
Abstract
Epidemiologic studies indicate that bacterial vaginosis (BV), a common alteration of lower genital tract flora in women, is associated with increased susceptibility to HIV infection. Other recent studies show that HIV is detected more frequently and at higher levels in the lower genital tract of HIV-seropositive women with BV. In vitro studies show that genital tract secretions from women with BV or flora associated with BV induce HIV expression in infected cells. The increased HIV expression appears to be due at least in part to activation through Toll-like receptors (TLR), specifically TLR2. Further research is needed to elucidate how BV contributes to HIV acquisition and transmission.
Similar content being viewed by others

Review
Bacterial vaginosis
Bacteria colonize the lower genital tract of most women and the predominant species of bacteria in healthy women is lactobacilli. Commonly found vaginal lactobacillus species include Lactobacillus crispatus, L. gasseri, L. jensenii and L. iners [1, 2]. Bacterial vaginosis (BV) is characterized by an alteration of genital tract flora such that the predominant bacteria are no longer lactobacilli, but instead consist of polymicrobial communities of multiple genera of gram positive and gram negative organisms [3]. Gardnerella vaginalis, Prevotella sp.,Bacteroides sp., Peptostreptococcus sp., Mycoplasma hominis and Mobiluncus sp. as well as other recently described bacteria are commonly found in BV [2, 3]. Lactobacilli, usually L. iners, are also frequently present in BV, but make up a relatively small proportion of the total flora [2, 4]. BV has been noted to be the most prevalent vaginal disorder in adult women worldwide with the frequency depending on the group that is studied [5]. BV is found in 24% to 37% of women attending STD clinics but seen at lower rates in women that are not sexually active.
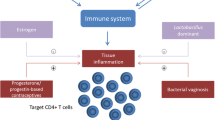
BV is associated with an increased risk of infections by HIV and some other organisms as discussed below, as well as with increased risk of preterm birth, which is a leading cause of infant death in the United States [6–8]. Treatment of BV can reduce preterm birth in high risk cases [7, 9]. BV is also associated with miscarriage and pelvic inflammatory disease [10–12].
Diagnosis of BV is commonly made by examination of four criteria: vaginal fluid pH (BV results in a pH >4.5); presence of clue cells (bacteria-coated epithelial cells); a homogenous discharge; and production of an amine odor when KOH is added to vaginal fluid [13]. Gram stains of vaginal fluid can also aid in diagnosis of BV [14].
Oral or intravaginal antibiotic treatment with metronidazole or clindamycin cures BV in most women, but BV can resolve spontaneously in nearly a third of subjects [15–18]. However, BV recurs in a significant fraction of treated women.
In vivo studies of the effects of BV on HIV susceptibility and expression
Several cross-sectional studies performed in Thailand [19], Uganda [20], and Malawi [21, 22] showed that women with BV had an increased incidence of HIV infection. While suggestive, these studies do not prove a cause and effect relationship between BV and HIV infection. However, a prospective study in Kenya [23] showed that the presence of BV and the absence of lactobacilli or absence of hydrogen peroxide-producing lactobacilli upon examination were all significantly associated with acquisition of HIV infection at follow-up.
There is also evidence, some of it through cross-sectional studies, that the presence of BV increases the risk of infection with several other sexually transmitted infections (STI), including herpes simplex virus type 2 (HSV-2), gonorrhea, Trichomonas vaginalis and Chlamydia trachomatis [23–25]. All of these STI have been suggested to increase susceptibility of women to sexual transmission of HIV [26], and so BV may both directly increase HIV susceptibility and indirectly increase it by increasing the number of women with these other STI.
While the above studies suggest that BV can influence susceptibility of women to HIV infection, other recent studies suggest that BV increases expression of HIV in the lower genital tract of women that are already infected with HIV. Thus, the levels of HIV, as assessed by HIV RNA, and the detection frequency of HIV in the genital tract are significantly higher in the genital tract of women with BV when compared to women without BV [27, 28]. HIV levels were inversely correlated with levels of lactobacilli but positively correlated with Mycoplasma hominis [27]. An additional study showed that women with lower levels of vaginal lactobacilli had higher genital tract HIV [29].
