1) where GnRH immunoreactive (IR) perikarya and axons are very abundant [128, 131]. The authors conclude that the function of CSF GnRH expresses itself in behavioral effects, induced by volume transmission via the flow of CSF to affect distant brain areas, such as periaqueductal gray (PAG) and hippocampal regions. These regions contain, indeed, high densities of GnRH receptor expressing neurons, while GnRH-IR axons are either scarce (PAG) or completely lacking [12, 15, 131]. The PAG is strongly involved in sexual behavior [132–135] which can be elicited by intraventricular (icv) but not by intravenous administration of GnRH [136]. These findings concerning the distribution and behavioral effects of melatonin and GnRH via the CSF comprise, in our view, compelling evidence for CSF messages controlling aspects of behavior.
That the exact site of release of a neuropeptide into the ventricular system is the determining factor for its given effects, has been shown in experiments involving transplants of the suprachiasmatic nucleus in the hypothalamus [109, 137]. Research aimed at identifying the mechanisms regulating food intake, such as experiments involving delivery of substances into either the third or the fourth ventricle, with an open or an experimentally-closed cerebral aqueduct, have shown that caudally-flowing substances and neuropeptides (glucose, leptin, neuropeptide tyrosine (NPY) reach specific receptive areas to influence food intake [138–140]. Apparently, most factors, influencing food intake, like cocaine- and amphetamine-regulated transcript (CART), bombesin and oxytocin, exert their effects in the caudal brainstem [141–144], while other factors are only effective in or around the third ventricle (NPY and calcitonin [140, 145].
In summary, about 80% of the CSF proteins are derived from the blood [146] while about 20% are produced by the brain. Brain areas bordering the ventricular system as well as supra- and subependymal cells and fibers lining the ventricular walls are in the most favourable position to add such substances to the CSF. Melatonin and GnRH, both released into the third ventricle but at different locations, and the experiments concerning substances controlling food intake administered into either the third or the fourth ventricle, and moving with the flow, are good examples for local additions into the CSF influencing remote brain areas. Apparently, the site of upstream release determines which downstream areas will be affected.
Flow-transport: speed and mechanisms
The question can be raised whether the flow-transport mechanisms work rapidly enough to induce behavioral effects in a relevant timescale. A quick survey of the literature shows that the time between administration or release of neuropeptides into the CSF and the observed behavioral or physiological effects turns out to be very short. In a series of experiments involving respiratory and cardiovascular control mechanisms in the cat, effects of icv administration of respiratory stimulants such as vasopressin into the lateral ventricle were observed within two minutes at the ventral surface of the brainstem. Since this surface lies beyond the foramina of Luschka, this finding indicates that the stimulus was moving with the CSF flow in the cat brain at a speed of more than 1 cm/min [57]. A tracer like HRP carefully administered into the lateral ventricle of the cat in order to avoid direct pressure effects, penetrates the outer surface of the brainstem within four or five minutes (see below) [25]. A similar rapid distribution of a tracer [14C]Inulin as well as of [125I]CRH after administration into the lateral ventricle has been observed in the rat [147]. Other data obtained from the rat also show rapid flow transport of neuropeptides. α-Melanocyte stimulating hormone (αMSH) injected into the lateral ventricle appeared within two minutes in the cisterna magna [148]. β-endorphin was observed in the CSF within minutes after electrical microstimulation of the arcuate hypothalamic nucleus, without any change in peripheral levels [100, 149]. Both interleukin-1 and CRF have been shown to induce widespread effects throughout the brain, starting a few minutes after icv administration [150, 151]. The effects of icv oxytocin on ventromedial hypothalamic neurons is manifest in under one minute [152]. Fenstermacher and his group showed a similar rapid distribution via the ventricular system, however their findings revealed that the flow over the cortical surface was considerably slower [146, 153, 154].
Recent measurements of the pulsatile flow rates in the cerebral aqueduct of the human brain by modern imaging and other techniques fully support the high speed of this CSF transport channel [75, 78, 83]. Maximal flow rates of 12 mm/s and higher were observed in the aqueduct [58, 59], while at other sites like the spinal canal, the flow rate was almost negligible [58].
In summary, transport of CSF contents by bulk flow is fast, working on a time scale of minutes, and is directly relevant for behavioral and physiological adaptations.
Exchange of substances between CSF and ECF: mechanisms and targets
Exchange of fluid contents between the ECF in the neuropile and the ventricular CSF occurs via different mechanisms and at different rates: 1: diffusion, 2: perivascular pump mechanisms, and 3: tanycytes and ECF-contacting neurons.
1: Diffusion. This is a rather slow process, depending on ECF diffusion parameters like volume fraction, tortuosity and the diffusion coefficient of the neuroactive substance [155–157]. Diffusion in the ECF compartment may appear anisotropic, due to the hindrances imposed by local differences in tortuosity [155]. White matter tracts are guiding tracts for the interstitial fluid flow as well as for the dissemination of primary brain tumors [158]. Diffusion and exchange between cerebrospinal and extracellular fluid compartments may occur in both directions [37, 77] through an uninterrupted single-layered lining of the ventricular system, consisting of several types of ciliated ependymal cells [95, 159]. Due to the lack of tight junctions between ependymal cells, the CSF-ECF barrier can be described as open or non-existent. In that respect, the CVO's form a notable exception with a closed CSF-ECF barrier and an open blood-ECF barrier (usually called blood-brain barrier, or BBB), (see McKinley et al[160]). Interestingly from a cybernetic point of view, the open CSF-ECF compartment throughout the brain is functionally separated from the open BBB-ECF compartment inside the CVO's. These two compartments can be described as being disjoint or even complementary, but the direct neuroanatomical relationships between the paraventricular hypothalamic nucleus and the CVO's [161–168] manage the bidirectional communication and mutual tuning of both compartments.
In general, concentration differences and gradients determine the direction of the diffusion between CSF and ECF. High levels of neuroactive substances in the CSF will enter the ECF, while high local concentrations in the ECF will diffuse into the CSF. For brain areas like the medial hypothalamus, as well as the periaqueductal gray, such diffusion processes will result in continuous equilibration processes in the levels of neuroactive substances involved in the induction or maintenance of behavioral states. Widely divergent substances and techniques have been used to measure diffusion distances. Diffusion over distances of 0.5 mm and at other sites up to 1 mm away from the wall of the third ventricle, from the cerebral aqueduct and from the floor of the fourth ventricle [25, 147, 151, 169, 170] are reasonable estimations. In rat brain, areas such as the lateral septum and parts of the bed nucleus of the stria terminalis, the dorsal midline thalamus and habenular nuclei, the medial hypothalamic regions, the mesencephalic periaqueductal gray as well as several brainstem core and paracore areas [7] can, therefore, be influenced directly by the composition of the CSF. Diffusion of any relevant substance from the CSF into these regions would influence a number of behavior-relevant brain functions [27].
Diffusion in the other direction, that is from the ECF into the CSF occurs as well and may also play a role over larger distances. This situation was observed after infusion of a small amount of beta-endorphin into the striatum of the rat [171]. Despite the considerable distance to the ventricular system, the infused beta-endorphin appeared in the CSF within 10 minutes, reaching peak values in the cisterna magna after about 30 minutes [171]. This experiment shows that diffusion within the brain may occur over considerable distances and in a behaviour-relevant time-scale during the CSF-ECF exchange processes.
On the other hand, many different substances have been injected into the ventricular system of a variety of vertebrates and a speed and depth of penetration has been observed which far exceeds the capabilities of diffusion. Apparently, active transport processes are involved as well. Two of these will be described.
2: Perivascular pump mechanisms have been shown to play an important role in the distribution of CSF contents. The pump involves the perivascular space of vessels penetrating the outer surface of the brain and is driven by the arterial pulsations of the cerebral vessels [172, 173]. The differences between the passive diffusion processes are striking. While the diffusion rate of the neuroanatomical tracer horseradish-peroxidase (HRP) in brain tissue amounts to less than 1 mm/hr, after an intraventricular or cisternal injection, HRP can be observed along all microvessels of the neuraxis. HRP molecules penetrated deeply (several mm) within five minutes [172]. This fast spread occurs along perivascular spaces and is aided by a paravascular fluid circulation system, driven by the pulsations of the cerebral arteries. In parallel to an inward flow along the arterial perivascular spaces, an outward flow may occur along the venous paravascular routes, adding interstitial fluid to the ECF/CSF in the subarachnoid space [172, 174]. The existence of these paravascular routes provides the neuroactive CSF contents with a widespread and fast portal to enter less superficial brain areas and to affect a much larger variety of regions in brain or spinal cord [62, 175, 176]. A more recent study, in which the tracer [14C]inulin was used, confirmed the rapid and extensive distribution of icv-injected substances. Interestingly, it was concluded that the paravascular fluid circulation may even preferentially target specific brain areas like the hippocampus, the basal forebrain including the hypothalamus and the ventral brainstem surface [147]. This perivascular pump has been described as a "powerful mechanism for the distribution of therapeutic molecules within the brain" [173]. Similar mechanisms may support neuropeptides released into the CSF to reach remote destinations. GnRH, for example, could reach the hippocampal regions, which are richly supplied with GnRH receptors while a GnRH innervation is virtually lacking [131].
