
Abstract
In this work, a nonlinear deterministic model for schistosomiasis transmission including delays with two general incidence functions is considered. Rigourous mathematical analysis is done. We show that the stability of the disease-free equilibrium and the existence of an endemic equilibrium for the model are stated in terms of key thresholds parameters known as basic reproduction number . This study of the dynamic of the model is globally asymptotically stable if , and the unique endemic equilibrium is globally asymptotically stable when . Some numerical simulations are provided to support the theoretical result with respect to in this paper.
Similar content being viewed by others

1 Introduction
Schistosomiasis is a serious health problem in developing countries. Indeed, despite the remarkable achievements in schistosomiasis control over the past five decades, there are about 240 million people infected worldwide, and more than 700 million people live in endemic areas [1]. There are two patterns of schistosomiasis. We note the urinary schistosomiasis and the intestinal schistosomiasis. The first one is caused by Schistosoma haematobium, when the second is caused by any of the organisms Schistosoma intercalatum, Schistosoma mansoni, Schistosoma japonicum and Schistosoma mekongi. Mathematical modeling of schistosomiasis transmission can help in the development of the strategies for control. Thus, several mathematical models for this disease have been done (see [2–13] and the references therein). In [11], a discrete delay model for the transmission is studied. The delay appears in the incidence term including masse action SI (S: susceptible, I: infectious). It appears that the incidence function form is determinative in the study of the model system. Then, changing the form of the incidence can potentially change the behaviour of the system. In this paper, a mathematical model is derived with a bounded delay distributed and two general incidence functions term f and g. The model described here considers two population hosts, humans and snails, and is structured as follows: Susceptible (uninfected) and infectious humans and susceptible (uninfected) and infected snails. The paper is organized as follows. In Section 2, we present the mathematical model, and we study the mathematical properties of the model system. In Section 3, we derive some results about the basic reproduction number, the disease-free equilibrium and the endemic equilibrium. Section 4 is devoted to the global stability of the disease-free equilibrium. In Section 5, we study the global stability of the endemic equilibrium. Section 6 is devoted to numerical simulation. Finally, in Section 7, we end by a conclusion.
2 The mathematical model
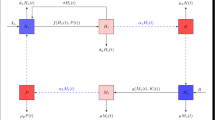
In this section, we derive a mathematical model for the spread of schistosomiasis. Here, we consider human and snail populations. We assume that all newborns are susceptible, and that the infection does not result in death of human and snail populations. Further, it is assumed that a susceptible host became infected only by contact with water, in which there exist cercariae from infected snails, and a susceptible snail became infected by contact with miracidia coming from parasite eggs released in feces and urine of infected hosts (see Figure 1 below).

Transfer diagram for the mathematical model.
We denote by
-
the susceptible (uninfected) human population size;
-
the infected human population size (infectious humans);
-
the susceptible (uninfected) snail population size;
-
the infected and shedding snail population size (shedding snail population size).
We also denote by
-
the recruitment rate of susceptible humans;
-
the recruitment rate of susceptible snails;
-
the per capita natural death rate of humans;
-
the per capita natural death rate of snails;
-
the transit time from cercaria in water to schistosomule in a human host;
-
the transit time from parasite eggs to miracidia to infect a snail;
-
and the Lebesgue integrable functions, which give the relative infectivity of snails and humans (respectively) of different infection ages.
Note that the support of and has a positive measure in any open interval having supremum h, so that the interval of integration is not artificially extended by concluding with an interval, for which the integral is automatically zero. On the other hand, we choose in the model two real numbers α and γ, so that and .
As general as possible, the incidence functions f and g must satisfy technical conditions. Thus, we assume that
-
H1 f and g are non-negative functions on the non-negative quadrant,
-
H2 for all , and .
Remark 2.1 f and g are two incidence functions, which explain the contact between two species. Therefore, f and g are non-negative. Note also that when there is no one infected in the human and snail populations, then the incidence functions are equal to zero. The incidence functions are also equal to zero, when there is no one susceptible in the human and snail populations.
Let us denote by and the partial derivatives of f with respect to the first and to the second variable and and those of g with respect to the first and to the second variable. For mathematical simplicity, we shall make now a simplification that will allow us to carry out an analysis, namely we assume that the disease-induced death rate is neglected. Using the notations and , the model equations are given as follows
We assume that system (2.1) holds with given initial conditions
where . We also define the sup norm on as , . Standard theory of functional differential equations (see [14]) can be used to show that solutions of system (2.1) exist and are differentiable for all .
The delay is inspired by the life history of the schistosomiasis. Indeed, it is possible that some hosts or intermediate hosts (snails) die due to natural death during the incubation period, respectively (see [11]).
Theorem 2.2 The positive orthant
is positively invariant for system (2.1).
To prove Theorem 2.2, we need the following result.
Theorem 2.3 [15]
Let be a differentiable function, and let . Let be the vector field, and let G be the closed set such that for all . If for all , then the set G is positively invariant.
