
Abstract
Members of the NF-kB transcription factor family are differentially expressed in the B cell lineage. Disruption of individual or two NF-kB subunits exhibits distinct defects in B lymphocyte development, activation, and survival. However, the role each NF-kB plays during B cell development has been obscured by molecular compensation. To address this issue, a trans-dominant form of IkBα was transduced into bone marrow cells to act as a pan-inhibitor of NF-kB using a retroviral system. While the development of T-lymphocytes and myeloid cell lineages was not grossly affected by the transduced IkBα gene, a significant reduction in the number and percentage of B lineage cells was apparent in IkBα transduced chimeric mice. IkBα expression decreased the percentage of pre-B and immature B cell subsets in the bone marrow and further impaired the development of follicular mature B cells and marginal zone B cells in the periphery. Introduction of the Bcl-X transgene completely restored the pre-B and immature B cell pool in the bone marrow. However, despite a significant improvement of overall viability of the B cell lineage, Bcl-X expression was insufficient to overcome the maturation block resulting from NF-kB inhibition. Together, our study suggests that NF-kB activity is required for two distinct checkpoints during B cell development: one is for pre-B/immature B cell viability, the other is to provide both survival and maturation signals to ensure the proper development of follicular mature B cells.
Similar content being viewed by others


Introduction
Glucocorticoid hormones were the first agents to be used as immunosuppressives, now more than 50 years ago. Since their introduction they have been the cornerstones used in most immunosuppressive treatment regimens for the prevention of organ transplant rejection, and the suppression of inflammation due to allergy and autoimmunity. Nowell first reported an explanation for the immunosuppressive action of glucocorticoids in 1961, when he found that they markedly inhibited lymphocyte proliferation in response to mitogenic lectins [1]. This was traced to an efficient inhibition of the production of interleukin 2 (then termed T Cell Growth Factor) by mitogen or antigen activated T cells almost 20 years later [2].
A major advance in understanding of the molecular basis of the effect of glucocorticoids on the expression of IL2 and other cytokine genes was reported by several groups working independently about 10 years ago, when it was found that glucocorticoid receptors physically interacted with NF-kB and as well induced the expression of the Inhibitor of NF-kB (IkB) [3–6]. Glucocorticoids have also been shown to inhibit other transcription factors, including AP-1 and NFAT [7]. Because there is an overall NF-kB suppression of transcription, the effects of glucocorticoids are protean and dramatic on many tissues, and thus their use as immunosuppressive agents is limited.
In an attempt to identify the roles played by the various members of the NF-kB family in the development and function of the immune system, we and others have explored the effects of the targeted disruption of the individual members of the NF-kB family. Our efforts have focused on c-Rel, the only member of the NF-kB family restricted to expression in the hematopoietic system. Mice lacking the c-Rel gene were found to have apparently normal lymphoid development, but there are profound defects in the antigen-induced proliferation, survival, and cytokine production by T cells and B cells [8]. By comparison, other members of the NF-kB family were found to play significant roles in the development or maturation of lymphocytes and hematopoietic cells. Because glucocorticoids inhibit the activation of all members of the NF-kB family, we sought to mimic the effects of glucocorticoids by the expression of activated IkB in hematopoietic precursor cells.
B cell development proceeds with several checkpoints that ensure the proper assembly of surface antigen receptors with non-self reactivity. In the bone marrow, the pre-BCR signals the rearrangement of the light chain and subsequent differentiation into immature B cells. Immature B cells bearing antigen receptor with self-reactivity are eliminated by negative selection process both in bone marrow and periphery. In the periphery, the newly immigrated Transitional B cells (CD24-hi/IgM+/IgD-lo) can be further subdivided into T1 (CD21-, CD23-) and T2 (CD21+, CD23+) groups based on CD21 and CD23 expression [9–12], or into T1, T2, T3 groups by including the additional marker AA4 [11]. Extracellular factors such as Blys produced by myeloid cells in the spleen and lymph nodes prolong the survival of T2 and mature B cell subsets in the periphery [13–16], while the differentiation of T2 cells into follicular mature B cells may require both Blys and BCR signals [17].
It has been well established that components of pre-BCR and BCR signaling pathways are required for B cell development. For instance, deletion of Igα, Igβ or Igμ chain arrests B cell development at the pro-B stage [18–20], while conditional deletion of the BCR on mature B cells leads to rapid loss of B cells, suggesting that basal BCR activity is required for mature B cell survival [21]. Furthermore, deletion of BCR signaling molecules, such as the tyrosine kinases Syk and Btk, severely affect the maturation of follicular B cells [22].
NF-kB is one of the key transcription factors activated by Pre-BCR and BCR signals. Not surprisingly, disruption of NF-kB members has been shown to impair BCR-mediated proliferation, survival, and Ig class switching [8, 23, 24]. Five members of the NF-kB transcription factor family (p50, p52, p65, c-Rel, RelB) display unique expression patterns and functional roles in the development of hematopoietic system, as revealed by the knockout mouse studies. Each of these five members is also subject to different regulation by IkB kinase (IKK) [8, 23, 24]. The IKK(β,α,γ) complex is primarily responsible for the activation of the NF-kB members, such as p50, p65, c-Rel, via the conventional pathway. By contrast, the IKKα complex is responsible for regulating the alternative pathway via its unique function in p100 processing and generation of the p52 complex [23, 25–27].
Accordingly, individual NF-kB subsets play distinct roles in the maturation of the B cell compartment [8]. Initial studies using cell lines derived from various B cell differentiation stages demonstrated that p50/p65 is the major inducible complex in pre-B cells and non-lymphoid cells [28]. Mature B cells, however, constitutively express c-Rel in the nucleus (in addition to p50 and p65), while terminally differentiated plasma cells and LPS-differentiated B cells express all five isoforms (p52, RelB, c-Rel, p50, and p65). Studies revealed that p50/p65 is essential for B cell development as fetal liver cells derived from p50/p65 double knockout mice are blocked in development at an early precursor stage and are unable to generate B and T lymphocytes [29]. Co-transfer of wild type bone marrow cells rescues lymphoid development in p50/p65 double knockout mice, suggesting that a cell-extrinsic factor is responsible for the defect [29].
