
Abstract
Background
Human Leukocyte Antigen (HLA)-E is a non-classical class I HLA molecule that can be stabilized by ligands donated by other classical (HLA-A, -B, -C) and non-classical (HLA-G) family members. HLA-E engages a variety of immune receptors expressed by cytotoxic T lymphocytes (CTLs), Natural killer (NK) cells and NK-CTLs. In view of the opposing outcomes (activation or inhibition) of the different HLA-E receptors, the preferred role (if any) of HLA-E expressed in vivo on tumor cells remains to be established.
Methods
Taking advantage of MEM-E/02, a recently characterized antibody to denatured HLA-E molecules, HLA-E expression was assessed by immunohistochemistry on an archival collection (formalin-fixed paraffin-embedded) of 149 colorectal primary carcinoma lesions paired with their morphologically normal mucosae. Lymphoid infiltrates were assessed for the expression of the HLA-E-specific, inhibitory, non-rearranging receptor NKG2A.
Results
High HLA-E expression did not significantly correlate with the expression of classical HLA-B and HLA-C molecules, but it did correlate with high expression of its preferential ligand donor HLA-A. In addition, it correlated with lymphoid cell infiltrates expressing the inhibitory NKG2A receptor, and was an independent predictor of good prognosis, particularly in a subset of patients whose tumors express HLA-A levels resembling those of their paired normal counterparts (HLA-A). Thus, combination phenotypes (HLA-Elo-int/HLA-AE and HLA-Ehi/HLA-AE) of classical and non-classical class I HLA molecules mark two graded levels of good prognosis.
Conclusions
These results suggest that HLA-E favors activating immune responses to colorectal carcinoma. They also provide evidence in humans that tumor cells entertain extensive negotiation with the immune system until a compromise between recognition and escape is reached. It is implied that this process occurs stepwise, as predicted by the widely accepted 'immunoediting' model.
Similar content being viewed by others

Background
Human Leukocyte Antigen (HLA)-E is a cell surface, non-classical Major Histocompatibility class I molecule recognized by immune receptors expressed by cytotoxic T lymphocytes (CTLs), Natural Killer (NK) cells, and the more recently described subset of NK-CTLs. These receptors are either inhibitory or activating [1–3].
Inhibition, on the one hand, results from the engagement of the NKG2A receptor with HLA-E heavy chains that have been stabilized upon heterotrimeric assembly with their light chain subunit, called β2-microglobulin (β2m), and peptide ligands derived from the signal sequences of 'permissive' class I heavy chains, both classical (HLA-A, -B, -C) and non-classical (HLA-G). Activation, on the other hand, results from: (a) the competitive relief of NKG2A-mediated inhibition upon HLA-E assembly with peptides from donor proteins other than HLA class I; (b) the direct engagement of the activating NKG2C receptor isoform; and (c) antigen-specific recognition through the T cell receptor (TcR) expressed by NK-CTLs [1–4]. Balancing and integration of opposing signals (often dubbed activation-inhibition) is not unique of HLA-E, but is indeed widely adopted to control cytotoxic responses and regulate complex immune networks. Thus, HLA-E may provide key information to understand how virus-infected [5] and tumor cells walk the thin line between immune surveillance and immune evasion.
HLA-A, -B, -C down-regulation has been viewed as a major subterfuge to deceive T cells [6] for some time now, but it is unlikely to provide a comprehensive explanation of immune evasion, since it impairs ligand donation to HLA-E and the direct engagement of inhibitory NK receptors [1, 7–9]. Accordingly, several immunohistochemical studies failed to confirm an association between HLA-A, -B, - C loss and poor prognosis [10–12], and our own studies were consistent with activation-inhibition models [13–16]. We showed that early-passage melanoma, breast carcinoma, and lung carcinoma cells, like virus-infected cells, avoid both extremes of overly low or high HLA class I expression, which would expose them to lysis by NK and CTLs, respectively. Similar 'low profile' HLA phenotypes were also observed in vivo, in colorectal carcinoma lesions, and were associated with a favorable prognosis, whereas extreme down-and up-regulation with respect to the normal autologous mucosa were rare and associated with a poor prognosis, particularly when involving the HLA-A locus [16]. Possibly, these altered HLA phenotypes mark tumor cells refractory to immune elimination.
In an effort to characterize monoclonal antibodies (mAbs) to HLA-E, we found that MEM-E/02 binds a linear epitope highly restricted to the HLA-E polypeptide and fully available upon denaturation [17]. Using MEM-E/02, we recently observed that HLA-E is constitutively co-expressed with HLA-A, -B, -C molecules in a fraction of neoplastic tissues and on the surface of most cultured tumor cell lines. In these conditions, HLA-E is functional [18]. In the present report, we describe the use of MEM-E/02 on archival collections of formalin-fixed/paraffin-embedded colorectal adenocarcinoma tissues and their paired, morphologically normal mucosae. We found that overall HLA-E expression is a significant prognosticator, and in combination with HLA-A expression and the presence of NKG2A infiltrating lymphoid cells provides a clue to understand tumor immune surveillance in vivo.