A number of mechanisms have been suggested that could account for the increase in susceptibility to HIV and/or increased expression of HIV in the genital tract (Table 1) [26, 30]. These include decreased levels of hydrogen peroxide-producing lactobacilli, production by BV flora of enzymes (e.g. mucinases) that degrade protective mechanisms such as mucous, or production by BV flora of stimulatory substances that increase influx of target cells, HIV expression or infection of cells (see below). In fact, BV is associated with increased levels of pro-inflammatory cytokines such as IL-1β and IL-8 [Reviewed in [31]]. IL-1β can induce the production of other pro-inflammatory cytokines, and IL-8 is known to recruit immune cells, thus possibly increasing the number of cellular targets for HIV infection [32].
In vitro studies of the relationship between BV and HIV
A number of in vitro studies show that genital tract fluids from women with BV are highly stimulatory for immune cells and can up-regulate expression of HIV. Thus, incubation of the chronically-HIV-infected monocytic cell line U1 with genital fluid from women with BV substantially increased HIV expression [33–36]. In contrast, genital fluid collected from women without BV did not induce HIV expression. HIV expression was also induced in T cell lines and in peripheral blood mononuclear cells by genital fluids from women with BV [35, 37]. The substances in genital fluids that stimulated HIV expression in cells were found to function through activation of NF-kB [37].
Bacteria from BV have also been tested for their ability to stimulate HIV expression in cells. Gardnerella vaginalis, the bacterium most frequently isolated in BV, significantly induced HIV expression in U1 cells [38, 39]. Lysozyme treatment reduced U1 activation suggesting a cell wall component of G. vaginalis was involved in stimulation of the U1 cells. Anaerobes Peptostreptococcus asaccharolyticus and Prevotella bivia also stimulated HIV expression [40] as did non-anaerobic bacteria Mycoplasma hominis and Streptococcus [39]. In contrast, other bacteria found in genital samples including Bacteroides ureolyticus, Peptostreptococcus anaerobius, and Lactobacillus acidophilus did not stimulate HIV expression [40].
Taken together, many of the above studies suggested that the HIV-stimulatory activity in genital fluids acted through Toll-like receptors (TLR). For example, genital fluids stimulated HIV expression through the NF-kB pathway [41], and stimulation of cells through TLR is well documented to activate NF-kB [42]. Also, many of the ligands for TLR are bacterial products [42] and BV mucosal fluids would be expected to contain such products. A recent study using the 293 cell line modified to express either TLR2, TLR4 or control cells expressing no functional TLR, directly determined whether mucosal fluids from women with BV stimulated cells through TLR [41]. The results showed that genital fluids from women with BV stimulated cells predominantly through TLR2, while surprisingly there was relatively little stimulation through TLR4. In contrast, fluids from women without BV stimulated cells relatively little through either TLR2 or TLR4. Further, the TLR2-positive cells supported higher levels of expression of the HIV promoter when exposed to genital secretions from women with BV, suggesting that HIV-infected cells in the genital tract might express higher levels of HIV during episodes of BV. Other studies showed that genital tract fluid from women with BV can stimulate lymphocytes and other cells to express higher levels of TLR4 and TNF-α [43].