The B cell lineage of p50/p52 double knockout mice also have a developmental block, in this case occurring at the immature T1 stage (IgM++, IgD-, CD21-, CD23-) due to an inability to receive Blys (BAFF) receptor signals [26, 30, 31]. It was later demonstrated that the Blys receptor could activate the alternative NF-kB signaling pathway, by inducing p100 processing into active p52 complexes [27, 32]. Deletion of both p65 and c-Rel perturbs the repopulation of the IgM-lo/IgD-hi mature B cell population which can be rescued by the expression of a Bcl-2 transgene, suggesting roles for c-Rel and p65 in mature B cell survival [33, 34]. p50 and c-Rel appear to regulate mature lymphocyte functions rather than early stage events, since combined deletion of p50 and c-Rel impairs the proliferation and survival of mature B cells without affecting early hematopoiesis [35, 36]. Finally, the development of MZ B cells is completely absent in p50 knockout mice and significantly reduced in c-Rel(-/-) mice or p65(-/-) fetal liver reconstituted mice [37], suggesting that multiple NF-kB members are involved in MZ B cell differentiation.
While the existing data suggest that both intrinsic and extrinsic cellular defects are accountable for many of the B cell developmental impairments observed in NF-kB knockout mice, several issues remain to be defined. First, due to molecular compensation, it is still inconclusive as to whether the residual NF-kB members compensate for certain missing individual activities. For example, Pre-BCR signaling may depend on redundant NF-kB activity for differentiation into immature B cells, as suggested by the NF-kB site occupancy in the Igκ enhancer in pre-B cells [38]. Surprisingly however, none of the NF-kB double knockout mice have impaired development of early B cell precursors in the bone marrow. In addition, the peripheral B cell maturation deficit observed in p65/c-Rel double knockout chimeras has been mainly attributed to a survival defect. However, the potential role of other NF-kB members (especially p50/p52) in the differentiation process prior to maturation into follicular B cells has not been addressed.
Here, we report the use of a pan-NF-kB inhibitor, IkBα, to block all NF-kB activity in bone marrow derived precursor cells. We show that expression of a dominant active form of IkBα significantly impairs B cell development at two stages: first at the pre-B expansion stage, the second occurring at transitional B cell maturation stage in the periphery. Furthermore, NF-kB not only contributes to immature B cell survival but also is required for maturation of follicular B cells. Accordingly, by inducing the expression of IkB, glucocorticoids have a profound effect on B-cell development and function, in addition to its already well-described effect of preventing T-cell proliferation and differentiation by blocking NF-kB activation of T cell cytokine gene expression.
Materials and methods
Preparation of retrovirus and determination of virus titer
A human cDNA encoding for the IkBα without amino acids 1–36 was inserted into the MIGR1 plasmid, which contains an MSV promoter and GFP [39, 40]. The target gene and GFP gene were separated by IRES and can be expressed separately. The plasmids were cotransfected with pHIT123 and PCGP into 293T cell by using calcium phosphate method. The 293T cells were seeded the previous day to give a maximum of 70% confluence/plate on the day of transfection. Ten micrograms of each plasmid was used for each transfection. At 48 hours post transfection, the supernatant was harvested and assayed for viral titer by infection on NIH3T3 cells. The retrovirus supernatant was stored at -80°C for future use.
Generation of chimeric mice with bone marrows infected with IkBα and control viruses
Female C57BL/6 mice (8–10 weeks old) were purchased from The Jackson Labs (USA) and kept in specific pathogen-free conditions. C57BL/6 mice were injected with 5-fluorouracil (5-FU, 250 mg/kg weight) per animal for 4 days. Bone marrow cells (BMCs) were isolated from tibias and femurs of mice and suspended in DMEM with 5% heat-inactivated fetal calf serum (FCS). Red blood cells were depleted using ACK lysis buffer [41]. These BMCs were cultured in 6-well plate with a cocktail of cytokines, IL3 (6 ng/ml), IL6 (10 ng/ml) and SCF (100 ng/ml), for one day. Then the retrovirus supernatant was added into the BMCs, and cultured for an additional 4–6 days. Cells were then collected and transferred into lethally irradiated mice (850 Rad). Bone marrow chimeras were analyzed at week 2, 4, 6 post-transfer.
Antibodies
Flow cytometric analyses of surface markers on hematopietic cells were performed on FACScan instruments (Becton Dickinson). PE- or biotin-conjugated antibodies specific for B220 (PE or biotin), IgM (PE), CD23 (PE), CD21 (Biotin), Gr-1 (PE), CD11b (PE), CD4 (PE) and CD8 (PE) were obtained from Pharmingen (San Diego, CA). Thy 1.2 (biotin), and the Tricolour-conjugated streptavidin was obtained from CALTAG (Burlingame, CA).
Electrophoretic mobility shift assay (EMSA)
The nuclear and cytosolic fractionation procedure was prepared as previously described [28]. Purified B lymphocytes (1 × 107 cells per treatment) were harvested, washed once with PBS, resuspended into 100 μl of Buffer A {10 mM HEPES (pH 7.9), 1.5 mM MgCl2, 10 mM KCl and 0.5 mM DTT} plus 0.1% NP-40. Cells were lysed on ice for 2 min and checked for complete lysis under the microscope. Nuclei were spun down and the supernatant was saved as cytosolic fraction. The nuclear pellet was resuspended into 100 μl of Buffer C [20 mM HEPES (pH 7.9), 25% glycerol, 0.42 M NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM PMSF and 0.5 mM DTT], sonicated to decrease viscosity and rotated in the cold room for 30 min. Ultracentrifugation was performed to remove insoluble debris, and supernatant used as nuclear extract. Nuclear proteins were quantified by the commercial Bio-Rad protein assay, and EMSA was performed as described previously [28].
Results
Ectopic IkBα expression does not affect the development of granulocytes, monocyte, and T cell lineages
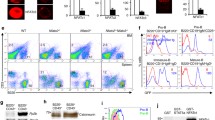
To assess the role of NF-kB during B cell development, a trans-dominant form of the NF-kB inhibitor, IkBα, was constructed into a GFP-containing retroviral vector, MIGR1 [39, 40, 42] and used for bone marrow infection and transfer experiments (Supplemental Figure 8 and Figure 9). The IkBα and control viruses were obtained by co-transfecting the expression vectors with the packaging plasmids into 293T cells. Viral titer was determined by infecting NIH3T3 cells. In a typical experiment, the transfection efficiency of MIGR1 and IkBα plasmids reached 68–76% in 293T cells (Figure 1A) with viral titers in the range of 1–5 × 107/ml as determined on 3T3 cells (Figure 1B). Bone marrow cells derived from 5-FU treated mice were then infected with retroviruses in culture for 48 hours. Roughly 60–90% infection efficiency of bone marrow cells infected with both MIGR1 and IkBα viruses was achieved prior to transfer into lethally-irradiated recipient mice (Figure 1C).

Transfection efficiency, viral titer, and infection efficiency Three plasmids (pHIT123, pCGP and MIGR1 or MIGR1-IkB) were transfected into 293T cell for 48 hours. The supernatant was collected and tested for viral titer by using NIH3T3 cell line. The viral supernatant was subsequently used for infecting bone marrow cells. Flow cytometry was performed to determine the transfection efficiency on 293T cells (A), viral titer on 3T3 cells (B), and viral infection efficiency on bone marrow cells (C) prior to transfer into lethally-irradiated recipient mice.