Methods
Patients and histological specimens
Patients (149 cases, 71 men and 78 women, median age 64, range 34-90 years) were radically resected for primary colorectal adenocarcinoma between 1988 and 2000. Pathological staging [19], performed according to the American Joint Committee on Cancer (AJCC)/International Union Against Cancer (UICC), identified 98 and 51 tumors at stages II and III, respectively. The latter were treated by adjuvant chemotherapy. According to the WHO classification, well (G1), moderately (G2), and poorly (G3) differentiated tumors were 10, 114, and 25, respectively. T2, T3 and T4 tumors were 26, 63, and 60, respectively. The present archival tissue collection largely overlaps (n = 126) with that previously described [16]. Uninvolved mucosal tissues were obtained from the same tissue block as the tumor lesion in 108 cases, and from a distinct tissue block in 41 cases. No patients were lost to clinical follow-up (continuously updated), and the median follow-up was 71.0 (range 3.0 - 175.1) months, with 45 relapses and 37 deaths due to cancer-related causes. This study was approved by the Ethical Committee.
Antibodies and Immunohistochemistry
MEM-E/02 is the only available mAb to HLA-E with an extensive biochemical characterization. It binds a linear epitope present on HLA-E but not HLA-A, -B, -C, -F or -G heavy chains [17]. HCA2 binds all the denatured HLA-A alleles except A24, and crossreacts with the rare HLA-B73 allele, HLA-G, HLA-E and HLA-F [20, 21]. However, HLA-E and HLA-F bind weakly and almost exclusively under native conditions [21], and HLA-G accounts for less than 5% of the total HCA2 reactivity in a large cell panel [15]. HC10 binds HLA-B and weakly crossreacts with HLA-C, whereas L31 binds HLA-C and a few crossreacting HLA-B alleles [20, 22, 23]. CD8 and CD56 were identified by mAbs 1A5 and 1B6 (Novocastra, UK), respectively. The goat polyclonal antibody N-19 to NKG2A was from Santa Cruz (CA, USA). Sections (5-μm thick) from routinely formalin-fixed (18-24 h) paraffin-embedded tissue blocks were stained by Immunohistochemistry (IHC). Antigen retrieval for HCA2, HC10, L31, CD8 and CD56 staining was carried out as described [23]. For MEM-E/02 and NKG2A staining, slides were dewaxed, rehydrated and heated in a water bath at 96°C for 40 minutes in EDTA (pH 8.0), followed by cooling at room temperature. Staining was revealed by a supersensitive streptavidin-biotin immunoperoxidase system (Biogenex, Menarini, Italy) for all antibodies. In addition, NKG2A staining was also revealed by a Goat AP-Polymer Kit (Biocare Medical, CA, USA). The chromogenic substrate was 3,3'-diaminobenzidine for all stainings. In addition, NKG2A staining was also revealed by Warp Red (Biocare Medical). Preliminary specificity assessments were performed on a panel of HLA-E-positive and HLA-E-negative cell lines [17] fixed and embedded as above. Different staining sessions included repeated staining of an internal normalization control (a section from a known specimen) to ensure day-to-day consistency. Because weak staining was invariably associated with the presence of negative areas, whereas strong staining tended to be homogeneous (this applies to both normal and tumor tissues), staining heterogeneity and staining intensity were factored into a single scale (termed 'absolute' intensity score hereafter). The absolute intensity score may acquire one of four possible values: 0 (undetectable or faint in <20% of the cells); 1 (faint to weak in 20% but ≤70% of the cells); 2 (weak to moderate in >70% of the cells); 3 (intense in>70% of the cells). In analogy with our previously cited study [16], we calculated an additional intensity scale (termed 'relative' hereafter) by subtracting the absolute score of the normal mucosa from that of its paired colon carcinoma specimen. Subtraction of two series of scores, each graded on a four-digit scale, resulted in the assignment of one of 7 possible algebraic values (-3, -2, -1, 0, +1, +2, +3) to each normal-neoplastic pair. Intermediate (-1, 0, and +1) and extreme (-3, -2, +2, and +3) relative scores identify pairs in which the tumor is similar to and deviates from, respectively, its normal paired mucosa. CD3, CD56 and NKG2A infiltrates were assessed in six randomly chosen high power fields (HPF, ×400), and expressed as either the percentage of positive cells in the total infiltrate, or the average number of positive cells in each HPF.