3: In addition to the slow diffusion mechanisms for local/regional effects and the rapid perivascular pump mechanisms, involving the whole or some preferential large parts of the brain, special ependymal cells (tanycytes) and neurons contact the CSF in order to internalize specific constituents, to release them at specific sites and for specific purposes, and are not covered by the general mechanisms mentioned before.
Tanycytes compose a specific group of ependymal cells. Those lining the ventral part of the third ventricle have basal processes that extend to the hypophyseal-portal vasculature as well as into the mediobasal hypothalamus, suggesting a role in pituitary and neural control. As reviewed by Bruni [95] and Rodriguez et al. [177], multiple tanycyte functions have been proposed over time. Tanycytes can be observed also in the walls of the lateral and fourth ventricles as well as along the central canal of the spinal cord [178]. Tanycyte extensions apparently radiate from the third ventricle into hypothalamic nuclei, but also laterally into the caudate putamen or, from the floor of the fourth ventricle, ventrally into the lower brainstem areas [178]. Several types of tanycytes have been described with specifically different endocytotic mechanisms [179, 180]. Some of these recent experiments compared peripheral and icv administration of tracers and neuropeptides and the distribution and tanycyte-labelling patterns inside the brain were very different [179, 181]. This suggests that tanycytes of a given type are specifically selective, each one retrieving a different substance from the CSF and underlining the highly specific nature of tanycyte messages. Our understanding of the functional effects tanycyte messages is only preliminary and major steps for elucidating their role, remain to be taken.
Very recently, melanocortin-concentrating-hormone-(MCH) immunoreactive ependymal cells, extending between the floor of the 4th ventricle and the midbrain raphe nuclei have been reported [182]. The authors concluded that these elongated tanycytes internalized MCH released from the hypothalamus into the CSF, to act on specific target neurons in the dorsal raphe nucleus. Since serotonergic neurons are numerous in this midbrain raphe nucleus, internalization of MCH followed by release from these specific tanycytes may have far-reaching effects on serotonergic transmission and therefore on behavior.
Additionally, neurons may directly contact the CSF [31, 32]. Such neurons have been observed at a considerable distance from the ventricular system. Some use selective receptor-mediated uptake mechanisms to obtain specific substances from the CSF and have been observed in brain areas such as the hippocampus, septum and cortex [170]. Other CSF-contacting neurons were observed in anterodorsal thalamus, supramammillary nucleus, dorsal raphe nucleus, the floor of the fourth ventricle as well as in the lateral superior olive nucleus [178]. This implies, for example, that if the supramammillary region is functionally affected by the CSF contents, the activity of the whole septohippocampal complex is involved, linking functional (learning) and behavioral (fear/anxiety) consequences to the composition of the CSF. In particular, the dorsal raphe nucleus seems to have strong links with the CSF, with several CSF-contacting groups of neurons [178, 183] in addition to the special tanycytes, mentioned before. Recently, it was shown that the locus coeruleus accumulates about 50% of a nerve growth factor, not by neuronal transport mechanisms but directly from the CSF [184]. Taken together with the special MCH-transmitting ependymal cells, [182], the activity of the raphe neurons or the serotonergic system, as well as the locus coeruleus neurons or the noradrenergic system, are under control of factors circulating in the CSF. Considering that most of these findings are very recent, it is only to be expected that during the next few years more brain areas and more functional and transmitter systems will be discovered that derive part of their controlling information from CSF.
In summary, the interaction between the CSF and its contents occurs in three different but complementary ways: 1) diffusion: a slow and local process, over short distances, 2) via perivascular spaces and an active overall pump system driven by the vascular system, and 3) specific uptake mechanisms: depending on the receptors exposed to the CSF.
The existence of these mechanisms fully supports the original notion of Borison et al, (1980) that "the simple concept that the subarachnoid space constitutes a catch basin for interstitial fluid is no longer tenable. Rather, the CSF of that space must be considered as being in dynamic chemical communication with all parts of the central nervous system" [25]. These mechanisms are in full agreement with the conclusions drawn by Abbott after a careful discussion of available data about interstitial fluid (ISF = ECF) flow, including the CSF [185] "Chemical signals produced within the brain are able to use the flowing ECF (= ISF) as a communication route", with rapid bulk flow occurring within the CSF system, ... "or vectorial signalling from upstream to downstream sites". These mechanisms are ... "extending the sphere of influence of a bioactive molecule beyond that which could be achieved by diffusion"... "and provide the basis for a site specific communication pathway, by which molecules released from site A could be specifically designed to be carried to and influence site B" (pp 550,551). Specificity and range would be determined by factors like the anatomy of the communication route, by the presence of appropriate receptors and by activation mechanisms to limit or confine the signal spread [185]. Abbott (2004) concludes that "the ability to predict the effects of ECF/ISF flow ... is likely to play an important role in developing future therapeutic strategies for CNS targeting and repair".
Neuropeptides form an integral part of the CSF composition
In the preceding sections, it has been mentioned that many neuropeptides have been found in the CSF. For many of them, CSF levels bear hardly any relationship to peripheral levels in the blood, and they are actively secreted by choroid plexus, glial and neuronal cells into the CSF [36]. Active release of neuropeptides from brain areas bordering the ventricular system with the aim or at least the effect of influencing downstream brain areas at a distance, has been repeatedly observed for a range of different neuropeptides and transmitters [12, 14, 36, 42, 99, 102, 108, 123, 129–131, 136, 184, 186–188]. Sewards and Sewards [28] discussed extensively the evidence available for the neuropeptides vasopressin and corticotrophin-releasing-hormone, and concluded that these were actively secreted into the CSF to influence downstream receptive areas involved in specific kinds of behavior.
An interesting recent paper [189] showed the existence of numerous nanoparticles in the CSF, composing a "matrix of membrane- and protein rich nano-scaled structures with many transduction components bounded by lipid membranes". They concluded that this "bulk flow of nanostructures generates a more dispersed signal delivery, of longer duration", in order to "regulate brain behaviors known to require slower, more gradual, and more sustained modulations, such as reported for sleep, appetite, mood and vasomotor regulation". This conclusion is in full agreement with our hypothesis as proposed in the present review.
In summary, there are convincing indications for neuropeptide release into the CSF, and for transport via the CSF bulk flow as a special kind of volume transmission towards receptive brain areas, to propose a message function for specific CSF contents.
Behavioral effects of CSF contents
The question that remains is: what are the functional significances and the specific contributions of such relatively uncontrolled and broad-spectrum CSF-release systems, in addition to precisely targeted synaptic or more diffuse non-synaptic release mechanisms? Many instances have been discussed above such as melatonin, GnRH, feeding experiments with administration into the 3rd or 4th ventricle, that provide evidence for peptidergic and other signals that arrive at their target areas via the CSF at a concentration level sufficient to induce obvious behavioral changes. Taken together, with the many papers reporting that administration of substances into CSF is much more effective than peripheral administration for inducing behavioral changes, we judge the evidence as overwhelming for the statement that the CSF (the third circulation) contains meaningful messages to be recognized by the appropriate sensitive brain target areas to elicit a neural and physiological/behavioral response. Therefore, we support the view that broadcasting messages via the CSF is an appropriate way to put several parts of the CNS into a behavioral state. Such a brain state is subservient to the performance of specific behaviors or groups of behavioral elements that share a common motivational system and may include the temporary suppression of specific behavioral or physiological reactions. These CSF messages have to be considered as a partially-independent signal. Simultaneously, these messages are constantly supporting, sustaining and in mutual exchange with the ECF contents released by axon terminals all over the brain and specifically in brain areas adjoining the CSF.