Recipient chimeras were next analyzed at two and four weeks post bone marrow transfer. Bone marrow and spleen cells from the recipients were stained with various hematopoietic lineage markers and analyzed by flow cytometry. Chimeric mice receiving the control MIGR1-infected bone marrows reconstituted with granulocytes and macrophages within normal range as the non-manipulated control mice. The percentage of Gr1+ and Mac1+ cells are comparable in both GFP+ or GFP- populations of the MIGR1 reconstituted recipients, suggesting that MIGR1 (and GFP expression) control virus did not affect the differentiation of these cell lineages. By contrast, chimeras receiving the IkBα-infected bone marrow cells appeared to have a slightly higher percentage of Gr1+ and Mac1+ cells in the GFP+ population than in the GFP- population in both bone marrow and spleen (data not shown). This apparent increase in granulocyte and monocyte ratio, in IkBα-expressing (GFP+) population, correlated with the dramatic decrease in B cell numbers (see below). The total cell number in bone marrow and spleens were comparable in MIGR1 and IkBα reconstituted recipients (Figure 7).
The development of the T cell compartment was also examined by staining the bone marrow, spleen, and thymus with CD4 and CD8 antibodies. As expected, bone marrow contains very few CD4 or CD8 positive cells. However, in the spleen, IkBα expression slightly decreased the CD4 and CD8 single positive T cell percentage. Repopulation of thymocytes in the thymus became visible four weeks post-transfer. Interestingly, IkBα expression in the T cell lineage did not grossly affect the percentage of CD4/CD8 double positive, double negative, or single positive population despite high viral transduction efficiency (greater than 84% cells were GFP+) (data not shown). Because these results are reproducible among several independent experiments, our data suggest that NF-kB does not play a major role in T cell development, contrary to its observed effects on B cell development (see below).
Ectopic IkBα expression severely blocks B cell development in the bone marrow
B cell development was next examined using various B cell markers. Chimeras reconstituted with the IkBα-expressing bone marrow had dramatically reduced numbers of B220+ cells in the bone marrow and spleen as compared to those receiving MIGR1-infected control BM cells (Figure 2A). Whereas we observed a reconstitution level of 83% in BM and 86% in spleen of B220+ cells within the GFP+ population of MIGR1 chimeras, there was only 55% in BM and 21% in spleen of B220+ cells in the GFP+ population of IkBα-transduced chimeras (Figure 2A). These results suggest that ectopic IkBα expression affects early B cell development beginning in the bone marrow. Moreover, the lower percentage of B cells in the spleen than in the bone marrow suggests a further deficiency as immature B cells emerge into the periphery (Figure 7).

IkBα overexpression leads to a reduction in the Pre-B/immature B cell compartment in the bone marrow Bone marrow cells were isolated from the MIGR1 control and IkBα-transduced chimeras at two and four weeks post-BM transplantation. Cells were stained with biotin-conjugated B220 (A) or co-stained with PE-conjugated CD43 and biotin-conjugated B220 antibodies (B) and analyzed by flow cytometry. For (B), B220+ and CD43+ populations were assessed based on the GFP+ population. The data is a representative of five experiments.
To trace the effect of IkBα on B cell progenitors, bone marrow cells were co-stained with B220 and CD43 antibodies to determine the percentage of cells at pro-B and pre-B stages. As shown in Figure 2B, the percentage of B220+/CD43- pre-B/immature B cells in the IkBα transduced chimeras is significantly lower than that of the MIGR1 control chimeras (47% vs. 70%). By comparison, the percentage of the B220+/CD43+ pro-B cells is comparable in the IkBα and MIGR1 transduced chimeras (16% vs. 15%). These results indicate that IkBα expression interferes with the pro-B to pre-B transition, which coincides with the requirement of the pre-BCR signals for further differentiation into immature B cells.
IkBα expression also arrests B cell maturation in the periphery
In the IkBα transduced chimeras, decreased B cell numbers are also observed in the periphery, and to much greater extent than in the bone marrow (21% vs 55%, Figure 2A), indicating additional deficiency occurring after immature B cells have emigrated from the bone marrow. To further analyze this defect, we examined B cell subsets in the spleen for the presence of different B cell subsets.
Purified splenic B cells (>95% B220+) derived from both MIGR1 and IkBα transduced chimeras were co-stained with CD21 and CD23 antibodies. GFP+ populations were further gated into follicular (CD21+/CD23+), marginal zone (CD21+/CD23-) and immature (CD21-/CD23-) B cell subsets. As shown in Figure 3A, follicular B cells are significantly reduced in the IkBα transduced chimeras as compared to the MIGR1 controls (34% in the IkBα group vs. 54% in the MIGR1 group), correlating with increased numbers of immature B cells (58% of CD21-, CD23-) occurring in the IkBα-transduced chimera than in the control group (36%). In addition, IkBα chimeras had slightly reduced levels of marginal zone B cells versus controls (2% vs. 7%). Together, these results demonstrate that IkBα expression affects B cell maturation into follicular and marginal zone B cells.

IkBα overexpression impairs the maturation of follicular and marginal zone B cells in the spleen Purified splenic B cells (>95% B220+) were isolated from the MIGR1 control and IkBα-transduced chimeras two and four weeks after bone marrow transplantation. (A) Cells were co-stained with biotin-conjugated CD21 and PE-conjugated CD23 antibodies and analyzed by flow cytometry. After gating on GFP+ population, the percentage of follicular B cells (CD21+/CD23+), marginal zone B cells (CD21+/CD23-), and transitional B cells (CD21-/CD23-) were enumerated. (B) Cells were co-stained with biotin-conjugated CD24 and PE-conjugated IgM antibodies and analyzed by flow cytometry. After gating on GFP+ population, the percentage of Transitional immature B cells (CD24-hi, IgM+) and mature B cells (CD24-lo, IgM+) were enumerated. (C) Cells were co-stained with biotin-conjugated CD21 and PE-conjugated IgM antibodies and analyzed by flow cytometry. After gating on the GFP+ population, the percentage of Transitional 1 B cells (CD21-, IgM+) and more mature B cells (CD21+, IgM+, containing T2, FB, and MZ B cells) were enumerated.