Statistical analysis
Descriptive statistics were used to summarize pertinent study information. The Spearman rho correlation was used to determine relationships between parameters. The Hazard risk and the confidence limits were estimated for each variable using the Cox univariate model. Significance was defined at the p < 0.05 level. A multivariate Cox proportional hazard model was also developed using stepwise regression (forward selection) by selecting those predictive variables that were significant upon univariate analysis. Enter limit and remove limit were p = 0.10 and p = 0.15 respectively. Survival was calculated by the Kaplan-Meier product-limit method from the date of surgery until the time of death (overall survival; OS), relapse (disease-free survival; DFS), or last visit (OS and DFS), whichever applicable. The log-rank test was used to assess differences between subgroups. Significance was defined at the p < 0.05 level. Statistical analysis was carried out by the SPSS v. 13.0 (SPSS, Milan, Italy) software. Optimal cut-offs of NKG2A immunoreactivity defining two groups of patients with significantly different MEM-E/02 intensity scores were found using the Receiver Operating Characteristic (ROC) curve analysis.
HLA-A, -B, -C typing
Genomic DNAs were extracted from the available frozen tumor tissue samples (n = 29), and typed for HLA-A, -B, and -C using sequence-specific primers (Olerup-SSP, Genovision, Vienna, Austria).
Results
Analysis of MEM-E/02 immunoreactivity in colorectal carcinomas and paired mucosa specimens
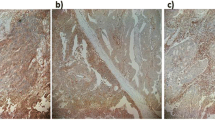
Colorectal carcinomas from stage II/III patients (n = 149), each paired with morphologically normal colonic mucosa from the same patient, were stained by immunohistochemistry with the HLA-E-specific MEM-E/02 antibody. Representative examples of staining grades (recorded as absolute scores, see Materials and Methods) are provided in Figure 1A-C. HLA-E was detected in nearly all the specimens, approximately half of which displayed the highest absolute score of 3. High absolute scores were most frequent also with HCA2 (to HLA-A), HC10 (to HLA-B) and L31 (to HLA-C), as shown in Additional file 1, Fig. S1A. Similar results were obtained by plotting the staining intensities of the normal mucosae (not shown). Thus, HLA-E is expressed in essentially all normal and neoplastic colonic tissues, often at high levels.

Immunohistochemical staining of formalin-fixed, paraffin-embedded colorectal carcinoma tissues. A-C: Representative reactivity scores of MEM-E/02 to HLA-E (see Materials and Methods): A = 1, B = 2, C = 3. D: staining of lymphoid infiltrates with an antibody to the HLA-E-specific receptor NKG2A. Magnification: 10×, 40×, 20×, and 40× respectively. Scale bar = 100 μm.
Next, we calculated the seven-digit relative intensity scores (see Materials and Methods) of paired immunohistochemical determinations. Representative results (Additional file 1, Fig. S1B) demonstrate that tumors resembling their normal paired mucosa (relative scores: +1, 0, and -1) were far more numerous than tumors undergoing consistent up-regulation or down-regulation (relative scores: -2, -3 and +2, +3), a finding that applies to all the tested HLA class I molecules: HLA-E, HLA-A, HLA-B and HLA-C.
Finally, flow cytometry of 3 representative colorectal carcinoma cell lines provided evidence that at least some of the denatured HLA-A and HLA-E molecules detected by HCA2 and MEM-E/02 on archival tissues derive from native molecules that are co-expressed on the surface of live cells (Additional file 1, Fig. S2).
Co-expression of HLA-E and permissive class I alleles
It has been shown that signal sequence-derived peptides from permissive alleles (most HLA-A and some HLA-C) carry methionine at P2, whereas peptides from non-permissive alleles (most HLA-B) carry threonine [24, 25]. On this basis, absolute HLA-E levels are expected to correlate with (a) the levels of co-expressed products from at least some HLA-A, -B, and -C loci, and (b) the number of permissive alleles carried at the DNA level. To verify prediction (a), the staining intensities of MEM-E/02 (to HLA-E) were correlated with the staining intensities of HCA2, HC10, and L31 (preferentially reactive with HLA-A, HLA-B and HLA-C, respectively) in the 149 tumor specimens. A significant (Table 1, p = 0.01) correlation was detected between HLA-E and HLA-A, but not HLA-B or HLA-C. To verify prediction (b), 29 high-quality genomic DNAs (required for reliable allele-specific PCR) from colon carcinoma lesions were typed for HLA-A, -B, -C. From 3 to 6 permissive alleles (alleles and signal sequences listed at <http://hla.alleles.org/data/index.html>) were identified in each DNA. As in the complete case collection, HLA-E and HLA-A staining intensities did correlate also in this smaller subset (Table 1, p = 0.02; also see Additional file 1, Fig. S3A). In addition, there was a weak non-significant trend for correlation between the number of the putative HLA-A/-B/-C donor alleles and HLA-E staining intensity (Additional file 1, Fig. S3B). We conclude that HLA-E and at least its preferred ligand donors (from the HLA-A locus) are coordinately expressed in colorectal carcinoma lesions in vivo.