Proof of Theorem 2.2 Let
We will prove that is positively invariant. Then let
L is differentiable, and for all . The vector field on is
Then . This proves that is positively invariant. Similarly, we prove that , , are positively invariant. Then is positively invariant for system (2.1). □
Therefore, the model is mathematically well posed and epidemiologically reasonable since all the variables remain non-negatives for all .
Theorem 2.4 Assume that and .
There exists such as all feasible solutions of model system (2.1) enter the set
Proof Let . Adding the first two equations of (2.1), we get
According to [16], it follows that
where . Thus, as , .
Similarly, we prove that . □
3 Basic reproduction number, disease-free equilibrium and endemic equilibrium
The disease-free equilibrium is given by
Proposition 3.1 The basic reproduction number for model system (2.1) is defined by
Proof Rewriting the system (2.1) as
where
Now, making the assessment in each compartment j
where
-
denote the rate of appearance of new infections in each class j,
-
denote the rate of transfer into each class j by all other means,
-
denote the rate of transfer out of each class j.
Let , and let be the healthy population and the infected population,
Then,
and
On , we get
and
Thus, we obtain
According to [17], we conclude the proof. □
The basic reproduction number represents the average number of new case generated by a single infected individual in a completely susceptible population (see [18]).
Theorem 3.2 If , then is locally asymptotically stable.
Proof Suppose that . Since H2 holds then, and for all and . It follows that the linearization of system (2.1) at is
Now, we replace by into (3.11) to get
After that, we rearrange before cancelling from each term, and we obtain
where
Denote by
then there exist non-zero solutions if and only if . It follows that the characteristic equation is given by
Now, we will show that all solutions λ of this equation have a negative real part. By contradiction, suppose that λ has non-negative real part. With this in mind, we have and . Also
This implies that λ cannot be a solution of the characteristic equation. Hence, all eigenvalues have negative real part, and then is locally asymptotically stable. □
Now, we will study the behaviour of system (2.1) when .
Theorem 3.3 If , then there exists an endemic equilibrium.
Proof Let . Setting the right-hand side of system (2.1) equal to zero, we know that is a positive equilibrium if and only if
Combining the two first equations and the two last equations of (3.17) gives
Let
and
Now, we define the continuous function ϕ by
Hence, it follows that any solution of equation in the set corresponds to an equilibrium, with , that is an equilibrium. Since H2 holds, then and . Then the sufficient condition for equation to have a solution in is that ϕ increasing at 0. This implies that an endemic equilibrium exists if
where
Note that . Then, inequality (3.22) becomes
Hence,
equivalently, we have
Thus, . □
4 Global stability of the disease-free equilibrium
In this section, we study the global behaviour of the disease-free equilibrium. For that, we assume that
-
H3 for all , and ,
-
H4 and .
We have the following result.
Theorem 4.1 Let . Assume that H3 and H4 hold, and , then the disease-free equilibrium is globally asymptotically stable if .
Proof We consider the Lyapunov function
Differentiating V with respect to time, we obtain
Since H3 holds and , we get
Adding and subtracting the quantity , we have
Using hypothesis H4, we obtain
For , we get
with equality only if and . According to LaSalle’s extension to Lyapunov’s method [19], the limit set of each solution is contained in the largest invariant set, for which and , which is the singleton . This means that the disease-free equilibrium is globally asymptotically stable on . □
5 Global stability of the endemic equilibrium
In this section, we assume that f and g satisfies the conditions
-
H5 for all , and ,
-
H6 for all , and .
Theorem 5.1 Assume that H5 and H6 hold, then if the endemic equilibrium is globally asymptotically stable.
Proof Consider system (2.1) at , we get
Let
We can see that the function for all , and it has the strict global minimum . Now, define the functions
Note that , , and with equality only if , , and . Now, we consider the Lyapunov function
where and . To evaluate , we will calculate separately the different derivatives , , and ,
Then we use the first equation of (5.1) to get
After that, we evaluate ,
Then, using the third equation of (5.1), we obtain
Then, combining (5.6) and (5.8), we get
where .
Adding and subtracting the quantity , we obtain
Next, we calculate ,
Then, we use the second equation of (5.1) to get
Now, we evaluate ,
Then, using the fourth equation of (5.1), we obtain
Then, combining (5.12) and (5.14), we get
where .
Adding and subtracting the quantity , we obtain
Since H5 holds, then and . It follows that
Since H6 holds, it follows that
for all with equality only for , , and . Hence, the endemic equilibrium is the only positively invariant set of system (2.1) contained in . Then it follows that is globally asymptotically stable on (see [19]). □
6 Numerical simulation
In this section, we derive the computation work that supports our study. In this computation, the functions f and g are chosen as follows: and (mass action). Two different cases of computational simulations are studied: in the first case (see Figure 2), , while in the second case (see Figure 3), . The parameters values used are the following (see [20]): , , , . We also use the delays parameters . For the first case, we use and , which give . In the second case, we use and to get .