The developmental block at the immature B cell stage imposed by IkBα expression was further supported by staining purified splenic B cells with additional markers: CD24, CD21, and IgM. Immature B cells express high CD24 levels compared to low levels expressed by the mature B cells. When the GFP+ populations were further analyzed for CD24 and IgM levels, it was evident that 60% of cells were of the immature B phenotype (CD24-hi, IgM+) in the IkBα-transduced chimeras compared to only 29% in the MIGR1 control group at four weeks post transfer (Figure 3B, bottom panels). Accordingly, the percentage of mature B cells (CD24-lo, IgM+) is lower in the IkBα group (13%) than in the MIGR1 control group (42%). At two weeks post-transfer, most B cells expressed high CD24 levels, indicative of their immature phenotype (Figure 3B, top panels).
Immature B cells can be further divided into transitional 1 (T1) and T2 types based on the CD21 and CD23 expression. Whereas the T1-B cells are negative for both markers (CD21-, CD23-), T2-B cells express both markers (CD21+, CD23+). To facilitate the identification of these B cell subsets, purified splenic B cells were co-stained with CD21 and IgM. GFP+ populations were further gated and analyzed for the CD21 and IgM expression levels. As shown in Figure 3C, there is a greater percentage of T1-B cells in the IkBα-transduced chimeras than the MIGR1 controls (31% vs. 19%) and a lower percentage of more mature (T2, follicular, and MZ) B cell populations in the IkBα group compared to control group (44% vs. 50%).
Collectively, these data demonstrate that ectopic IkBα expression affects B cell development at two checkpoints: it impairs the pro-B to pre-B transition in the bone marrow, and causes a developmental arrest of B cell maturation in the periphery with reduced mature follicular B cells and marginal B cells.
Bcl-X transgene rescues the pre-B deficiency imposed by IkBα expression
The majority of the NF-kB-mediated biological activities have been attributed to the regulation of cell survival genes by NF-kB proteins [43–48]. In addition to providing survival signals, certain NF-kB members have also been shown to participate in cell cycle, differentiation, and effector functions in immune cells [8, 49–51]. For example, c-Rel deletion leads to impaired cell cycle progression that is independent of cell survival defect in B-lymphocytes [49, 51]. RelB and p50/p65 are required for dendritic cell differentiation [52–55], whereas p50/c-Rel deletion impairs only mature dendritic cell cytokine production and costimulation function without affecting differentiation [55, 56].
It is therefore important to discern whether NF-kB contributes to B cell development through maintaining B cell survival and/or regulating differentiation processes. To address this issue, we utilized Bcl-X transgenic mice that preferentially express Bcl-X in the B cell lineage [47, 57, 58]. Bone marrow cells derived from Bcl-X transgenic mice (as well as non-transgenic control mice) were infected with IkBα or MIGR1 retroviruses as described above, and transferred into irradiated recipients. Bone marrow and spleen of the chimeric mice were then analyzed two or four weeks after reconstitution. Consistent with the above findings, IkBα transduced chimeras had a reduced percentage of B220+ cells in the bone marrow. By gating on GFP+ populations, only 51% B220+ cells were detected in the IkBα-expressing BM cells as compared to 71% B220+ cells in the MIGR1 reconstituted chimeras (Figure 4A). Interestingly however, the Bcl-X transgene expression completely restored B cell percentage in the IkBα-transduced chimeras (Figure 7). B220+ cells within the GFP+ populations were 57% and 64% in the MIGR1 and the IkBα-chimeras, respectively. A similar trend was observed for chimeras analyzed two weeks post transfer (Figure 4A, four panels on the left).

Bcl-X completely rescues the Pro-B to Pre-B transition in IkBα-transduced chimeras Bone marrows derived from Bcl-X transgenic (WTtg) and non-transgenic (WT) mice were infected with MIGR1 control and IkBα-viruses in vitro and transferred to lethally-irradiated recipients as described in Materials and Methods. Two and four weeks after bone marrow transplantation, reconstituted bone marrow cells were isolated from the MIGR1 control and IkBα-transduced chimeras. Cells were stained with biotin-conjugated B220 (A) or co-stained with PE-conjugated CD43 and biotin-conjugated B220 antibodies (B) followed by flow cytometry. For (B), the percentage of cells expressing B220 and CD43 was determined based on the GFP+ population.
Further analysis of the B cell precursors using CD43 and B220 double staining revealed that, among the GFP+ populations, IkBα overexpression severely reduced the percentage of Pre-B/immature B cells (16%) compared to MIGR1 control (55%) at four weeks post reconstitution (Figure 4B, four panels on the right). Intriguingly, the Bcl-X transgene completely restored the Pre-B/immature B cell population in IkBα-transduced chimeras (40%) to MIRG1 control levels (39%). These results thus suggested that the IkBα-transduced Pre-B/immature B cell developmental block resulted from diminished cell viability.
Bcl-X transgene rescues the viability of immature B cells but fails to override the maturation block imposed by IkBα expression
The effect of Bcl-X transgene on B cell maturation in periphery was further analyzed in the spleens of chimeric mice at four weeks post reconstitution. Consistent with previous results (Figure 2A), there was a significant decline in the B220+ cell percentage within the splenic GFP+ population of the IkBα-transduced chimeras (45%) compared to MIGR1 transduced chimeras (85%) (Figure 5A). Bcl-X transgene was able to restore majority of B220+ cells in the spleen of IkBα transduced chimeras (78% as compared to 89% in MIGR1 samples), suggesting that IkBα induced loss of peripheral B cells occurs via apoptosis. However, when B cell subsets were further analyzed, it was evident that B cell maturation was not fully restored. Staining of the purified splenic B cells (>95% B220+) with CD21 and CD23 (and further gating for GFP+ population) revealed that follicular B cells constitute 64% and 87% of total B cells in the chimeras receiving the control wild type and in Bcl-X transgenic bone marrow, respectively (Figure 5B, two panels on the left). However, the IkBα-transduced bone marrow chimeras had reduced follicular B cells (49%) (Figure 5B). Interestingly, although the Bcl-X transgene rescued peripheral B cell numbers in the IkBα-transduced chimeras (Figure 7, data not shown), only 16% of the B cells consist of follicular mature B cells. The majority of the rescued B cells were arrested at the immature B cell stage (75%) (Figure 5B). Using additional markers including CD24, CD21, and IgM, we reached similar conclusions where we observed that a relatively high percentage (44% for WT, 38% for WTtg) of B cells exhibited immature phenotype (CD24-hi, IgM+, or CD21-, IgM+) in IkBα-transduced chimeras compared to control groups (15% for WT, 23% for WTtg) (Figures 5C and data not shown). The mature B cell percentage was significantly reduced in IkBα-transduced chimeras regardless of Bcl-X expression (17% for WT, 19% for WTtg) as compared to control samples (59% for WT, 52% for WTtg). Therefore, despite significantly improved cell viability and cell number, Bcl-X expression is insufficient to override the maturation block imposed by IkBα protein expression. These data suggest that NF-kB not only provide survival signals for immature B cells but also is required for the final maturation of follicular B cells in the periphery.