Association between HLA-E expression and survival
Next, univariate analysis and multivariate analysis of DFS and OS (Cox model) were performed on the entire series of 149 stage II/III colorectal cancer patients. In agreement with our previous study [16], advanced tumor stage and extreme up-/down-regulation of HCA2-reactive (HLA-A) molecules (relative scores of +3, +2, -3, and -2) were significant and independent predictors of both DFS and OS (Table 2, top two lines in both sections), whereas the absolute reactivity scores of HCA2, HC10 and L31 (Table 2, last three lines in both sections) as well as the relative reactivity scores of HC10 and L31 (not shown) were not. Contrary to HCA2, and quite surprisingly, the relative MEM-E/02 scale did not identify any significant prognostic category (not shown).
However, a high MEM-E/02 absolute staining intensity score of 3 did display a significant correlation with favorable prognosis upon both univariate analysis and multivariate analysis (Table 2, third line from top in both sections). Accordingly, Kaplan-Meier analysis (Figure 2A and 2B) was consistent with significantly longer DFS and OS in patients bearing tumors with high levels of absolute MEM-E/02 reactivity (HLA-Ehi).

Kaplan-Meier survival curves of patients with colorectal carcinoma. Five-year disease-free (A) and overall (B) survival rates of 149 stage II-III colorectal cancer patients according to the MEM-E/02 (HLA-E) absolute immunohistochemical score (MEM-E/02hi= 3; MEM-E/02lo/int: 0, 1 and 2). Five-year disease-free (C) and overall (D) survival of stage II-III colon carcinoma patients according to the HCA2 (HLA-A) relative immunohistochemical score (0, -1, +1 vs +2, +3, -2, -3). Patients (n = 94) with an HCA2 reactivity mimicking that seen in the normal paired mucosa (relative scores 0, -1 and +1) were sorted into MEM-E/02hi (absolute reactivity score = 3) and MEM-E/02lo/int (0, 1, and 2). The percent survival, shown in ordinate, is analytically displayed for the different groups at the end of the observation period, along with the statistical significance (p) of the observations.
Next, we investigated whether a combined evaluation of relative HCA2 reactivity and absolute MEM-E/02 reactivity might improve the accuracy of outcome prediction. In the patients with unfavorable HCA2 phenotypes (extreme HLA-A up-/down-regulation), absolute HLA-E expression did not significantly associate with 5-year DFS and OS (not shown), suggesting that 'abnormal' HCA2 reactivity is a dominant prognostic factor, and the influence of HLA-E in this tumor subset is negligible. In contrast (Figure 2C and 2D), among patients bearing tumors with HLA-A levels similar to that of the normal autologous mucosa (that are by themselves associated with favorable prognosis), high and low/intermediate HLA-E expression levels further discriminated two numerically equivalent populations (n = 49 vs 45) significantly differing in their chances of relapse and death within 5 years.
Correlation between HLA-E expression in tumors and NKG2A expression in lymphoid infiltrates
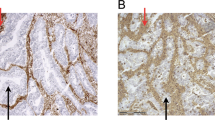
Sixteen lesions expressing the highest HLA-E levels (MEM-E/02 score = 3), and 12 lesions expressing the lowest levels (score = 1) were assessed for NKG2A (Figure 1D), CD8 and CD56 infiltrates (not shown). Up to 5% and 20% of colorectal carcinoma infiltrates were reactive with NKG2A and CD8, respectively. In contrast, only scattered CD56 cells (no more than 2 per microscopic field) were detected. As shown by a representative stain of serial sections (Figure 3), the distribution of NKG2A-positive and CD8-positive cells partly overlaps.

Immunohistochemical staining for CD8 and NKG2A. Two consecutive sections of a colorectal carcinoma lesion displaying a strongly and homogeneously positive (MEM-E/02 score = 3) neoplastic mucosa were stained by mAb 1A5 to CD8 (A and C) and the polyclonal antibody N-19 to NKG2A (B and D). Different chromogens reveal CD8 and NKG2A cells in a large peritumoral infiltrate. Counterstaining with Haematoxilin. Magnification 10 × (panel A and B) and 20 × (panel C and D). Scale bar = 100 μm.
Upon ROC analysis there was no significant correlation between CD8 infiltrate (either the extent or number of positive cells) and HLA-E expression. In contrast, high and low levels of the HLA-E ligand in tumor tissues correlated with high and low numbers, respectively, of infiltrating lymphoid cells expressing its specific receptor NKG2A (Additional file 1, Table S1; p = 0.04).
Discussion
Although intracellular transport of HLA-E is believed to be quite inefficient [26], colorectal carcinoma cells (like most neoplastic cell lines) co-express on the surface HLA-E and classical HLA class I molecules, including HLA-A ([18] and Additional file 1, Fig. S2).
The present immunohistochemical study of colorectal carcinoma lesions with the MEM-E/02 antibody [17] reveals that high expression of HLA-E significantly correlates with high expression of HLA-A, but not HLA-B or HLA-C. Since HLA-E ligands donated by most HLA-B and at least some HLA-C alleles are poorer NKG2A inhibitors than those donated by HLA-A alleles [27], in vitro mechanisms of ligand donation and HLA-E stabilization appear to be recapitulated in vivo.