Sewards and Sewards [28] proposed that the presence of elevated CSF levels of CRF are involved in the motivation of fear while the presence of vasopressin is involved in the power-dominance drive motivation. They hypothesized that the elevated neuropeptide levels in the CSF are detected and transduced into neuronal activities by, predominantly, hypothalamic neurons in the vicinity of the third ventricle. The purpose of this system was proposed to maintain a state of fear or anger and consequent vigilant or aggressive behavior, after the initial fear- or anger-inducing stimulus itself is no longer perceptible [27]. In other words, CSF messages prolong the duration of a motivated state. Interestingly, important destination areas for this transmission (the medial hypothalamus, periaqueductal gray, midline thalamus and medial) prefrontal cortex are all bordering the CSF system, at the inner or outer surface of the brain. In addition, each of these destination areas is extensively connected to other parts of the limbic system and functionally involved in a range of differently motivated behaviors [27].
The time scale of these effects, starting within minutes but extending over time [57] up to more than an hour [190, 191] are fully appropriate to induce and sustain behavioral changes reflecting a modified behavioral state of the animal. The anatomical observations of many varicose peptidergic fibers present close to and in parallel to the ventricular system [12–15, 22] suggests that messages arriving via the axons by terminal and varicosity release may be supported by messages arriving via the CSF in a mutually sustaining way, because of the free exchange between ECF and CSF. Such sustaining CSF messages in harmonic balance with neuronal communication mechanisms are in a perfect position to play a role in the rise and decline of successive motivational states and for necessary transitional states.
Conclusions
There is considerable evidence that peptidergic and other substances are actively secreted into and distributed via the CSF spreading with surprising speed to many brain areas. We propose that they can be considered functionally as messages, because their presence can influence the activity of specific groups of receptive neurons, located near the ventricular and subarachnoid surfaces of the brain, downstream from the site of release. As such messages remain active considerably longer than a few seconds, they may form part of the mechanisms underlying the changes in successive behavioural states.
References
Frijda N: The laws of emotions. 2007, London: Psychology Press
Hinde RA: Animal Behaviour; A Synthesis of Ethology and Comparative Psychology. 1970, New York: McGRAW-Hill Book Cy, second
Mogenson GJ: The neurobiology of Behavior: an introduction. 1977, Hillsdale: Lawrence Erlbaum Associates
Panksepp J: Affective Neuroscience, The foundations of human and animal emotions. 1998, New York: Oxford University Press
Swanson LW: Brain Architecture, understanding the basic plan. 2003, New York: Oxford University Press
Barendregt H: Buddhist Phenomenology (Part 1). Conference on Topics and Perspectives of Contemporary Logic and Philosophy of Science; Cesena, Italy. Edited by: dalla Chiara M. 1988, Clueb, Bologna, 37-55.
Nieuwenhuys R, Veening JG, van Domburg P: Core and paracores; some new chemoarchitectural entities in the mammalian neuraxis. Acta Morphol Neerl Scand. 1988, 26: 131-163.
Veening JG: Brain and behaviour: morphological and functional aspects of the hypothalamus in the rat. Eur J Morphol. 1992, 30: 53-66.
Nieuwenhuys R: Chemoarchitecture of the Brain. 1985, Springer-Verlag, Berlin
Nieuwenhuys R, Voogd J, van Huijzen C: The Human Central Nervous System. 2008, Berlin: Springer, 4
Nieuwenhuys R: Comparative aspects of volume transmission, with sidelight on other forms of intercellular communication. Prog Brain Res. 2000, 125: 49-126. full_text.
Buma P: Characterization of luteinizing hormone-releasing hormone fibres in the mesencephalic central grey substance of the rat. Neuroendocrinology. 1989, 49: 623-630. 10.1159/000125179.
Buma P, Nieuwenhuys R: Ultrastructural demonstration of oxytocin and vasopressin release sites in the neural lobe and median eminence of the rat by tannic acid and immunogold methods. Neurosci Lett. 1987, 74: 151-157. 10.1016/0304-3940(87)90141-8.
Buma P, Veening J, Hafmans T, Joosten H, Nieuwenhuys R: Ultrastructure of the periaqueductal grey matter of the rat: an electron microscopical and horseradish peroxidase study. J Comp Neurol. 1992, 319: 519-535. 10.1002/cne.903190405.
Veening JG, Buma P, ter Horst GJ, Roeling TAP, Luiten PGM, Nieuwenhuys R: Hypothalamic projections to the PAG in the rat: topographical, immunoelectronmicroscopical and functional aspects. The Midbrain Periaqueductal Gray Matter. Edited by: Depaulis A, Bandler R. 1991, New York: Plenum Press, 387-415.
Agnati LF, Bjelke B, Fuxe K: Volume versus wiring transmission in the brain: a new theoretical frame for neuropsychopharmacology. Med Res Rev. 1995, 15: 33-45. 10.1002/med.2610150104.
Agnati LF, Fuxe K: Volume transmission as a key feature of information handling in the central nervous system possible new interpretative value of the Turing's B-type machine. Prog Brain Res. 2000, 125: 3-19. full_text.
Agnati LF, Fuxe K, Zoli M, Ozini I, Toffano G, Ferraguti F: A correlation analysis of the regional distribution of central enkephalin and beta-endorphin immunoreactive terminals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring transmission. Acta Physiol Scand. 1986, 128: 201-207. 10.1111/j.1748-1716.1986.tb07967.x.
Fuxe K, Dahlstrom A, Hoistad M, Marcellino D, Jansson A, Rivera A, Diaz-Cabiale Z, Jacobsen K, Tinner-Staines B, Hagman B, Leo G, Staines W, Guidolin D, Kehr J, Genedani S, Belluardo N, Agnati LF: From the Golgi-Cajal mapping to the transmitter-based characterization of the neuronal networks leading to two modes of brain communication: wiring and volume transmission. Brain Res Rev. 2007, 55: 17-54. 10.1016/j.brainresrev.2007.02.009.
Zoli M, Jansson A, Sykova E, Agnati LF, Fuxe K: Volume transmission in the CNS and its relevance for neuropsychopharmacology. Trends Pharmacol Sci. 1999, 20: 142-150. 10.1016/S0165-6147(99)01343-7.
Zoli M, Torri C, Ferrari R, Jansson A, Zini I, Fuxe K, Agnati LF: The emergence of the volume transmission concept. Brain Res Brain Res Rev. 1998, 26: 136-147. 10.1016/S0165-0173(97)00048-9.
Mouton LJ, Kerstens L, Want Van der J, Holstege G: Dorsal border periaqueductal gray neurons project to the area directly adjacent to the central canal ependyma of the C4-T8 spinal cord in the cat. Exp Brain Res. 1996, 112: 11-23. 10.1007/BF00227173.
Cushing H, Goetsch E: Concerning the secretion of the infundibular lobe of the pituitary body and its presence in the cerebrospinal fluid. Amer J Physiol. 1910, 27: 60-86.
Friedman GS, Friedman MH: An examination of cerebrospinal fluid for oxytocinergic activity as tested by the rabbit uterine fistula. Amer J Physiol. 1932, 103: 244-254.
Borison HL, Borison R, McCarthy LE: Brain stem penetration by horseradish peroxidase from the cerebrospinal fluid spaces in the cat. Exp Neurol. 1980, 69: 271-289. 10.1016/0014-4886(80)90211-3.
Nicholson C: Signals that go with the flow. Trends Neurosci. 1999, 22: 143-145. 10.1016/S0166-2236(98)01388-5.
Sewards TV, Sewards MA: Representations of motivational drives in mesial cortex, medial thalamus, hypothalamus and midbrain. Brain Res Bull. 2003, 61: 25-49. 10.1016/S0361-9230(03)00069-8.
Sewards TV, Sewards MA: Fear and power-dominance motivation: proposed contributions of peptide hormones present in cerebrospinal fluid and plasma. Neurosci Biobehav Rev. 2003, 27: 247-267. 10.1016/S0149-7634(03)00034-4.
Skipor J, Thiery JC: The choroid plexus--cerebrospinal fluid system: undervaluated pathway of neuroendocrine signaling into the brain. Acta Neurobiol Exp (Wars). 2008, 68: 414-428.
McFarland WL, Morgane PJ, Jacobs MS: Ventricular system of the brain of the dolphin, Tursiops truncatus, with comparative anatomical observations and relations to brain specializations. J Comp Neurol. 1969, 135: 275-368. 10.1002/cne.901350304.