Bcl-X expression significantly improves B cell viability but is insufficient to drive the maturation of transitional B cells Bone marrow derived from Bcl-X transgenic (WTtg) and non-transgenic (WT) mice were infected with MIGR1 control and IkBα-viruses in vitro and transferred to lethally-irradiated recipients. Splenocytes were isolated from the MIGR1 control and IkBα-transduced chimeras two and four weeks after bone marrow transplantation. (A) Cells were stained with PE-conjugated B220 antibody and analyzed by flow cytometry. (B) Cells were co-stained with biotin-conjugated CD21 and PE-conjugated CD23 antibodies and analyzed by flow cytometry. After gating on GFP+ population, the percentage of follicular B cells (CD21+/CD23+), marginal zone B cells (CD21+/CD23-), and transitional B cells (CD21-/CD23-) were enumerated. (C) Cells were co-stained with biotin-conjugated CD24 and PE-conjugated IgM antibodies and analyzed by flow cytometry. After gating on GFP+ cells, the percentage of Transitional immature B cells (CD24-hi, IgM+) and mature B cells (CD24-lo, IgM+) were enumerated.
Discussion
In this study, we address the role of NF-kB in B-cell lymphopoiesis and differentiation by introducing a pan-NF-kB inhibitor with retroviral transduction and bone marrow transfer approaches. Our data demonstrate that NF-kB inhibition affects B cell differentiation at the pro-B to pre-B cell transition. An additional impairment is observed in the periphery where the survival and maturation of transitional immature B cells is affected. Interestingly, Bcl-X expression can rescue the defect at the pre-B stage and significantly increases the number of transitional B cells in the spleen. Nonetheless, these peripheral B cells remain arrested at the immature stage. This study suggests that NF-kB activity is required not only for the overall survival of B cells at all developmental stages, but also for the maturation of transitional B cells into follicular B cells in the periphery.
Numerous studies have established the contribution of pre-BCR and BCR signals in B cell development and survival. Deletions of the Igμ or Igα/β chains lead to developmental arrest at the pro-B stage, suggesting that the pre-BCR signals are essential for inducing Ig light chain gene rearrangement and differentiation into pre-B and immature B cells [18–20]. Supporting this notion is the finding that in vivo occupancy of the κB site is observed by in vivo footprinting analysis of the Igκ intronic enhancer in primary pro-B and pre-B cells [38]. This observation is consistent with our finding that IkBα expression blocks Pro-B to Pre-B cell stage transition, as NF-kB inhibition should prevent Igκ gene expression and perturb the assembly of IgM on cell surface.
Targeted disruption of the surface BCR on mature B cells results in rapid cell death, indicating that the basal BCR activity is required for maintaining the survival of resting mature B cells [21]. Further studies on downstream signaling molecules have suggested more elaborate developmental controls after the emigration of immature B cells to the periphery. Ablation of many of the signaling components (e.g. tyrosine kinases, adaptor molecules) and downstream transcription factors (e.g. NF-kB) affects the final maturation of transitional immature B cells [22]. For example, deletion of the Syk kinase completely blocks the maturation of transitional B cells into recirculating follicular B cells [59]. On the other hand, expression of Bcl-2 results in a greater accumulation of immature B cells both in the bone marrow and in spleen, however, these cells retain a the transitional B cell phenotype. These observations are remarkably similar to our findings using IkBα-transduced bone marrow chimeras, thus suggesting that Syk and NF-kB are likely to fall in the same signaling pathway.
While the exact mechanism governing the maturation of transitional T1, T2, into follicular or marginal zone B cells is still unclear, knockout studies have identified genes that are involved in these processes. For example, Blys(-/-) mice have normal levels of T1 cells, but lack T2, marginal zone, and follicular mature B cells in the periphery [60, 61]. This phenotype is mimicked by that of the NF-kB p50/p52 double knockout mice. In fact, Blys has been shown to induce p100 processing and the generation of active p52 complexes, further supporting the role of these NF-kB members (p50, p52) in T1 to T2 transition [26, 27].
A few studies have also shown that the late-stage T2 cells may require both Blys and BCR signals for differentiation into follicular mature B cells [17]. This is consistent with the observation that many of the gene-targeted deletions of BCR signaling components affect the differentiation into the IgM-lo/IgD-hi follicular mature B cell stage, including Lyn, Btk, PLC-γ2, BCAP, vav, PI3K (p85, p110δ), and PKCβ [22, 62]. Coincidently, these signaling molecules have been shown to regulate conventional NF-kB activation. Thus, it is of interest to note that the final maturation from late-stage T2 cells to follicular B cells requires BCR-dependent NF-kB mediated differentiation signaling, as survival signal provided by Blys or Bcl-X alone is insufficient to complete the maturation processes. This notion is further supported by our data showing that Bcl-X restores the transitional B cell number, but is unable to induce the maturation of these cells into follicular B cells. Furthermore, studies on NF-kB p65/c-Rel and p50/c-Rel double knockout mice are consistent with the view that these conventional NF-kB complexes are required for the generation of follicular mature B cells [33, 35].
Based on previous NF-kB knockout mouse studies and the findings from this report, we propose the following model to describe the roles of distinct NF-kB members in B cell differentiation (see Figure 6, Model). In the bone marrow, the Pre-BCR signals are required for Ig light chain expression and the assembly of IgM, which marks the differentiation of immature B cells. The p50 and p65 NF-kB members are particularly important in mediating the Pre-BCR signaling and may participate in the regulation of Igκ gene expression. In the periphery, Blys-mediated survival signals of transitional B cells absolutely require the presence of p52. Blys signals may eventually cooperate with BCR signals (via p65, c-Rel) in driving maturation into the follicular B cells. Mature B cells critically depend on c-Rel complexes to regulate cell survival and cell cycle programs upon recognition of antigens by BCR on the cell surface.

Model for the roles of NF-kB members in B cell development Distinct NF-kB members are employed at different stages of B cell development. Previous studies indicate that p50 and p65 are likely involved in the Pre-BCR-mediated differentiation process, including the Ig light chain expression. C-Rel expression becomes detectable following Pre-B to immature B cell differentiation [38]. Meanwhile, p50 and p52 members are critical for the survival and differentiation of Transitional B cells, particularly in response to Blys signals. The final maturation of late-stage T2 B cells into follicular B cells also requires BCR signals that induce p65 and c-Rel activity. In mature B cells, the c-Rel complexes are especially important for BCR-mediated survival and cell cycle progression. All NF-kB members have been shown to participate in Ig class switching.