Ligand donation by permissive HLA class I alleles is a well-known strategy to signal integrity of the so-called antigen processing machinery, and prevent inappropriate immune lysis upon HLA-E:NKG2A inhibition [1, 2]. Consistent with HLA-E enabling active scrutiny by the immune system (rather than immunoevasive responses), we show herein that HLA-Ehi phenotypes are associated with a lymphoid infiltrate (mainly CD8) enriched in the HLA-E-specific inhibitory receptor NKG2A, and a favorable prognosis.
Similar to previous studies [28, 29], in which the extent of the colorectal carcinoma lymphoid infiltrate was correlated with a favorable prognosis, only scattered NK cells were detected herein. However, this does not obviously rule out a role of infiltrating NK cells in vivo.
Interestingly, we found that high HLA-E expression is associated per se with a favorable outcome, but it is the combination between high HLA-E (on an absolute scale) and similar HLA-A levels in the tumor compared to the normal mucosa (on a relative expression scale) that results in the precise identification of graded levels of prognosis.
Also of note, HLA-A clearly dominates the prognostic landscape both in quantitative and qualitative terms. Quantitatively, HLA-A identifies a wider prognostic gap between favorable and poor outcomes. Qualitatively, HLA-A was shown previously to be an early-stage prognosticator [16], whereas HLA-E is shown herein to influence prognosis particularly within a cohort of patients pre-selected for a favorable outcome through HLA-A phenotyping.
The easiest interpretation of these results involves the assignment of the observed HLA-A and HLA-E phenotypic variants to the three classical phases (elimination, equilibrium and escape; boxes n. 1, 2 and 3 in Figure 4) of the widely accepted 'immunoediting' theory. This theory was originally proposed by Dunn et al.[30] to interpret murine models of tumor escape. We are not aware of previous attempts to classify specific class I HLA phenotypes in the context of stepwise immunoediting. We propose that in the first immunoediting phase (box 1, elimination), HLA-Ahi and HLA-Alo phenotypes are suppressed, for instance by anti-tumor CTL and NK cells equipped with functionally dominant activating and inhibitory receptors, respectively. As described by us several years ago [13, 14], this results in the selection of 'low profile' (HLA-A phenotypes, e.g. colorectal carcinoma cells expressing HLA-A levels similar to those of their normal counterparts. Next, two phenotypes arise characterized by different HLA-E levels (box 2, immunoediting): HLA-A/HLA-Elo/int and HLA-A/HLA-Ehi. Low/intermediate and high HLA-E levels are hypothesized to result: (a) the former from limited ligand donation and HLA-E stabilization, as a result of HLA-Ahi suppression during the previous elimination phase; (b) the latter from HLA-A-independent events of ligand donation. Why both might incite anti-tumor immune responses, at two different levels, could be explained by relief of lytic inhibition and/or triggering immune responses. There are precedents for both. Relief of inhibition by competitive binding of non-HLA-derived ligands is known to favor NK lysis [4]. Direct triggering of cytotoxic CD8 lymphocytes has been described to result from the presentation of a non-conventional tumor antigen in the context of an undefined non-classical class I molecule [31].

Class I HLA expression, immunoediting, and survival through colorectal carcinoma stages. Stepwise selection of HLA phenotypes results in distinct tumor variants populating the three classical steps (elimination, equilibrium and escape; shaded boxes) of immunoediting. Equilibrium is defined herein quasi-equilibrium. Unlike 'true' equilibrium, that corresponds to occult cancer and dormancy [30], and was elusive even in experimental models of mouse tumorigenesis [39], quasi-equilibrium refers to edited tumor cells embedded in a clinically evident tumor mass, sufficiently large to be immunophenotyped in a human pathology facility.
Finally (box 3, escape), extreme HLA-A phenotypes (HLA-A↑↓)1, very different from those seen in the normal mucosa, and initially suppressed during the elimination phase, may (re)appear. Having forced all the immune checkpoints, these phenotypes are not surprisingly associated with the poorest outcomes regardless of HLA-E expression, found to be irrelevant in this group.
Thus, we propose a dual outcome model whereby HLA-E expression, far from being a manifestation of immune inhibition and escape, represents instead a prerequisite for active immunological scrutiny of edited HLA tumor variants on the one hand, and triggering immune responses on the other.
Recently, Levy et al. reported poor MEM-E/02 reactivity in most colorectal carcinomas, and an association between high HLA-E expression, detected in just 8 lesions, and poor prognosis [32]. Poor reactivity is difficult to reconcile with the high levels of HLA-E detected by us with the same MEM-E/02 antibody in most lesions, and the prognostic associations proposed by Levy et al. are based on a limited number of cases. Another example of HLA-E association with poor prognosis is a recent study on early-stage breast carcinoma [33].