Dendritic cells (DC), cells important for antigen processing and presentation to the immune system, are found in the lower genital tract and are known to express both TLR2 and TLR4. DC are suggested to be one of the first cells that take up HIV during sexual transmission [44, 45]. DC are also potent antigen presenting cells whose function would be important for vaccination against a number of mucosal pathogens, including HIV. We have investigated the hypothesis that genital tract secretions from women with BV might substantially affect either DC antigen presenting function or DC uptake and infection by HIV. We observed that secretions from women with BV potently stimulate secretion of IL-12 by monocyte-derived dendritic (MDDC) (Figure 1) [46]. Genital fluid from women with BV also increased MDDC secretion of IL-23 and p40 and upregulated cell surface HLA-DR, CD40 and CD83. Further, BV fluids decreased MDDC endocytic ability (a marker of stimulation and maturation of DC) and increased proliferation of T cells in an allogeneic MLR with MDDC as the antigen presenting cells [46]. Genital fluids from women without BV had much lower or no stimulatory activity for MDDC. These studies suggest that BV may substantially affect local DC antigen presenting function in women.

Bacterial Vaginosis induces IL-12p70 production by Dendritic Cells. Monocyte-derived dendritic cells (MDDC) were produced from monocytes isolated from the blood of normal donors using standard methods [46]. MDDC were incubated for 48 hours with either culture medium alone (Medium), lipopolysaccharide at 1 μg/ml (LPS), or genital tract secretions collected by cervicalvaginal lavage from women with BV (BV CVL) or normal flora (Normal CVL). The BV CVL and Normal CVL were pools of equal amounts of CVL from 15 and 14 women respectively. Status of CVL donors was determined by gram stain. Supernatants were harvested and analyzed for IL-12p70 by ELISA.
Since the above studies showed that BV genital secretions potently stimulate DC, we hypothesized that this stimulation might increase infection of DC or enhance the ability of DC to transfer HIV to T cells. However, our studies to date do not show BV enhancing HIV infection of DC (Fig. 2) or transfer of HIV by DC to T cells (Fig. 3). In fact, BV genital secretions appear to suppress HIV transfer to T cells (Fig. 3). While our studies currently do not support a role for direct effects of BV genital secretions on DC in enhancing HIV transmission, these findings do not rule out the possibility that BV promotes HIV transmission by altering DC function or trafficking in vivo.

Effect of Bacterial Vaginosis on HIV infection of MDDC. MDDC were produced and treated with either Medium alone, BV CVL, Normal CVL or LPS for 48 hr as described in the Figure 1 legend. Treated MDDC were incubated with HIV-1Bal for 24 hr. DNA was then isolated from the MDDC and analyzed for HIV DNA copies by real time PCR. Bars represent mean + standard error.

Effect of Bacterial Vaginosis on HIV transfer from MDDC to T cells. MDDC were produced and treated as described in the Figure 2 legend and then exposed to HIV-Bal for 2 hours. Free virus was removed by washing and MDDC were incubated with PHA stimulated PBMC. Five days later supernatants were harvested analyzed for p24 production by ELISA. Bars represent mean + standard error.
Conclusion
While it has become evident that BV has effects on HIV transmission, HIV genital tract levels and HIV expression in vitro, further work is needed to identify the mechanisms responsible for these effects. For example, questions remain regarding the direct contribution of bacterial flora versus indirect mechanisms through immune cells, immune mediators such as cytokines or other mediators. New in vitro experimental systems or animal models are needed to help elucidate these mechanisms and are likely to lead to increased understanding of ways to prevent the spread of the HIV epidemic.
References
Antonio MA, Hawes SE, Hillier SL: The identification of vaginal Lactobacillus species and the demographic and microbiologic characteristics of women colonized by these species. J Infect Dis. 1999, 180: 1950-1956. 10.1086/315109
Fredricks DN, Fiedler TL, Marrazzo JM: Molecular identification of bacteria associated with bacterial vaginosis. N Engl J Med. 2005, 353: 1899-1911. 10.1056/NEJMoa043802
Hill GB: The microbiology of bacterial vaginosis. Am J Obstet Gynecol. 1993, 169: 450-454.
Eschenbach DA, Davick PR, Williams BL, Klebanoff SJ, Young-Smith K, Critchlow CM, Holmes KK: Prevalence of hydrogen peroxide-producing Lactobacillus species in normal women and women with bacterial vaginosis. J Clin Microbiol. 1989, 27: 251-256.