Vigh B, Vigh-Teichmann I: Actual problems of the cerebrospinal fluid-contacting neurons. Microsc Res Tech. 1998, 41: 57-83. 10.1002/(SICI)1097-0029(19980401)41:1<57::AID-JEMT6>3.0.CO;2-R.
Vigh B, Manzano e Silva MJ, Frank CL, Vincze C, Czirok SJ, Szabo A, Lukats A, Szel A: The system of cerebrospinal fluid-contacting neurons. Its supposed role in the nonsynaptic signal transmission of the brain. Histol Histopathol. 2004, 19: 607-628.
Calle M, Claassen IE, Veening JG, Kozicz T, Roubos EW, Barendregt HP: Opioid peptides, CRF, and urocortin in cerebrospinal fluid-contacting neurons in Xenopus laevis. Ann N Y Acad Sci. 2005, 1040: 249-252. 10.1196/annals.1327.035.
Calle M: Integrative Physiology and Communication in the Amphibian brain. PhD-Thesis. 2006, Radboud University, Nijmegen, Depts of Cellular animal Physiology/Computational Sciences
Vigh B, Manzano e Silva MJ, Frank CL, David C, Czirok SJ, Vincze C, Racz G, Lukats A, Szel A: The circumventricular organs of the brain: do they represent a cerebrospinal fluid-dependent regulatory system?. Medical Hypotheses and Research. 2004, 1: 77-100.
Reiber H: Proteins in cerebrospinal fluid and blood: barriers, CSF flow rate and source-related dynamics. Restor Neurol Neurosci. 2003, 21: 79-96.
Milhorat TH: The third circulation revisited. J Neurosurg. 1975, 42: 628-645. 10.3171/jns.1975.42.6.0628.
Del Bigio MR, Enno TL: Effect of hydrocephalus on rat brain extracellular compartment. Cerebrospinal Fluid Res. 2008, 5: 12-10.1186/1743-8454-5-12.
Weller RO, Wisniewski H: Histological and ultrastructural changes with experimental hydrocephalus in adult rabbits. Brain. 1969, 92: 819-828. 10.1093/brain/92.4.819.
Weller RO, Djuanda E, Yow HY, Carare RO: Lymphatic drainage of the brain and the pathophysiology of neurological disease. Acta Neuropathol. 2009, 117: 1-14. 10.1007/s00401-008-0457-0.
Szentistvanyi I, Patlak CS, Ellis RA, Cserr HF: Drainage of interstitial fluid from different regions of rat brain. Am J Physiol. 1984, 246: F835-844.
Tricoire H, Moller M, Chemineau P, Malpaux B: Origin of cerebrospinal fluid melatonin and possible function in the integration of photoperiod. Reprod Suppl. 2003, 61: 311-321.
Zougman A, Pilch B, Podtelejnikov A, Kiehntopf M, Schnabel C, Kumar C, Mann M: Integrated Analysis of the Cerebrospinal Fluid Peptidome and Proteome. J Proteome Res. 2008, 7: 386-399. 10.1021/pr070501k.
Yuan X, Desiderio DM: Human cerebrospinal fluid peptidomics. J Mass Spectrom. 2005, 40: 176-181. 10.1002/jms.737.
Edsbagge M, Tisell M, Jacobsson L, Wikkelso C: Spinal CSF absorption in healthy individuals. Am J Physiol Regul Integr Comp Physiol. 2004, 287: R1450-1455.
Perez-Figares JM, Jimenez AJ, Rodriguez EM: Subcommissural organ, cerebrospinal fluid circulation, and hydrocephalus. Microsc Res Tech. 2001, 52: 591-607. 10.1002/1097-0029(20010301)52:5<591::AID-JEMT1043>3.0.CO;2-7.
May C, Kaye JA, Atack JR, Schapiro MB, Friedland RP, Rapoport SI: Cerebrospinal fluid production is reduced in healthy aging. Neurology. 1990, 40: 500-503.
Preston JE: Ageing choroid plexus-cerebrospinal fluid system. Microsc Res Tech. 2001, 52: 31-37. 10.1002/1097-0029(20010101)52:1<31::AID-JEMT5>3.0.CO;2-T.
Perez J, Garrido O, Cifuentes M, Alonso FJ, Estivill-Torrus G, Eller G, Nualart F, Lopez-Avalos MD, Fernandez-Llebrez P, Rodriguez EM: Bovine Reissner's fiber (RF) and the central canal of the spinal cord: an immunocytochemical study using a set of monoclonal antibodies against the RF-glycoproteins. Cell Tissue Res. 1996, 286: 33-42. 10.1007/s004410050672.
Vio K, Rodriguez S, Yulis CR, Oliver C, Rodriguez EM: The subcommissural organ of the rat secretes Reissner's fiber glycoproteins and CSF-soluble proteins reaching the internal and external CSF compartments. Cerebrospinal Fluid Res. 2008, 5: 3-10.1186/1743-8454-5-3.
Kohno K: Electron microscopic studies on Reissner's fiber and the ependymal cells in the spinal cord of the rat. Z Zellforsch Mikrosk Anat. 1969, 94: 565-573. 10.1007/BF00936062.
Nualart F, Hein S: Biosynthesis and molecular biology of the secretory proteins of the subcommissural organ. Microsc Res Tech. 2001, 52: 468-483. 10.1002/1097-0029(20010301)52:5<468::AID-JEMT1033>3.0.CO;2-U.
Rodriguez S, Vio K, Wagner C, Barria M, Navarrete EH, Ramirez VD, Perez-Figares JM, Rodriguez EM: Changes in the cerebrospinal-fluid monoamines in rats with an immunoneutralization of the subcommissural organ-Reissner's fiber complex by maternal delivery of antibodies. Exp Brain Res. 1999, 128: 278-290. 10.1007/s002210050848.
Caprile T, Hein S, Rodriguez S, Montecinos H, Rodriguez E: Reissner fiber binds and transports away monoamines present in the cerebrospinal fluid. Brain Res Mol Brain Res. 2003, 110: 177-192. 10.1016/S0169-328X(02)00565-X.
Jimenez AJ, Fernandez-Llebrez P, Perez-Figares JM: Neural input and neural control of the subcommissural organ. Microsc Res Tech. 2001, 52: 520-533. 10.1002/1097-0029(20010301)52:5<520::AID-JEMT1037>3.0.CO;2-6.
Johanson CE, Duncan JA, Klinge PM, Brinker T, Stopa EG, Silverberg GD: Multiplicity of cerebrospinal fluid functions: New challenges in health and disease. Cerebrospinal Fluid Res. 2008, 5: 10-10.1186/1743-8454-5-10.
Feldberg W: The ventral surface of the brain stem: a scarcely explored region of pharmacological sensitivity. Neuroscience. 1976, 1: 427-441. 10.1016/0306-4522(76)90093-2.
Gupta S, Soellinger M, Boesiger P, Poulikakos D, Kurtcuoglu V: Three-dimensional computational modeling of subject-specific cerebrospinal fluid flow in the subarachnoid space. J Biomech Eng. 2009, 131: 021010-10.1115/1.3005171.
Howden L, Giddings D, Power H, Aroussi A, Vloeberghs M, Garnett M, Walker D: Three-dimensional cerebrospinal fluid flow within the human ventricular system. Comput Methods Biomech Biomed Engin. 2008, 11: 123-133. 10.1080/10255840701492118.
Cifuentes M, Rodriguez S, Perez J, Grondona JM, Rodriguez EM, Fernandez-Llebrez P: Decreased cerebrospinal fluid flow through the central canal of the spinal cord of rats immunologically deprived of Reissner's fibre. Exp Brain Res. 1994, 98: 431-440. 10.1007/BF00233981.
Nakayama Y: The openings of the central canal in the filum terminale internum of some mammals. J Neurocytol. 1976, 5: 531-544. 10.1007/BF01175567.
Stoodley MA, Jones NR, Brown CJ: Evidence for rapid fluid flow from the subarachnoid space into the spinal cord central canal in the rat. Brain Res. 1996, 707: 155-164. 10.1016/0006-8993(95)01228-1.
Maillot C: The perispinal spaces. Constitution, organization and relations with the cerebrospinal fluid (CSF). J Neuroradiol. 1991, 18: 18-31.