Our results better agree with two cDNA profiling studies of advanced melanoma in which high levels of HLA-E transcripts contributed molecular signatures of favourable prognosis [34, 35], and a recent study on glioblastoma [36]. Possibly, the balance between activating and inhibitory functions of HLA-E may differ in different tumor histotypes. Additional work is required to investigate these issues in the context of activation-inhibition models of tumor immune surveillance.
Conclusions
In summary, this is one of the few reports [32, 33, 37] addressing the prognostic significance of non-classical class I molecules in colorectal carcinoma. Additionally, we provide evidence (albeit indirect) for a functional interplay between classical and non-classical HLA molecules in vivo. This is also the first report to suggest in situ engagement of tumor cells and T lymphocytes expressing a defined ligand:non-rearranging immune receptor pair.
Note added in proof
At the time of proof-editing a paper [38] appeared in PubMed describing a poor prognosis associated with HLA-E expression in colorectal carcinoma and NKG2A expression in the lymphoid infiltrate.
References
Braud VM, Allan DSJ, O'Callaghan CA, Söderström K, D'Andrea A, Ogg GS, Lazetic S, Young NT, Bell JI, Phillips JH, Lanier LL, McMichael AJ: HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature. 1998, 391: 795-799. 10.1038/35869.
Rodgers J, Cook R: MHC class Ib molecules bridge innate and acquired immunity. Nat Rev Immunol. 2005, 5: 459-471. 10.1038/nri1635.
Moretta L, Romagnani C, Pietra G, Moretta A, Mingari MC: NK-CTLs, a novel HLA-E-restricted T-cell subset. Trends Immunol. 2003, 24: 136-143. 10.1016/S1471-4906(03)00031-0.
Michaelsson J, Teixeira de Matos C, Achour A, Lanier LL, Kärre K, Soderstrom K: A signal peptide derived from hsp60 binds HLA-E and interferes with CD94/NKG2A recognition. J Exp Med. 2005, 196: 1403-1414.
Cohen GB, Gandhi RT, Davis DM, Mandelboim O, Chen BK, Strominger JL, Baltimore D: The selective downregulation of class I major histocompatibility complex proteins by HIV-1 protects HIV-infected cells from NK cells. Immunity. 1999, 10: 661-671. 10.1016/S1074-7613(00)80065-5.
Smith MEF, Bodmer WF, Bodmer JG: Selective loss of HLA-A, B, C locus products in colorectal adenocarcinoma. Lancet. 1988, I: 823-824.
Lee N, Goodlett DR, Ishitani A, Marquardt H, Geraghty DE: HLA-E surface expression depends on binding of TAP-dependent peptides derived from certain HLA class I signal sequences. J Immunol. 1998, 160: 4951-4960.
Tomasec P, Braud VM, Rickards C, Powell MB, McSharry BP, Gadola S, Cerundolo V, Borysiewicz LK, McMichael AJ, Wilkinson GWG: Surface expression of HLA-E, an inhibitor of natural killer cells, enhanced by human cytomegalovirus gpUL40. Science. 2000, 287: 1031-1033. 10.1126/science.287.5455.1031.
Ulbrecht M, Martinozzi S, Grzeschik M, Hengel H, Ellwart JW, Pla M, Weiss EH: The human cytomegalovirus UL40 gene product contains a ligand for HLA-E and prevents NK cell-mediated lysis. J Immunol. 2000, 164: 5019-5022.
Gudmundsdóttir I, Jónasson JG, Sigurdsson H, Olafsdóttir K, Tryggvadóttir L, Ogmundsdóttir HM: Altered expression of HLA class I antigens in breast cancer: Association with prognosis. Int J Cancer. 2000, 89: 500-505. 10.1002/1097-0215(20001120)89:6<500::AID-IJC6>3.0.CO;2-#.
Menon AG, Morreau H, Tollenaar RA, Alphenaar E, Van Puijenbroek M, Putter H, Janssen-Van Rhijn CM, Van De Velde CJ, Fleuren GJ, Kuppen PJ: Down-regulation of HLA-A expression correlates with a better prognosis in colorectal cancer patients. Lab Invest. 2002, 82: 1725-1733.
Watson NF, Ramage JM, Madjd Z, Spendlove I, Ellis IO, Scholefield JH, Durrant LG: Immunosurveillance is active in colorectal cancer as downregulation but not complete loss of MHC class I expression correlates with a poor prognosis. Int J Cancer. 2005, 118: 6-10.
Giacomini P, Giorda E, Fraioli R, Nicotra MR, Vitale N, Setini A, Delfino L, Morabito A, Benevolo M, Venturo I, Mottolese M, Ferrara GB, Natali PG: Low prevalence of selective human leukocyte antigen (HLA)-A and HLA-B epitope losses in early-passage tumor cell lines. Cancer Res. 1999, 59: 2657-2667.
Giorda E, Sibilio L, Martayan A, Moretti S, Venturo I, Mottolese M, Ferrara GB, Cappellacci S, Eibenschutz L, Catricalà C, Grammatico P, Giacomini P: The antigen processing machinery of Human Leukocyte Antigens: linked patterns of gene expression in neoplastic cells. Cancer Res. 2003, 63: 4119-4127.