Sobel JD: What's new in bacterial vaginosis and trichomoniasis?. Infect Dis Clin North Am. 2005, 19: 387-406. 10.1016/j.idc.2005.03.001
Hillier SL, Nugent RP, Eschenbach DA, Krohn MA, Gibbs RS, Martin DH, Cotch MF, Edelman R, Pastorek JG, Rao AV: Association between bacterial vaginosis and preterm delivery of a low-birth-weight infant. The Vaginal Infections and Prematurity Study Group [see comments]. N Engl J Med. 1995, 333: 1737-1742. 10.1056/NEJM199512283332604
McGregor JA, French JI, Parker R, Draper D, Patterson E, Jones W, Thorsgard K, McFee J: Prevention of premature birth by screening and treatment for common genital tract infections: results of a prospective controlled evaluation. Am J Obstet Gynecol. 1995, 173: 157-167. 10.1016/0002-9378(95)90184-1
Leitich H, Bodner-Adler B, Brunbauer M, Kaider A, Egarter C, Husslein P: Bacterial vaginosis as a risk factor for preterm delivery: a meta-analysis. Am J Obstet Gynecol. 2003, 189: 139-147. 10.1067/mob.2003.339
Hauth JC, Goldenberg RL, Andrews WW, DuBard MB, Copper RL: Reduced incidence of preterm delivery with metronidazole and erythromycin in women with bacterial vaginosis [see comments]. N Engl J Med. 1995, 333: 1732-1736. 10.1056/NEJM199512283332603
Hillier SL, Kiviat NB, Hawes SE, Hasselquist MB, Hanssen PW, Eschenbach DA, Holmes KK: Role of bacterial vaginosis-associated microorganisms in endometritis. Am J Obstet Gynecol. 1996, 175: 435-441. 10.1016/S0002-9378(96)70158-8
Ralph SG, Rutherford AJ, Wilson JD: Influence of bacterial vaginosis on conception and miscarriage in the first trimester: cohort study. Bmj. 1999, 319: 220-223.
Haggerty CL, Hillier SL, Bass DC, Ness RB: Bacterial vaginosis and anaerobic bacteria are associated with endometritis. Clin Infect Dis. 2004, 39: 990-995. 10.1086/423963
Amsel R, Totten PA, Spiegel CA, Chen KC, Eschenbach D, Holmes KK: Nonspecific vaginitis. Diagnostic criteria and microbial and epidemiologic associations. Am J Med. 1983, 74: 14-22. 10.1016/0002-9343(83)91112-9
Nugent RP, Krohn MA, Hillier SL: Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. J Clin Microbiol. 1991, 29: 297-301.
Sobel JD: Vaginitis. N Engl J Med. 1997, 337: 1896-1903. 10.1056/NEJM199712253372607
Owen MK, Clenney TL: Management of vaginitis. Am Fam Physician. 2004, 70: 2125-2132.