Johnston M, Zakharov A, Koh L, Armstrong D: Subarachnoid injection of Microfil reveals connections between cerebrospinal fluid and nasal lymphatics in the non-human primate. Neuropathol Appl Neurobiol. 2005, 31: 632-640. 10.1111/j.1365-2990.2005.00679.x.
Johnston M, Zakharov A, Papaiconomou C, Salmasi G, Armstrong D: Evidence of connections between cerebrospinal fluid and nasal lymphatic vessels in humans, non-human primates and other mammalian species. Cerebrospinal Fluid Res. 2004, 1: 2-10.1186/1743-8454-1-2.
Koh L, Zakharov A, Nagra G, Armstrong D, Friendship R, Johnston M: Development of cerebrospinal fluid absorption sites in the pig and rat: connections between the subarachnoid space and lymphatic vessels in the olfactory turbinates. Anat Embryol (Berl). 2006, 211: 335-344. 10.1007/s00429-006-0085-1.
Nagra G, Koh L, Zakharov A, Armstrong D, Johnston M: Quantification of cerebrospinal fluid transport across the cribriform plate into lymphatics in rats. Am J Physiol Regul Integr Comp Physiol. 2006, 291: R1383-1389.
Walter BA, Valera VA, Takahashi S, Matsuno K, Ushiki T: Evidence of antibody production in the rat cervical lymph nodes after antigen administration into the cerebrospinal fluid. Arch Histol Cytol. 2006, 69: 37-47. 10.1679/aohc.69.37.
Johnston M, Armstrong D, Koh L: Possible role of the cavernous sinus veins in cerebrospinal fluid absorption. Cerebrospinal Fluid Res. 2007, 4: 3-10.1186/1743-8454-4-3.
Nagra G, Li J, McAllister JP, Miller J, Wagshul M, Johnston M: Impaired lymphatic cerebrospinal fluid absorption in a rat model of kaolin-induced communicating hydrocephalus. Am J Physiol Regul Integr Comp Physiol. 2008, 294: R1752-1759.
Bradbury MW, Cserr HF, Westrop RJ: Drainage of cerebral interstitial fluid into deep cervical lymph of the rabbit. Am J Physiol. 1981, 240: F329-336.
Cserr HF, Harling-Berg CJ, Knopf PM: Drainage of brain extracellular fluid into blood and deep cervical lymph and its immunological significance. Brain Pathol. 1992, 2: 269-276. 10.1111/j.1750-3639.1992.tb00703.x.
Weller RO, Engelhardt B, Phillips MJ: Lymphocyte targeting of the central nervous system: a review of afferent and efferent CNS-immune pathways. Brain Pathol. 1996, 6: 275-288. 10.1111/j.1750-3639.1996.tb00855.x.
Bering EA: Choroid plexus and arterial pulsation of cerebrospinal fluid; demonstration of the choroid plexuses as a cerebrospinal fluid pump. AMA Arch Neurol Psychiatry. 1955, 73: 165-172.
Greitz D: Cerebrospinal fluid circulation and associated intracranial dynamics. A radiologic investigation using MR imaging and radionuclide cisternography. Acta Radiol Suppl. 1993, 386: 1-23.
Greitz D, Franck A, Nordell B: On the pulsatile nature of intracranial and spinal CSF-circulation demonstrated by MR imaging. Acta Radiol. 1993, 34: 321-328.
Stivaros SM, Jackson A: Changing concepts of cerebrospinal fluid hydrodynamics: role of phase-contrast magnetic resonance imaging and implications for cerebral microvascular disease. Neurotherapeutics. 2007, 4: 511-522. 10.1016/j.nurt.2007.04.007.
Wagshul ME, Chen JJ, Egnor MR, McCormack EJ, Roche PE: Amplitude and phase of cerebrospinal fluid pulsations: experimental studies and review of the literature. J Neurosurg. 2006, 104: 810-819. 10.3171/jns.2006.104.5.810.
Egnor M, Zheng L, Rosiello A, Gutman F, Davis R: A model of pulsations in communicating hydrocephalus. Pediatr Neurosurg. 2002, 36: 281-303. 10.1159/000063533.
Greitz D: Radiological assessment of hydrocephalus: new theories and implications for therapy. Neurosurg Rev. 2004, 27: 145-165. discussion 166-147
Bhadelia RA, Bogdan AR, Kaplan RF, Wolpert SM: Cerebrospinal fluid pulsation amplitude and its quantitative relationship to cerebral blood flow pulsations: a phase-contrast MR flow imaging study. Neuroradiology. 1997, 39: 258-264. 10.1007/s002340050404.
Friese S, Hamhaber U, Erb M, Klose U: B-waves in cerebral and spinal cerebrospinal fluid pulsation measurement by magnetic resonance imaging. J Comput Assist Tomogr. 2004, 28: 255-262. 10.1097/00004728-200403000-00016.
McCormack EJ, Egnor MR, Wagshul ME: Improved cerebrospinal fluid flow measurements using phase contrast balanced steady-state free precession. Magn Reson Imaging. 2007, 25: 172-182. 10.1016/j.mri.2006.09.023.
Stoquart-ElSankari S, Baledent O, Gondry-Jouet C, Makki M, Godefroy O, Meyer ME: Aging effects on cerebral blood and cerebrospinal fluid flows. J Cereb Blood Flow Metab. 2007, 27: 1563-1572. 10.1038/sj.jcbfm.9600462.
Kelm S, Schauer R: Sialic acids in molecular and cellular interactions. Int Rev Cytol. 1997, 175: 137-240. 10.1016/S0074-7696(08)62127-0.
Nguyen T, Chin WC, O'Brien JA, Verdugo P, Berger AJ: Intracellular pathways regulating ciliary beating of rat brain ependymal cells. J Physiol. 2001, 531: 131-140. 10.1111/j.1469-7793.2001.0131j.x.
Greenstone MA, Jones RW, Dewar A, Neville BG, Cole PJ: Hydrocephalus and primary ciliary dyskinesia. Arch Dis Child. 1984, 59: 481-482. 10.1136/adc.59.5.481.
Vio K, Rodriguez S, Navarrete EH, Perez-Figares JM, Jimenez AJ, Rodriguez EM: Hydrocephalus induced by immunological blockage of the subcommissural organ-Reissner's fiber (RF) complex by maternal transfer of anti-RF antibodies. Exp Brain Res. 2000, 135: 41-52. 10.1007/s002210000474.
Sawamoto K, Wichterle H, Gonzalez-Perez O, Cholfin JA, Yamada M, Spassky N, Murcia NS, Garcia-Verdugo JM, Marin O, Rubenstein JL, Tessier-Lavigne M, Okano H, Alvarez-Buylla A: New neurons follow the flow of cerebrospinal fluid in the adult brain. Science. 2006, 311: 629-632. 10.1126/science.1119133.
Curtis MA, Kam M, Nannmark U, Anderson MF, Axell MZ, Wikkelso C, Holtas S, van Roon-Mom WM, Bjork-Eriksson T, Nordborg C, Frisen J, Dragunow M, Faull RL, Eriksson PS: Human neuroblasts migrate to the olfactory bulb via a lateral ventricular extension. Science. 2007, 315: 1243-1249. 10.1126/science.1136281.
Benraiss A, Bruel-Jungerman E, Lu G, Davidson BL, Goldman SA: Sustained ependymal transduction by aa4V-bdnf potentiates tonic neuronal addition to the adult rodent olfactory bulb. Society for Neuroscience; San Diego, CA, USA. 2007
Miyan JA, Nabiyouni M, Zendah M: Development of the brain: a vital role for cerebrospinal fluid. Can J Physiol Pharmacol. 2003, 81: 317-328. 10.1139/y03-027.
Altman J, Bayer SA: Development of the diencephalon in the rat. III. Ontogeny of the specialized ventricular linings of the hypothalamic third ventricle. J Comp Neurol. 1978, 182: 995-1015. 10.1002/cne.901820513.
Johansson PA, Dziegielewska KM, Ek CJ, Habgood MD, Liddelow SA, Potter AM, Stolp HB, Saunders NR: Blood-CSF barrier function in the rat embryo. Eur J Neurosci. 2006, 24: 65-76. 10.1111/j.1460-9568.2006.04904.x.
Bruni JE: Ependymal development, proliferation, and functions: a review. Microsc Res Tech. 1998, 41: 2-13. 10.1002/(SICI)1097-0029(19980401)41:1<2::AID-JEMT2>3.0.CO;2-Z.