Fruci D, Ferracuti S, Limongi MZ, Cunsolo V, Giorda E, Fraioli R, Sibilio L, Carroll O, Hattori A, van Endert PM, Giacomini P: Expression of endoplasmic reticulum aminopeptidases in EBV-B cell lines from healthy donors and in leukemia/lymphoma, carcinoma, and melanoma cell lines. J Immunol. 2006, 176: 4869-4879.
Benevolo M, Mottolese M, Piperno G, Sperduti I, Cione A, Sibilio L, Martayan A, Donnorso RP, Cosimelli M, Giacomini P: HLA-A, -B, -C expression in colon carcinoma mimics that of the normal colonic mucosa and is prognostically relevant. Am J Surg Pathol. 2007, 31: 76-84. 10.1097/01.pas.0000213343.55605.b9.
Lo Monaco E, Sibilio L, Melucci E, Tremante E, Suchànek M, Horejsi V, Martayan A, Giacomini P: HLA-E: strong association with β2m and surface expression in the absence of HLA class I signal sequence-derived peptides. J Immunol. 2008, 181: 5442-5450.
Lo Monaco E, Tremante E, Cerboni C, Melucci E, Sibilio L, Zingoni A, Nicotra MR, Natali PG, Giacomini P: Human Leukocyte Antigen E contributes to protect tumor cells from lysis by natural killer cells. Neoplasia. 2011, 13: 822-830.
TNM classification of malignant tumors. 2002, Hoboken, NJ: John Wiley & Sons, 6
Stam NJ, Vroom TM, Peters PJ, Pastoors EB, Ploegh HL: HLA-A and HLA-B-specific monoclonal antibodies reactive with free heavy chains in Western blots, in formalin-fixed, paraffin-embedded tissue sections and in cryoimmuno-electron microscopy. Int Immunol. 1990, 2: 113-125. 10.1093/intimm/2.2.113.
Seitz C, Uchanska-Ziegler B, Zank A, Ziegler A: The monoclonal antibody HCA2 recognises a broadly shared epitope on selected classical as well as several non-classical HLA class I molecules. Mol Immunol. 1998, 35: 819-827. 10.1016/S0161-5890(98)00077-7.
Setini A, Beretta A, De Santis C, Meneveri R, Martayan A, Mazzilli MC, Appella E, Siccardi AG, Natali PG, Giacomini P: Distinctive features of the α1 domain α helix of HLA-C heavy chains free of β2-microglobulin. Hum Immunol. 1996, 46: 69-81. 10.1016/0198-8859(96)00011-0.
Giacomini P, Beretta A, Nicotra MR, Ciccarelli G, Martayan A, Cerboni C, Lopalco L, Bini D, Delfino L, Ferrara GB, Siccardi AG, Natali PG: HLA-C heavy chains free of β2m: distribution in normal tissues and neoplastic lesions of non-lymphoid origin and IFN-γ responsiveness. Tissue Antigens. 1997, 50: 555-566. 10.1111/j.1399-0039.1997.tb02913.x.
Braud VM, Jones EY, McMichael A: The human major histocompatibility complex class Ib molecule HLA-E binds signal sequence-derived peptides with primary anchor residues at positions 2 and 9. Eur J Immunol. 1997, 27: 1164-1169. 10.1002/eji.1830270517.
Borrego F, Ulbrecht M, Weiss EH, Coligan JE, Brooks AG: Recognition of human histocompatibility leukocyte antigen (HLA)-E complexed with HLA class I signal sequence-derived peptides by CD94/NKG2 confers protection from natural killer cell-mediated lysis. J Exp Med. 1998, 187: 813-818. 10.1084/jem.187.5.813.
Ulbrecht M, Kellermann J, Johnson JP, Weiss EH: Impaired intracellular transport and cell surface expression of nonpolymorphic HLA-E: evidence for inefficient peptide binding. J Exp Med. 1992, 176: 1083-1090. 10.1084/jem.176.4.1083.
Brooks AG, Borrego F, Posch PE, Patamawenu A, Scorzelli CJ, Ulbrecht M, Weiss EH, Coligan JE: Specific recognition of HLA-E, but not classical, HLA class I molecules by soluble CD94/NKG2A and NK cells. J Immunol. 1999, 162: 305-313.
Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, Lagorce-Pages C, Tosolini M, Camus M, Berger A, Wind P, Zinzindohoue F, Bruneval P, Cugnenc PH, Trajanoski Z, Fridman WH, Pages F: Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science. 2006, 313: 1960-1964. 10.1126/science.1129139.
Ohtani H: Focus on TILs: prognostic significance of tumor infiltrating lymphocytes in human colorectal cancer. Cancer Immun. 2007, 7: 4-12.
Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD: Cancer immunoediting: from immunosurveillance to tumor escape. Nat Immunol. 2002, 3: 991-998. 10.1038/ni1102-991.
Housseau F, Bright RK, Simonis T, Nishimura MI, Topalian SL: Recognition of a shared human prostate cancer-associated antigen by nonclassical MHC-restricted CD8+ T cells. J Immunol. 1999, 163: 6330-6337.
Levy EM, Bianchini M, Von Euw EM, Barrio MM, Bravo AI, Furman D, Domenichini E, Macagno C, Pinsky V, Zucchini C, Valvassori L, Mordoh J: Human leukocyte antigen-E protein is overexpressed in primary human colorectal cancer. Int J Oncol. 2008, 32: 633-641.
de Kruijf EM, Sajet A, van Nes JG, Natanov R, Putter H, Smit VT, Liefers GJ, van den Elsen PJ, van de Velde CJ, Kuppen PJ: HLA-E and HLA-G Expression in Classical HLA Class I-Negative Tumors Is of Prognostic Value for Clinical Outcome of Early Breast Cancer Patients. J Immunol. 2010, 185: 7452-7459. 10.4049/jimmunol.1002629.
John T, Black MA, Toro TT, Leader D, Gedye CA, Davis ID, Guilford PJ, Cebon JS: Predicting clinical outcome through molecular profiling in stage III melanoma. Clin Cancer Res. 2008, 14: 5173-5180. 10.1158/1078-0432.CCR-07-4170.
Mandruzzato S, Callegaro A, Turcatel G, Francescato S, Montesco MC, Chiarion-Sileni V, Mocellin S, Rossi CR, Bicciato S, Wang E, Marincola FM, Zanovello P: A gene expression signature associated with survival in metastatic melanoma. J Transl Med. 2006, 4: 50-10.1186/1479-5876-4-50.
Kren L, Slaby O, Muckova K, Lzicarova E, Sova M, Vybihal V, Svoboda T, Fadrus P, Lakomy R, Vanhara P, Krenova Z, Sterba J, Smrcka M, Michalek J: Expression of immune-modulatory molecules HLA-G and HLA-E by tumor cells in glioblastomas: An unexpected prognostic significance?. Neuropathology. 2011, 31: 129-134. 10.1111/j.1440-1789.2010.01149.x.
Ye SR, Yang H, Li K, Dong DD, Lin XM, Yie SM: Human leukocyte antigen G expression: as a significant prognostic indicator for patients with colorectal cancer. Mod Pathol. 2007, 20: 375-383. 10.1038/modpathol.3800751.
Bossard C, Bezieau S, Matysiak-Budnik T, Volteau C, Laboisse CL, Jotereau F, Mosnier JF: HLA-E/beta2 microglobulin over-expression in colorectal cancer is associated with recruitment of inhibitory immune cells and tumor progression. Int J Cancer. 2011
Koebel CM, Vermi W, Swann JB, Zerafa N, Rodig SJ, Old LJ, Smyth MJ, Schreiber RD: Adaptive immunity maintains occult cancer in an equilibrium state. Nature. 2007, 450: 903-907. 10.1038/nature06309.
Cifaldi L, Lo Monaco E, Forloni M, Giorda E, Lorenzi S, Petrini S, Tremante E, Pende D, Locatelli F, Giacomini P, Fruci D: NK cells efficiently reject lymphoma silenced for the endoplasmic reticulum aminopeptidase associated with antigen processing. Cancer Res. 2011
Bicknell DC, Rowan A, Bodmer WF: β2-microglobulin mutations: a study of established colorectal cell lines and fresh tumors. ProcNatlAcadSci. 1994, 91: 4751-4755.
Acknowledgements
Supported by funds from the Italian Ministry of Health agreement 2007 (to MB and PG) and by Associazione Italiana Ricerca sul Cancro (AIRC IG to PG). Dr. Hidde Ploegh (Whitehead Institute for Biomedical Research, Cambridge, MA) and Dr. Antonio Siccardi (S. Raffaele Institute, Milan, Italy) are gratefully acknowledged for generously providing HCA2, HC-10, and L31. Rocco Fraioli, Maria Vincenza Sarcone, and Paula Franke are gratefully acknowledged for skillful technical assistance, secretarial support, and linguistic revision, respectively.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
MC provided surgical specimens and supervised patient follow-up. MM and MC organized the tissue bank and the clinical pathological database. MGD reviewed and staged pathological specimens. MB, MM, CE and FR performed and evaluated immunohistochemistry. ET and ELM prepared the antibodies and performed flow cytometry and molecular HLA typing. IS participated in the design of the study and performed statistical analysis. MB, MM, MC and PG conceived, designed, and coordinated the study. MB and PG wrote the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Benevolo, M., Mottolese, M., Tremante, E. et al. High expression of HLA-E in colorectal carcinoma is associated with a favorable prognosis. J Transl Med 9, 184 (2011). https://doi.org/10.1186/1479-5876-9-184
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1479-5876-9-184