Hay PE: Recurrent bacterial vaginosis. Dermatol Clin. 1998, 16: 769-773. xii–xiii, 10.1016/S0733-8635(05)70044-9
Carey JC, Klebanoff MA, Hauth JC, Hillier SL, Thom EA, Ernest JM, Heine RP, Nugent RP, Fischer ML, Leveno KJ: Metronidazole to prevent preterm delivery in pregnant women with asymptomatic bacterial vaginosis. National Institute of Child Health and Human Development Network of Maternal-Fetal Medicine Units [see comments]. N Engl J Med. 2000, 342: 534-540. 10.1056/NEJM200002243420802
Cohen CR, Duerr A, Pruithithada N, Rugpao S, Hillier S, Garcia P, Nelson K: Bacterial vaginosis and HIV seroprevalence among female commercial sex workers in Chiang Mai, Thailand. Aids. 1995, 9: 1093-1097. 10.1097/00002030-199509000-00017
Sewankambo N, Gray RH, Wawer MJ, Paxton L, McNaim D, Wabwire-Mangen F, Serwadda D, Li C, Kiwanuka N, Hillier SL: HIV-1 infection associated with abnormal vaginal flora morphology and bacterial vaginosis [see comments] [published erratum appears in Lancet 1997 Oct 4;350(9083):1036]. Lancet. 1997, 350: 546-550. 10.1016/S0140-6736(97)01063-5
Taha TE, Hoover DR, Dallabetta GA, Kumwenda NI, Mtimavalye LA, Yang LP, Liomba GN, Broadhead RL, Chiphangwi JD, Miotti PG: Bacterial vaginosis and disturbances of vaginal flora: association with increased acquisition of HIV. Aids. 1998, 12: 1699-1706. 10.1097/00002030-199813000-00019
Taha TE, Gray RH, Kumwenda NI, Hoover DR, Mtimavalye LA, Liomba GN, Chiphangwi JD, Dallabetta GA, Miotti PG: HIV infection and disturbances of vaginal flora during pregnancy. J Acquir Immune Defic Syndr Hum Retrovirol. 1999, 20: 52-59.
Martin HL, Richardson BA, Nyange PM, Lavreys L, Hillier SL, Chohan B, Mandaliya K, Ndinya-Achola JO, Bwayo J, Kreiss J: Vaginal lactobacilli, microbial flora, and risk of human immunodeficiency virus type 1 and sexually transmitted disease acquisition. J Infect Dis. 1999, 180: 1863-1868. 10.1086/315127
Cherpes TL, Meyn LA, Krohn MA, Lurie JG, Hillier SL: Association between acquisition of herpes simplex virus type 2 in women and bacterial vaginosis. Clin Infect Dis. 2003, 37: 319-325. 10.1086/375819
Wiesenfeld HC, Hillier SL, Krohn MA, Landers DV, Sweet RL: Bacterial vaginosis is a strong predictor of Neisseria gonorrhoeae and Chlamydia trachomatis infection. Clin Infect Dis. 2003, 36: 663-668. 10.1086/367658
Sobel JD: Gynecologic infections in human immunodeficiency virus-infected women. Clin Infect Dis. 2000, 31: 1225-1233. 10.1086/317436
Sha BE, Zariffard MR, Wang QJ, Chen HY, Bremer J, Cohen MH, Spear GT: Female genital-tract HIV load correlates inversely with Lactobacillus species but positively with bacterial vaginosis and Mycoplasma hominis. J Infect Dis. 2005, 191: 25-32. 10.1086/426394
Cu-Uvin S, Hogan JW, Caliendo AM, Harwell J, Mayer KH, Carpenter CC: Association between bacterial vaginosis and expression of human immunodeficiency virus type 1 RNA in the female genital tract. Clin Infect Dis. 2001, 33: 894-896. 10.1086/322613
Coleman JS, Hitti J, Bukusi EA, Mwachari C, Muliro A, Nguti R, Gausman R, Jensen S, Patton D, Lockhart D: Infectious correlates of HIV-1 shedding in the female upper and lower genital tracts. Aids. 2007, 21: 755-759. 10.1097/QAD.0b013e328012b838
Hillier SL: The vaginal microbial ecosystem and resistance to HIV. AIDS Res Hum Retroviruses. 1998, 14 (Suppl 1): S17-21.
St John E, Mares D, Spear GT: Bacterial vaginosis and host immunity. Curr HIV/AIDS Rep. 2007, 4: 22-28. 10.1007/s11904-007-0004-y
Dinarello CA: Biologic basis for interleukin-1 in disease. Blood. 1996, 87: 2095-2147.