It is somewhat surprising to us that the IkBα expression does not grossly affect thymocyte development. Our finding, however, is consistent with several reports using IkBα transgenic mice. According to one study, double positive (DP) CD4+ CD8+ T cell development is normal in IkBα transgenic mice but single positive (SP) populations are significantly reduced [63], whereas in another study only CD8+ thymocytes were affected [64]. In both studies, peripheral CD4 and CD8 T cell numbers were severely diminished. Similar results have been obtained in p65-c-Rel double knockout that deletion of both c-Rel and p65 leads to only decreased peripheral CD4+ and CD8+ T cell numbers without affecting early T cell development [34, 65]. Moreover, a detailed analysis of IkBα transgenic double negative (DN) thymocytes has revealed a specific reduction in the number of DN-IV stage thymocytes. The subtle differences in these studies could be attributed to differing levels of IkBα expression. In addition, Lck mediated expression of IkBα gene may be more specifically targeted to T cell precursors, whereas these precursors may be under-represented in the bone marrow cultures as used in this report.

Total and GFP+ cell numbers in the bone marrow and spleens of virus-transduced bone marrow chimeras. Total cell number and percentage of GFP+ cells in the bone marrow and spleen of individual chimera was determined. WTtg-MIGR1 and WTtg-IkB represent the Bcl-X transgenic mice that were transduced with MIGR1 and IkBa virus, respectively. Statistical analysis was performed on combined data from all chimeras used in several independent experiments. The GFP+ cell number was calculated by multiplying the total cell number with the GFP+ percentage. The number in the parentheses represents the percentage of cell numbers in the IkBa group over the cell number in the MIGR1 group (100%). ND: Not determined.

MIGR1-IkBa construct. The cDNA encoding human IkBa (from aas 38-280) was cloned into the Bgl II and EcoRI sites of the MIGR1 retroviral plasmid [36, 37]. The expression of IkBa was confirmed by western blot analysis of transfected 293 cell line (data not shown).

Inhibition of NF-kB binding activity in splenic B cells derived from MIGR1 and IkBa transduced chimeras. Splenic B cells were purified from MIGR1 control or IkBa virus-transduced bone marrow chimeras and treated with medium, anti-CD40, or anti-IgM for 4 hours. Nuclear extracts were prepared from the cells and analyzed by EMSA using 32P-labeled IgkB oligonucleotides as described in Materials and Methods. The transduction efficiency for MIGR1 control or IkBa virus-transduced cells used for this particular experiment is 29% and 15%, respectively. IkBa transduced B cells exhibit decreased NF-kB DNA binding activity, confirming that the IkBa described in the paper is a biological active inhibitor.
Our findings detailed in this report point towards the rational for the development of new pharmaceutical agents that mimic the effect of glucocorticoids on the activation of IkB that could function to be effective immunosuppressive drugs. Conversely, agents that target the suppression or removal of IkB in lymphoid cells could be developed as immunopotentiating compounds.
References
Nowell PC: Inhibition of human leukocyte mitosis by prednisolone in vitro. Cancer Res. 1961, 21: 1518-1521.
Gillis S, Crabtree GR, Smith KA: Glucocorticoid-induced inhibition of T cell growth factor production. I. The effect on mitogen-induced lymphocyte proliferation. J Immunol. 1979, 123: 1624-1631.
Scheinman RI, Cogswell PC, Lofquist AK, Baldwin A. S., Jr.: Role of transcriptional activation of I kappa B alpha in mediation of immunosuppression by glucocorticoids. Science. 1995, 270: 283-286.
Ray A, Prefontaine KE: Physical association and functional antagonism between the p65 subunit of transcription factor NF-kappa B and the glucocorticoid receptor. Proc Natl Acad Sci U S A. 1994, 91: 752-756.
Scheinman RI, Gualberto A, Jewell CM, Cidlowski JA, Baldwin A. S., Jr.: Characterization of mechanisms involved in transrepression of NF-kappa B by activated glucocorticoid receptors. Mol Cell Biol. 1995, 15: 943-953.
Auphan N, DiDonato JA, Rosette C, Helmberg A, Karin M: Immunosuppression by glucocorticoids: inhibition of NF-kappa B activity through induction of I kappa B synthesis. Science. 1995, 270: 286-290.
Adcock IM: Glucocorticoid-regulated transcription factors. Pulm Pharmacol Ther. 2001, 14: 211-219. 10.1006/pupt.2001.0283.
Liou HC, Hsia CY: Distinctions between c-Rel and other NF-kappaB proteins in immunity and disease. Bioessays. 2003, 25: 767-780. 10.1002/bies.10306.
Loder F, Mutschler B, Ray RJ, Paige CJ, Sideras P, Torres R, Lamers MC, Carsetti R: B cell development in the spleen takes place in discrete steps and is determined by the quality of B cell receptor-derived signals. J Exp Med. 1999, 190: 75-89. 10.1084/jem.190.1.75.
Su TT, Rawlings DJ: Transitional B lymphocyte subsets operate as distinct checkpoints in murine splenic B cell development. J Immunol. 2002, 168: 2101-2110.
Allman D, Lindsley RC, DeMuth W, Rudd K, Shinton SA, Hardy RR: Resolution of three nonproliferative immature splenic B cell subsets reveals multiple selection points during peripheral B cell maturation. J Immunol. 2001, 167: 6834-6840.
Chung JB, Sater RA, Fields ML, Erikson J, Monroe JG: CD23 defines two distinct subsets of immature B cells which differ in their responses to T cell help signals. Int Immunol. 2002, 14: 157-166. 10.1093/intimm/14.2.157.
Mackay F, Browning JL: BAFF: a fundamental survival factor for B cells. Nat Rev Immunol. 2002, 2: 465-475. 10.1038/nri844.
Do RK, Hatada E, Lee H, Tourigny MR, Hilbert D, Chen-Kiang S: Attenuation of apoptosis underlies B lymphocyte stimulator enhancement of humoral immune response. J Exp Med. 2000, 192: 953-964. 10.1084/jem.192.7.953.
Hsu BL, Harless SM, Lindsley RC, Hilbert DM, Cancro MP: Cutting edge: BLyS enables survival of transitional and mature B cells through distinct mediators. J Immunol. 2002, 168: 5993-5996.
Hatada EN, Do RK, Orlofsky A, Liou HC, Prystowsky M, MacLennan IC, Caamano J, Chen-Kiang S: NF-kappaB1 p50 is required for BLyS attenuation of apoptosis but dispensable for processing of NF-kappaB2 p100 to p52 in quiescent mature B cells. J Immunol. 2003, 171: 761-768.
Batten M, Groom J, Cachero TG, Qian F, Schneider P, Tschopp J, Browning JL, Mackay F: BAFF mediates survival of peripheral immature B lymphocytes. J Exp Med. 2000, 192: 1453-1466. 10.1084/jem.192.10.1453.