Redzic ZB, Preston JE, Duncan JA, Chodobski A, Szmydynger-Chodobska J: The choroid plexus-cerebrospinal fluid system: from development to aging. Curr Top Dev Biol. 2005, 71: 1-52. 10.1016/S0070-2153(05)71001-2.
Johanson C, McMillan P, Tavares R, Spangenberger A, Duncan J, Silverberg G, Stopa E: Homeostatic capabilities of the choroid plexus epithelium in Alzheimer's disease. Cerebrospinal Fluid Res. 2004, 1: 3-10.1186/1743-8454-1-3.
Johanson CE, Duncan JA, Stopa EG, Baird A: Enhanced prospects for drug delivery and brain targeting by the choroid plexus-CSF route. Pharm Res. 2005, 22: 1011-1037. 10.1007/s11095-005-6039-0.
Nilsson C, Lindvall-Axelsson M, Owman C: Neuroendocrine regulatory mechanisms in the choroid plexus-cerebrospinal fluid system. Brain Res Brain Res Rev. 1992, 17: 109-138. 10.1016/0165-0173(92)90011-A.
MacMillan SJ, Mark MA, Duggan AW: The release of beta-endorphin and the neuropeptide-receptor mismatch in the brain. Brain Res. 1998, 794: 127-136. 10.1016/S0006-8993(98)00223-6.
Martinez-Lorenzana G, Espinosa-Lopez L, Carranza M, Aramburo C, Paz-Tres C, Rojas-Piloni G, Condes-Lara M: PVN electrical stimulation prolongs withdrawal latencies and releases oxytocin in cerebrospinal fluid, plasma, and spinal cord tissue in intact and neuropathic rats. Pain. 2008, 140: 265-273. 10.1016/j.pain.2008.08.015.
Yang J, Yang Y, Chen JM, Liu WY, Wang CH, Lin BC: Effect of oxytocin on acupuncture analgesia in the rat. Neuropeptides. 2007, 41: 285-292. 10.1016/j.npep.2007.05.004.
Amico JA, Challinor SM, Cameron JL: Pattern of oxytocin concentrations in the plasma and cerebrospinal fluid of lactating rhesus monkeys (Macaca mulatta): evidence for functionally independent oxytocinergic pathways in primates. J Clin Endocrinol Metab. 1990, 71: 1531-1535. 10.1210/jcem-71-6-1531.
Amico JA, Tenicela R, Johnston J, Robinson AG: A time-dependent peak of oxytocin exists in cerebrospinal fluid but not in plasma of humans. J Clin Endocrinol Metab. 1983, 57: 947-951. 10.1210/jcem-57-5-947.
Devarajan K, Marchant EG, Rusak B: Circadian and light regulation of oxytocin and parvalbumin protein levels in the ciliated ependymal layer of the third ventricle in the C57 mouse. Neuroscience. 2005, 134: 539-547. 10.1016/j.neuroscience.2005.04.034.
Devarajan K, Rusak B: Oxytocin levels in the plasma and cerebrospinal fluid of male rats: effects of circadian phase, light and stress. Neurosci Lett. 2004, 367: 144-147. 10.1016/j.neulet.2004.05.112.
Ludwig M, Leng G: Dendritic peptide release and peptide-dependent behaviours. Nat Rev Neurosci. 2006, 7: 126-136. 10.1038/nrn1845.
Silver R, LeSauter J, Tresco PA, Lehman MN: A diffusible coupling signal from the transplanted suprachiasmatic nucleus controlling circadian locomotor rhythms. Nature. 1996, 382: 810-813. 10.1038/382810a0.
LeSauter J, Romero P, Cascio M, Silver R: Attachment site of grafted SCN influences precision of restored circadian rhythm. J Biol Rhythms. 1997, 12: 327-338. 10.1177/074873049701200405.
Castillo-Ruiz A, Ramanathan C, Smale L, Nunez AA: Evidence that neurons of the suprachiasmatic nucleus have access to the cerebrospinal fluid in the diurnal grass rat. Society for Neuroscience; Washington, DC. 2008
Martinez Martinez P, de Weerd H: Supraependymal cell clusters in the rat hypothalamus. AnatEmbryol. 1983, 168: 371-393.
Cupedo RN, De Weerd H: Serotonergic intraventricular axons in the habenular region; Phagocytosis after induced degeneration. AnatEmbryol. 1980, 158: 213-226.
Cupedo RN, De Weerd H: Morphology of ventricular wall surfaces following neurotoxin induced degeneration of supraependymal afferents. Acta Morphol Neerl Scand. 1988, 26: 19-31.
Mathew TC: Association between supraependymal nerve fibres and the ependymal cilia of the mammalian brain. Anat Histol Embryol. 1999, 28: 193-197. 10.1046/j.1439-0264.1999.00191.x.
Singh DR, Srivastava AK, Bhartiya D, Bajpai VK, Hasan M: Surface ultra-architecture of supraependymal elements in the vicinity of the area postrema of rhesus monkeys. Brain Res Bull. 1989, 23: 175-181. 10.1016/0361-9230(89)90144-5.
Cloft HJ, Mitchell JA: Immunocytochemical detection of oxytocin in the supraependymal neuronal complex of the golden hamster. Brain Res. 1994, 639: 233-239. 10.1016/0006-8993(94)91735-3.
Cloft HJ, Mitchell JA: Serotonergic innervation of the supraependymal neuronal complex of the golden hamster. Brain Res. 1997, 761: 210-216. 10.1016/S0006-8993(97)00324-7.
Chan-Palay V: Serotonin axons in the supra- and subependymal plexuses and in the leptomeninges; their roles in local alterations of cerebrospinal fluid and vasomotor activity. Brain Res. 1976, 102: 103-130. 10.1016/0006-8993(76)90578-3.
Brusco A, Lopez-Costa JJ, Tagliaferro P, Pecci Saavedra J: Serotonergic ependymal fibres in rat and monkey: light and electron microscopic immunocytochemical study. Biocell. 1998, 22: 115-122.
Parent A: Anatomical organization of monoamine - and acetylcholinesterase - containing neuronal systems in the vertebrate hypothalamus. Anatomy of the hypothalamus. Edited by: Morgane PJ, Panksepp J. 1979, New York: Marcel dekker, 1: 511-554. Morgane PJ, Panksepp, J. (Series Editor): Handbook of the hypothalamus
Martinez PM, de Weerd H: The fine structure of the ependymal surface of the recessus infundibularis in the rat. Anat Embryol (Berl). 1977, 151: 241-265. 10.1007/BF00318929.
Buma P, Veening J, Nieuwenhuys R: Ultrastructural Characterization of Adrenocorticotrope Hormone (ACTH) Immunoreactive Fibres in the Mesencephalic Central Grey Substance of the Rat. Eur J Neurosci. 1989, 1: 659-672. 10.1111/j.1460-9568.1989.tb00372.x.
Skinner DC, Malpaux B: High melatonin concentrations in third ventricular cerebrospinal fluid are not due to Galen vein blood recirculating through the choroid plexus. Endocrinology. 1999, 140: 4399-4405. 10.1210/en.140.10.4399.
Malpaux B, Tricoire H, Mailliet F, Daveau A, Migaud M, Skinner DC, Pelletier J, Chemineau P: Melatonin and seasonal reproduction: understanding the neuroendocrine mechanisms using the sheep as a model. Reprod Suppl. 2002, 59: 167-179.
Tricoire H, Malpaux B, Moller M: Cellular lining of the sheep pineal recess studied by light-, transmission-, and scanning electron microscopy: morphologic indications for a direct secretion of melatonin from the pineal gland to the cerebrospinal fluid. J Comp Neurol. 2003, 456: 39-47. 10.1002/cne.10477.
Thiery JC, Lomet D, Schumacher M, Liere P, Tricoire H, Locatelli A, Delagrange P, Malpaux B: Concentrations of estradiol in ewe cerebrospinal fluid are modulated by photoperiod through pineal-dependent mechanisms. J Pineal Res. 2006, 41: 306-312. 10.1111/j.1600-079X.2006.00370.x.
Malpaux B, Daveau A, Maurice-Mandon F, Duarte G, Chemineau P: Evidence that melatonin acts in the premammillary hypothalamic area to control reproduction in the ewe: presence of binding sites and stimulation of luteinizing hormone secretion by in situ microimplant delivery. Endocrinology. 1998, 139: 1508-1516. 10.1210/en.139.4.1508.