Case, where .

Case, where .
7 Conclusion
In this paper, a deterministic model of transmission of schistosomiasis with two general nonlinear incidence functions including distributed delay is derived. The global behaviour of the model system was studied. We proved that, if holds, then the disease-free equilibrium is globally asymptotically stable, which implies that the disease fades out from the population. If , then there exists a unique endemic equilibrium which is globally asymptotically stable, and this implies that the disease will persist in the population. This result suggests that the latent period in infection affects the prevalence of schistosomiasis, and it is an effective strategy on schistosomiasis control to lengthen in prepatent period on infected definitive hosts by drug treatment, for example.
Threshold analysis of the basic reproduction number shows that the use of public health education campaign could have positive, more determinant impact on the control of the schistosomiasis. Overall, an effective education campaign which focuses on drug treatment with reasonable coverage level could be helpful for countries concerned with the disease.
References
World Health Organization. http://www.who.int/schistosomiasis/en/. Available on May, 2012.
Allen EJ, Victory HD: Modelling and simulation of a schistosomiasis infection with biological control. Acta Trop. 2003, 87: 251-267. 10.1016/S0001-706X(03)00065-2
Anderson RM, May RM: Prevalence of schistosome infections within molluscan populations: observed patterns and theoretical predictions. Parasitology 1979, 79: 63-94. 10.1017/S0031182000051982
Anderson RM, May RM: Helminth infections of humans: mathematical models, population dynamics, and control. Adv. Parasitol. 1985, 24: 1-101.
Cohen JE: Mathematical models of schistosomiasis. Annu. Rev. Ecol. Syst. 1977, 8: 209-233. 10.1146/annurev.es.08.110177.001233
Feng Z, Li C, Milner FA: Schistosomiasis models with density dependence and age of infection in snail dynamics. Math. Biosci. 2002, 177-178: 271-286.
Feng Z, Li C, Milner FA: Schistosomiasis models with two migrating human groups. Math. Comput. Model. 2005, 41(11-12):1213-1230. 10.1016/j.mcm.2004.10.023
Feng Z, Milner FA: A new mathematical model of schistosomiasis. Innov. Appl. Math. In Mathematical Models in Medical and Health Science. Vanderbilt Univ. Press, Nashville; 1998:117-128. (Nashville, TN, 1997)
Macdonald G: The dynamics of helminth infections, with special reference to schistosomiasis. Trans. R. Soc. Trop. Med. Hyg. 1965, 59: 489-506. 10.1016/0035-9203(65)90152-5
Nasell I: A hybrid model of schistosomiasis with snail latency. Theor. Popul. Biol. 1976, 10: 47-69. 10.1016/0040-5809(76)90005-8
Yang Y, Xiao D: A mathematical model with delays for schistosomiasis. Chin. Ann. Math., Ser. B 2010, 31(4):433-446. 10.1007/s11401-010-0596-1
Woolhouse MEJ: On the application of mathematical models of schistosome transmission dynamics. I. Natural transmission. Acta Trop. 1991, 49: 1241-1270.
Woolhouse MEJ: On the application of mathematical models of schistosome transmission dynamics. II. Control. Acta Trop. 1992, 50: 189-204. 10.1016/0001-706X(92)90076-A
Hale JK, Verduyn Lunel S: Introduction to Functional Differential Equations. Springer, Berlin; 1993.
Bony JM: Principe du maximum, inégalite de Harnack et unicité du problème de Cauchy pour les opérateurs elliptiques dégénérés. Ann. Inst. Fourier (Grenoble) 1969, 19(1):277-304. 10.5802/aif.319
Birkhoff G, Rota GC: Ordinary Differential Equations. Ginn, Boston; 1982.
Van den Driesche P, Watmough J: Reproduction numbers and subthreshold endemic equilibria for the compartmental models of disease transmission. Math. Biosci. 2002, 180: 29-48. 10.1016/S0025-5564(02)00108-6
Anderson RM, May RM: Infectious Disease of Human. Oxford University Press, Oxford; 1991:17-19.
LaSalle JP: The Stability of Dynamical Systems. SIAM, Philadelphia; 1976.
Feng Z, Milner FA, Eppert A, Minchella DJ: Estimation of parameters governing the transmission dynamics of schistosomes. Appl. Math. Lett. 2004, 17(10):1105-1112. 10.1016/j.aml.2004.02.002
Acknowledgements
The authors express their deepest thanks to the editor and an anonymous referee for their comments and suggestions on the article.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
All authors carried out the paper. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Guiro, A., Ouaro, S. & Traore, A. Stability analysis of a schistosomiasis model with delays. Adv Differ Equ 2013, 303 (2013). https://doi.org/10.1186/1687-1847-2013-303
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1687-1847-2013-303