Kitamura D, Roes J, Kuhn R, Rajewsky K: A B cell-deficient mouse by targeted disruption of the membrane exon of the immunoglobulin mu chain gene. Nature. 1991, 350: 423-426. 10.1038/350423a0.
Reichlin A, Hu Y, Meffre E, Nagaoka H, Gong S, Kraus M, Rajewsky K, Nussenzweig MC: B cell development is arrested at the immature B cell stage in mice carrying a mutation in the cytoplasmic domain of immunoglobulin beta. J Exp Med. 2001, 193: 13-23. 10.1084/jem.193.1.13.
Meffre E, Nussenzweig MC: Deletion of immunoglobulin beta in developing B cells leads to cell death. Proc Natl Acad Sci U S A. 2002, 99: 11334-11339. 10.1073/pnas.172369999.
Lam KP, Kuhn R, Rajewsky K: In vivo ablation of surface immunoglobulin on mature B cells by inducible gene targeting results in rapid cell death. Cell. 1997, 90: 1073-1083.
Niiro H, Clark EA: Regulation of B-cell fate by antigen-receptor signals. Nat Rev Immunol. 2002, 2: 945-956. 10.1038/nri955.
Ghosh S, Karin M: Missing pieces in the NF-kappaB puzzle. Cell. 2002, 109 Suppl: S81-96. 10.1016/S0092-8674(02)00703-1.
Karin M, Lin A: NF-kappaB at the crossroads of life and death. 2002
Karin M, Delhase M: The I kappa B kinase (IKK) and NF-kappa B: key elements of proinflammatory signalling. Semin Immunol. 2000, 12: 85-98. 10.1006/smim.2000.0210.
Claudio E, Brown K, Park S, Wang H, Siebenlist U: BAFF-induced NEMO-independent processing of NF-kappaB2 in maturing B cells. Nat Immunol. 2002, 3: 958-965. 10.1038/ni842.
Kayagaki N, Yan M, Seshasayee D, Wang H, Lee W, French DM, Grewal IS, Cochran AG, Gordon NC, Yin J, Starovasnik MA, Dixit VM: BAFF/BLyS receptor 3 binds the B cell survival factor BAFF ligand through a discrete surface loop and promotes processing of NF-kappaB2. Immunity. 2002, 17: 515-524. 10.1016/S1074-7613(02)00425-9.
Liou HC, Sha WC, Scott ML, Baltimore D: Sequential induction of NF-kappa B/Rel family proteins during B-cell terminal differentiation. Mol Cell Biol. 1994, 14: 5349-5359.
Horwitz BH, Scott ML, Cherry SR, Bronson RT, Baltimore D: Failure of lymphopoiesis after adoptive transfer of NF-kappaB-deficient fetal liver cells. Immunity. 1997, 6: 765-772. 10.1016/S1074-7613(00)80451-3.
Franzoso G, Carlson L, Xing L, Poljak L, Shores EW, Brown KD, Leonardi A, Tran T, Boyce BF, Siebenlist U: Requirement for NF-kappaB in osteoclast and B-cell development. Genes Dev. 1997, 11: 3482-3496.
Franzoso G, Carlson L, Poljak L, Shores EW, Epstein S, Leonardi A, Grinberg A, Tran T, Scharton-Kersten T, Anver M, Love P, Brown K, Siebenlist U: Mice deficient in nuclear factor (NF)-kappa B/p52 present with defects in humoral responses, germinal center reactions, and splenic microarchitecture. J Exp Med. 1998, 187: 147-159. 10.1084/jem.187.2.147.
Claudio E, Brown K, Park S, Wang H, Siebenlist U: BAFF-induced NEMO-independent processing of NF-kappa B2 in maturing B cells. Nat Immunol. 2002, 3: 958-965. 10.1038/ni842.
Grossmann M, Metcalf D, Merryfull J, Beg A, Baltimore D, Gerondakis S: The combined absence of the transcription factors Rel and RelA leads to multiple hemopoietic cell defects. Proc Natl Acad Sci U S A. 1999, 96: 11848-11853. 10.1073/pnas.96.21.11848.
Grossmann M, O'Reilly LA, Gugasyan R, Strasser A, Adams JM, Gerondakis S: The anti-apoptotic activities of Rel and RelA required during B-cell maturation involve the regulation of Bcl-2 expression. Embo J. 2000, 19: 6351-6360. 10.1093/emboj/19.23.6351.
Pohl T, Gugasyan R, Grumont RJ, Strasser A, Metcalf D, Tarlinton D, Sha W, Baltimore D, Gerondakis S: The combined absence of NF-kappa B1 and c-Rel reveals that overlapping roles for these transcription factors in the B cell lineage are restricted to the activation and function of mature cells. Proc Natl Acad Sci U S A. 2002, 99: 4514-4519. 10.1073/pnas.072071599.
Grumont RJ, Rourke IJ, O'Reilly LA, Strasser A, Miyake K, Sha W, Gerondakis S: B lymphocytes differentially use the Rel and nuclear factor kappaB1 (NF-kappaB1) transcription factors to regulate cell cycle progression and apoptosis in quiescent and mitogen-activated cells. J Exp Med. 1998, 187: 663-674. 10.1084/jem.187.5.663.
Cariappa A, Liou HC, Horwitz BH, Pillai S: Nuclear factor kappa B is required for the development of marginal zone B lymphocytes. J Exp Med. 2000, 192: 1175-1182. 10.1084/jem.192.8.1175.
Shaffer AL, Peng A, Schlissel MS: In vivo occupancy of the kappa light chain enhancers in primary pro- and pre-B cells: a model for kappa locus activation. Immunity. 1997, 6: 131-143.
Pui JC, Allman D, Xu L, DeRocco S, Karnell FG, Bakkour S, Lee JY, Kadesch T, Hardy RR, Aster JC, Pear WS: Notch1 expression in early lymphopoiesis influences B versus T lineage determination. Immunity. 1999, 11: 299-308. 10.1016/S1074-7613(00)80105-3.
Izon DJ, Punt JA, Xu L, Karnell FG, Allman D, Myung PS, Boerth NJ, Pui JC, Koretzky GA, Pear WS: Notch1 regulates maturation of CD4+ and CD8+ thymocytes by modulating TCR signal strength. Immunity. 2001, 14: 253-264. 10.1016/S1074-7613(01)00107-8.
Tumang JR, Owyang A, Andjelic S, Jin Z, Hardy RR, Liou M-L, Liou H-C: C-Rel is essential for B lymphocyte survival and cell cycle progression. Eur. J. Immunol. 1998, 28: 4299-4312. 10.1002/(SICI)1521-4141(199812)28:12<4299::AID-IMMU4299>3.0.CO;2-Y.