Skinner DC, Caraty A, Malpaux B, Evans NP: Simultaneous measurement of gonadotropin-releasing hormone in the third ventricular cerebrospinal fluid and hypophyseal portal blood of the ewe. Endocrinology. 1997, 138: 4699-4704. 10.1210/en.138.11.4699.
Skinner DC, Malpaux B, Delaleu B, Caraty A: Luteinizing hormone (LH)-releasing hormone in third ventricular cerebrospinal fluid of the ewe: correlation with LH pulses and the LH surge. Endocrinology. 1995, 136: 3230-3237. 10.1210/en.136.8.3230.
Skinner DC, Caraty A, Evans NP: Does gonadotropin-releasing hormone in the cerebrospinal fluid modulate luteinizing hormone release?. Neuroendocrinology. 1998, 67: 37-44. 10.1159/000054296.
Caraty A, Skinner DC: Gonadotropin-releasing hormone in third ventricular cerebrospinal fluid: endogenous distribution and exogenous uptake. Endocrinology. 2008, 149: 5227-5234. 10.1210/en.2007-1636.
Riskind P, Moss RL: Effects of lesions of putative LHRH-containing pathways and midbrain nuclei on lordotic behavior and luteinizing hormone release in ovariectomized rats. Brain Res Bull. 1983, 11: 493-500. 10.1016/0361-9230(83)90120-X.
Riskind P, Moss RL: Midbrain LHRH infusions enhance lordotic behavior in ovariectomized estrogen-primed rats independently of a hypothalamic responsiveness to LHRH. Brain Res Bull. 1983, 11: 481-485. 10.1016/0361-9230(83)90118-1.
Schwartz-Giblin S, McCarthy MM: A sexual column in the PAG?. Trends Neurosci. 1995, 18: 129-10.1016/0166-2236(95)93889-6.
Ogawa S, Kow LM, Pfaff DW: Effects of lordosis-relevant neuropeptides on midbrain periaqueductal gray neuronal activity in vitro. Peptides. 1992, 13: 965-975. 10.1016/0196-9781(92)90058-B.
Caraty A, Delaleu B, Chesneau D, Fabre-Nys C: Sequential role of e2 and GnRH for the expression of estrous behavior in ewes. Endocrinology. 2002, 143: 139-145. 10.1210/en.143.1.139.
Ross JF, Switzer RC, Poston MR, Lawhorn GT: Distribution of bismuth in the brain after intraperitoneal dosing of bismuth subnitrate in mice: implications for routes of entry of xenobiotic metals into the brain. Brain Res. 1996, 725: 137-154.
Ritter RC, Slusser PG, Stone S: Glucoreceptors controlling feeding and blood glucose: location in the hindbrain. Science. 1981, 213: 451-452. 10.1126/science.6264602.
Adam CL, Findlay PA, Miller DW: Blood-brain leptin transport and appetite and reproductive neuroendocrine responses to intracerebroventricular leptin injection in sheep: influence of photoperiod. Endocrinology. 2006, 147: 4589-4598. 10.1210/en.2006-0576.
Faulconbridge LF, Grill HJ, Kaplan JM: Distinct forebrain and caudal brainstem contributions to the neuropeptide Y mediation of ghrelin hyperphagia. Diabetes. 2005, 54: 1985-1993. 10.2337/diabetes.54.7.1985.
Grill HJ, Carmody JS, Amanda Sadacca L, Williams DL, Kaplan JM: Attenuation of lipopolysaccharide anorexia by antagonism of caudal brain stem but not forebrain GLP-1-R. Am J Physiol Regul Integr Comp Physiol. 2004, 287: R1190-1193.
Aja S, Schwartz GJ, Kuhar MJ, Moran TH: Intracerebroventricular CART peptide reduces rat ingestive behavior and alters licking microstructure. Am J Physiol Regul Integr Comp Physiol. 2001, 280: R1613-1619.
Flynn FW: Fourth ventricle bombesin injection suppresses ingestive behaviors in rats. Am J Physiol. 1989, 256: R590-596.
Blevins JE, Schwartz MW, Baskin DG: Evidence that paraventricular nucleus oxytocin neurons link hypothalamic leptin action to caudal brain stem nuclei controlling meal size. Am J Physiol Regul Integr Comp Physiol. 2004, 287: R87-96.
Plata-Salaman CR, Oomura Y: Calcitonin as a feeding suppressant: localization of central action to the cerebral III ventricle. Physiol Behav. 1987, 40: 501-513. 10.1016/0031-9384(87)90037-0.
Fenstermacher J, Kaye T: Drug "diffusion" within the brain. Ann N Y Acad Sci. 1988, 531: 29-39. 10.1111/j.1749-6632.1988.tb31809.x.
Proescholdt MG, Hutto B, Brady LS, Herkenham M: Studies of cerebrospinal fluid flow and penetration into brain following lateral ventricle and cisterna magna injections of the tracer [14C]inulin in rat. Neuroscience. 2000, 95: 577-592. 10.1016/S0306-4522(99)00417-0.
De Rotte AA, Bouman HJ, van Wimersma Greidanus TB: Relationships between alpha-MSH levels in blood and in cerebrospinal fluid. Brain Res Bull. 1980, 5: 375-381. 10.1016/0361-9230(80)90060-X.
Bach FW, Yaksh TL: Release into ventriculo-cisternal perfusate of beta-endorphin- and Met-enkephalin-immunoreactivity: effects of electrical stimulation in the arcuate nucleus and periaqueductal gray of the rat. Brain Res. 1995, 690: 167-176. 10.1016/0006-8993(95)00600-U.
Bittencourt JC, Sawchenko PE: Do centrally administered neuropeptides access cognate receptors?: an analysis in the central corticotropin-releasing factor system. J Neurosci. 2000, 20: 1142-1156.
Konsman JP, Tridon V, Dantzer R: Diffusion and action of intracerebroventricularly injected interleukin-1 in the CNS. Neuroscience. 2000, 101: 957-967. 10.1016/S0306-4522(00)00403-6.
Sabatier N, Rowe I, Leng G: Central release of oxytocin and the ventromedial hypothalamus. Biochem Soc Trans. 2007, 35: 1247-1251. 10.1042/BST0351247.
Nagaraja TN, Patel P, Gorski M, Gorevic PD, Patlak CS, Fenstermacher JD: In normal rat, intraventricularly administered insulin-like growth factor-1 is rapidly cleared from CSF with limited distribution into brain. Cerebrospinal Fluid Res. 2005, 2: 5-10.1186/1743-8454-2-5.
Fenstermacher JD, Ghersi-Egea JF, Finnegan W, Chen JL: The rapid flow of cerebrospinal fluid from ventricles to cisterns via subarachnoid velae in the normal rat. Acta Neurochir Suppl. 1997, 70: 285-287.
Nicholson C, Sykova E: Extracellular space structure revealed by diffusion analysis. Trends Neurosci. 1998, 21: 207-215. 10.1016/S0166-2236(98)01261-2.
Sykova E: Extrasynaptic volume transmission and diffusion parameters of the extracellular space. Neuroscience. 2004, 129: 861-876. 10.1016/j.neuroscience.2004.06.077.
Sykova E, Vargova L: Extrasynaptic transmission and the diffusion parameters of the extracellular space. Neurochem Int. 2008, 52: 5-13. 10.1016/j.neuint.2007.04.007.
Geer CP, Grossman SA: Interstitial fluid flow along white matter tracts: a potentially important mechanism for the dissemination of primary brain tumors. J Neurooncol. 1997, 32: 193-201. 10.1023/A:1005761031077.
Bruni JE, Del Bigio MR, Clattenburg RE: Ependyma: normal and pathological. A review of the literature. Brain Res. 1985, 356: 1-19.
McKinley MJ, Clarke IJ, Oldfield BJ: Circumventricular organs. The human nervous system. Edited by: Paxinos G, Mai JK. 2004, New York: Academic Press, 562-591. full_text. 2
Ericsson A, Kovacs KJ, Sawchenko PE: A functional anatomical analysis of central pathways subserving the effects of interleukin-1 on stress-related neuroendocrine neurons. J Neurosci. 1994, 14: 897-913.
Ferguson AV, Bains JS: Actions of angiotensin in the subfornical organ and area postrema: implications for long term control of autonomic output. Clin Exp Pharmacol Physiol. 1997, 24: 96-101. 10.1111/j.1440-1681.1997.tb01790.x.