Cohn JA, Hashemi FB, Camarca M, Kong F, Xu J, Beckner SK, Kovacs AA, Reichelderfer PS, Spear GT: HIV-inducing factor in cervicovaginal secretions is associated with bacterial vaginosis in HIV-1-infected women. J Acquir Immune Defic Syndr. 2005, 39: 340-346. 10.1097/01.qai.0000146599.47925.e0
Olinger GG, Hashemi FB, Sha BE, Spear GT: Association of indicators of bacterial vaginosis with a female genital tract factor that induces expression of HIV-1. Aids. 1999, 13: 1905-1912. 10.1097/00002030-199910010-00013
Spear GT, al-Harthi L, Sha B, Saarloos MN, Hayden M, Massad LS, Benson C, Roebuck KA, Glick NR, Landay A: A potent activator of HIV-1 replication is present in the genital tract of a subset of HIV-1-infected and uninfected women. Aids. 1997, 11: 1319-1326. 10.1097/00002030-199711000-00005
Zariffard MR, Sha BE, Wang QJ, Chen HY, Bremer J, Cohen MH, Spear GT: Relationship of U1 cell HIV-stimulatory activity to bacterial vaginosis and HIV genital tract virus load. AIDS Res Hum Retroviruses. 2005, 21: 945-948. 10.1089/aid.2005.21.945
Al-Harthi L, Spear GT, Hashemi FB, Landay A, Sha BE, Roebuck KA: A human immunodeficiency virus (HIV)-inducing factor from the female genital tract activates HIV-1 gene expression through the kappaB enhancer. J Infect Dis. 1998, 178: 1343-1351. 10.1086/314444
Hashemi FB, Ghassemi M, Roebuck KA, Spear GT: Activation of human immunodeficiency virus type 1 expression by Gardnerella vaginalis. J Infect Dis. 1999, 179: 924-930. 10.1086/314674
Al-Harthi L, Roebuck KA, Olinger GG, Landay A, Sha BE, Hashemi FB, Spear GT: Bacterial vaginosis-associated microflora isolated from the female genital tract activates HIV-1 expression. J Acquir Immune Defic Syndr. 1999, 21: 194-202.
Hashemi FB, Ghassemi M, Faro S, Aroutcheva A, Spear GT: Induction of HIV-1 Expression by Anaerobes Associated With Bacterial Vaginosis. J Infect Dis. 2000, 181: 1574-1580. 10.1086/315455
Mares D, Simoes JA, Novak RM, Spear GT: TLR2-mediated cell stimulation in bacterial vaginosis. J Reprod Immunol. 2007,
Carmody RJ, Chen YH: Nuclear factor-kappaB: activation and regulation during toll-like receptor signaling. Cell Mol Immunol. 2007, 4: 31-41.
Zariffard MR, Novak RM, Lurain N, Sha BE, Graham P, Spear GT: Induction of tumor necrosis factor-alpha secretion and toll-like receptor 2 and 4 mRNA expression by genital mucosal fluids from women with bacterial vaginosis. J Infect Dis. 2005, 191: 1913-1921. 10.1086/429922
Lekkerkerker AN, van Kooyk Y, Geijtenbeek TB: Viral piracy: HIV-1 targets dendritic cells for transmission. Curr HIV Res. 2006, 4: 169-176. 10.2174/157016206776055020
Morrow G, Vachot L, Vagenas P, Robbiani M: Current concepts of HIV transmission. Curr HIV/AIDS Rep. 2007, 4: 29-35. 10.1007/s11904-007-0005-x
St John EP, Martinson J, Simoes JA, Landay AL, Spear GT: Dendritic cell activation and maturation induced by mucosal fluid from women with bacterial vaginosis. Clin Immunol. 2007, 125: 95-102. 10.1016/j.clim.2007.06.004
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Spear, G.T., St John, E. & Zariffard, M. Bacterial vaginosis and human immunodeficiency virus infection. AIDS Res Ther 4, 25 (2007). https://doi.org/10.1186/1742-6405-4-25
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1742-6405-4-25