Herman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, Ostrander MM, Choi DC, Cullinan WE: Central mechanisms of stress integration: hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front Neuroendocrinol. 2003, 24: 151-180. 10.1016/j.yfrne.2003.07.001.
Korf HW: Neuronal organization of the avian paraventricular nucleus: intrinsic, afferent, and efferent connections. J Exp Zool. 1984, 232: 387-395. 10.1002/jez.1402320303.
Larsen PJ, Mikkelsen JD: Functional identification of central afferent projections conveying information of acute "stress" to the hypothalamic paraventricular nucleus. J Neurosci. 1995, 15: 2609-2627.
Larsen PJ, Moller M, Mikkelsen JD: Efferent projections from the periventricular and medial parvicellular subnuclei of the hypothalamic paraventricular nucleus to circumventricular organs of the rat: a Phaseolus vulgaris-leucoagglutinin (PHA-L) tracing study. J Comp Neurol. 1991, 306: 462-479. 10.1002/cne.903060310.
Lind RW, Swanson LW, Ganten D: Organization of angiotensin II immunoreactive cells and fibers in the rat central nervous system. An immunohistochemical study. Neuroendocrinology. 1985, 40: 2-24. 10.1159/000124046.
Silverman AJ, Hoffman DL, Zimmerman EA: The descending afferent connections of the paraventricular nucleus of the hypothalamus (PVN). Brain Res Bull. 1981, 6: 47-61. 10.1016/S0361-9230(81)80068-8.
Bui JD, Nammari DR, Buckley DL, Inglis BA, Silver XS, Mareci TH, Phillips MI: In vivo dynamics and distribution of intracerebroventricularly administered gadodiamide, visualized by magnetic resonance imaging. Neuroscience. 1999, 90: 1115-1122. 10.1016/S0306-4522(98)00519-3.
Ferguson IA, Schweitzer JB, Bartlett PF, Johnson EM: Receptor-mediated retrograde transport in CNS neurons after intraventricular administration of NGF and growth factors. J Comp Neurol. 1991, 313: 680-692. 10.1002/cne.903130411.
Hoistad M, Samskog J, Jacobsen KX, Olsson A, Hansson HA, Brodin E, Fuxe K: Detection of beta-endorphin in the cerebrospinal fluid after intrastriatal microinjection into the rat brain. Brain Res. 2005, 1041: 167-180. 10.1016/j.brainres.2005.02.014.
Rennels ML, Gregory TF, Blaumanis OR, Fujimoto K, Grady PA: Evidence for a 'paravascular' fluid circulation in the mammalian central nervous system, provided by the rapid distribution of tracer protein throughout the brain from the subarachnoid space. Brain Res. 1985, 326: 47-63. 10.1016/0006-8993(85)91383-6.
Hadaczek P, Yamashita Y, Mirek H, Tamas L, Bohn MC, Noble C, Park JW, Bankiewicz K: The "perivascular pump" driven by arterial pulsation is a powerful mechanism for the distribution of therapeutic molecules within the brain. Mol Ther. 2006, 14: 69-78. 10.1016/j.ymthe.2006.02.018.
Rennels ML, Blaumanis OR, Grady PA: Rapid solute transport throughout the brain via paravascular fluid pathways. Adv Neurol. 1990, 52: 431-439.
Cifuentes M, Fernandez LP, Perez J, Perez-Figares JM, Rodriguez EM: Distribution of intraventricularly injected horseradish peroxidase in cerebrospinal fluid compartments of the rat spinal cord. Cell Tissue Res. 1992, 270: 485-494. 10.1007/BF00645050.
Stoodley MA, Brown SA, Brown CJ, Jones NR: Arterial pulsation-dependent perivascular cerebrospinal fluid flow into the central canal in the sheep spinal cord. J Neurosurg. 1997, 86: 686-693. 10.3171/jns.1997.86.4.0686.
Rodriguez EM, Blazquez JL, Pastor FE, Pelaez B, Pena P, Peruzzo B, Amat P: Hypothalamic tanycytes: a key component of brain-endocrine interaction. Int Rev Cytol. 2005, 247: 89-164. 10.1016/S0074-7696(05)47003-5.
Zhang LC, Zeng YM, Ting J, Cao JP, Wang MS: The distributions and signaling directions of the cerebrospinal fluid contacting neurons in the parenchyma of a rat brain. Brain Res. 2003, 989: 1-8. 10.1016/S0006-8993(03)03123-8.
Cheunsuang O, Stewart AL, Morris R: Differential uptake of molecules from the circulation and CSF reveals regional and cellular specialisation in CNS detection of homeostatic signals. Cell Tissue Res. 2006, 325: 397-402. 10.1007/s00441-006-0162-z.
Peruzzo B, Pastor FE, Blazquez JL, Amat P, Rodriguez EM: Polarized endocytosis and transcytosis in the hypothalamic tanycytes of the rat. Cell Tissue Res. 2004, 317: 147-164. 10.1007/s00441-004-0899-1.
Nonaka N, Farr SA, Kageyama H, Shioda S, Banks WA: Delivery of galanin-like peptide to the brain: targeting with intranasal delivery and cyclodextrins. J Pharmacol Exp Ther. 2008, 325: 513-519. 10.1124/jpet.107.132381.
Torterolo P, Lagos P, Sampogna S, Chase MH: Melanin-concentrating hormone (MCH) immunoreactivity in non-neuronal cells within the raphe nuclei and subventricular region of the brainstem of the cat. Brain Res. 2008, 1210: 163-178. 10.1016/j.brainres.2008.02.104.
Lu X, Geng X, Zhang L, Zeng Y: The methodology for labeling the distal cerebrospinal fluid-contacting neurons in rats. J Neurosci Methods. 2008, 168: 98-103. 10.1016/j.jneumeth.2007.09.033.
Feng C, Baryshnikova LM, von Bartheld CS: A novel systemic route for specific accumulation of Nerve Growth Factor in the Central Nervous System. Society for Neuroscience; San Diego, CA, USA. 2007
Abbott NJ: Evidence for bulk flow of brain interstitial fluid: significance for physiology and pathology. Neurochem Int. 2004, 45: 545-552. 10.1016/j.neuint.2003.11.006.
Condes-Lara M, Martinez-Lorenzana G, Rodriguez-Jimenez J, Rojas-Piloni G: Paraventricular hypothalamic nucleus stimulation modulates nociceptive responses in dorsal horn wide dynamic range neurons. Neurosci Lett. 2008, 444: 199-202. 10.1016/j.neulet.2008.08.027.
Yang J, Yang Y, Chen JM, Liu WY, Wang CH, Lin BC: Central oxytocin enhances antinociception in the rat. Peptides. 2007, 28: 1113-1119. 10.1016/j.peptides.2007.03.003.
Yu GZ, Kaba H, Okutani F, Takahashi S, Higuchi T, Seto K: The action of oxytocin originating in the hypothalamic paraventricular nucleus on mitral and granule cells in the rat main olfactory bulb. Neuroscience. 1996, 72: 1073-1082. 10.1016/0306-4522(95)00599-4.
Harrington MG, Fonteh AN, Oborina E, Liao P, Cowan RP, McComb G, Chavez JN, Rush J, Biringer RG, Huhmer AF: The morphology and biochemistry of nanostructures provide evidence for synthesis and signaling functions in human cerebrospinal fluid. Cerebrospinal Fluid Res. 2009, 6: 10-10.1186/1743-8454-6-10.
Waldherr M, Neumann ID: Centrally released oxytocin mediates mating-induced anxiolysis in male rats. Proc Natl Acad Sci USA. 2007, 104: 16681-16684. 10.1073/pnas.0705860104.
Veenema AH, Neumann ID: Central vasopressin and oxytocin release: regulation of complex social behaviours. Prog Brain Res. 2008, 170: 261-276. full_text.
Acknowledgements
Authors wish to thank Trynke de Jong for reading part of the manuscript and for several useful suggestions for improvement. In addition, they wish to thank Bruce Jenks and Tracy Brandmeyer for reading and improving the style of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
JV carried out most of the writing and composition of the paper and the preceding research. HB made substantial contributions to the conception and design of the paper, to the interpretation of the reviewed findings and its critical revision. All authors have read and approved the final version of the manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Veening, J.G., Barendregt, H.P. The regulation of brain states by neuroactive substances distributed via the cerebrospinal fluid; a review. Fluids Barriers CNS 7, 1 (2010). https://doi.org/10.1186/1743-8454-7-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1743-8454-7-1