Pear WS, Nolan GP, Scott ML, Baltimore D: Production of high-titer helper-free retroviruses by transient transfection. Proc. Natl. Acad. Sci. USA. 1993, 90: 8392-8396.
Wang CY, Mayo MW, Korneluk RG, Goeddel DV, Baldwin AS Jr: NF-kB antiapoptosis: induction of TRAF1 and TRAF2 and c-IAP1 and c-IAP2 to suppress caspase-8 activation. Science. 1998, 281: 1680-1683. 10.1126/science.281.5383.1680.
Zong W-X, Edelstein LC, Chen C, Bash J, Gelinas C: The prosurvival Bcl-2 homolog Bfl-1/A1 is a direct transcriptional target of NF-kB that blocks TNF-induced apoptosis. Genes & Dev. 1999, 13: 382-387.
Lee HH, Dadgostar H, Cheng Q, Shu J, Cheng G: NF-kappaB-mediated up-regulation of Bcl-x and Bfl-1/A1 is required for CD40 survival signaling in B lymphocytes. Proc Natl Acad Sci U S A. 1999, 96: 9136-9141. 10.1073/pnas.96.16.9136.
Grumont RJ, Rourke IJ, Gerondakis S: Rel-dependent induction of A1 transcription is required to protect B cells from antigen receptor ligation-induced apoptosis. Genes Dev. 1999, 13: 400-411.
Owyang AM, Tumang JR, Schram BR, Hsia CY, Behrens TW, Rothstein TL, Liou HC: c-Rel is required for the protection of B cells from antigen receptor- mediated, but not Fas-mediated, apoptosis. J Immunol. 2001, 167: 4948-4956.
Pizzi M, Goffi F, Boroni F, Benarese M, Perkins SE, Liou HC, Spano P: Opposing roles for NF-kappa B/Rel factors p65 and c-Rel in the modulation of neuron survival elicited by glutamate and interleukin- 1beta. J Biol Chem. 2002, 277: 20717-20723. 10.1074/jbc.M201014200.
Hsia CY, Cheng S, Owyang AM, Dowdy SF, Liou HC: c-Rel regulation of the cell cycle in primary mouse B lymphocytes. Int Immunol. 2002, 14: 905-916. 10.1093/intimm/dxf055.
Cheng S, Hsia CY, Leone G, Liou H-C: Cyclin E and Bcl-xL cooperatively induce cell cycle progression in c-Rel-/- B cells. Oncogene. 2003, in press:
Grumont RJ, Rourke IS, O' Reilly ZA, Strasser A, Miyake K, Sha W, Gerondakis S: B Lymphocytes differentially use the Rel and nuclear factor Kappa β1 (NF-kappaβ1) transcription factor to replicate cell cycle progression and apoptosis in quiescent and nitrogen activated cells. J EXP Med. 1998, 187: 663-674. 10.1084/jem.187.5.663.
Burkly L, Hession C, Ogata L, Reilly C, Marconi LA, Olson D, Tizard R, Cate R, Lo D: Expression of relB is required for the development of thymic medulla and dendritic cells. Nature. 1995, 373: 531-536. 10.1038/373531a0.
Wu L, D'Amico A, Winkel KD, Suter M, Lo D, Shortman K: RelB is essential for the development of myeloid-related CD8alpha- dendritic cells but not of lymphoid-related CD8alpha+ dendritic cells. Immunity. 1998, 9: 839-847.
Weih F, Carrasco D, Bravo R: Constitutive and inducible Rel/NF-kappa B activities in mouse thymus and spleen. Oncogene. 1994, 9: 3289-3297.
Ouaaz F, Arron J, Zheng Y, Choi Y, Beg AA: Dendritic cell development and survival require distinct NF-kappaB subunits. Immunity. 2002, 16: 257-270. 10.1016/S1074-7613(02)00272-8.
Boffa D, Feng B, Sharma V, Dematteo R, Suthanthiran M, Nunez R, Liou H-C: Selective loss of c-Rel compromises dendritic cell activation of T lymphocytes. Cell. Immunol. 2003, In press:
Grillot DA, Merino R, Pena JC, Fanslow WC, Finkelman FD, Thompson CB, Nunez G: bcl-x exhibits regulated expression during B cell development and activation and modulates lymphocyte survival in transgenic mice. J Exp Med. 1996, 183: 381-391.
Fang W, Weintraub BC, Dunlap B, Garside P, Pape KA, Jenkins MK, Goodnow CC, Mueller DL, Behrens TW: Self-reactive B lymphocytes overexpressing Bcl-xL escape negative selection and are tolerized by clonal anergy and receptor editing. Immunity. 1998, 9: 35-45.
Turner M, Gulbranson-Judge A, Quinn ME, Walters AE, MacLennan IC, Tybulewicz VL: Syk tyrosine kinase is required for the positive selection of immature B cells into the recirculating B cell pool. J Exp Med. 1997, 186: 2013-2021. 10.1084/jem.186.12.2013.
Schiemann B, Gommerman JL, Vora K, Cachero TG, Shulga-Morskaya S, Dobles M, Frew E, Scott ML: An essential role for BAFF in the normal development of B cells through a BCMA-independent pathway. Science. 2001, 293: 2111-2114. 10.1126/science.1061964.
Thompson JS, Bixler SA, Qian F, Vora K, Scott ML, Cachero TG, Hession C, Schneider P, Sizing ID, Mullen C, Strauch K, Zafari M, Benjamin CD, Tschopp J, Browning JL, Ambrose C: BAFF-R, a newly identified TNF receptor that specifically interacts with BAFF. Science. 2001, 293: 2108-2111. 10.1126/science.1061965.
Su TT, Guo B, Kawakami Y, Sommer K, Chae K, Humphries LA, Kato RM, Kang S, Patrone L, Wall R, Teitell M, Leitges M, Kawakami T, Rawlings DJ: PKC-beta controls I kappa B kinase lipid raft recruitment and activation in response to BCR signaling. Nat Immunol. 2002, 3: 780-786.
Esslinger CW, Wilson A, Sordat B, Beermann F, Jongeneel CV: Abnormal T lymphocyte development induced by targeted overexpression of IkappaB alpha. J Immunol. 1997, 158: 5075-5078.
Boothby MR, Mora AL, Scherer DC, Brockman JA, Ballard DW: Perturbation of the T lymphocyte lineage in transgenic mice expressing a constitutive repressor of nuclear factor (NF)-kappaB. J Exp Med. 1997, 185: 1897-1907. 10.1084/jem.185.11.1897.
Gerondakis S, Grossmann M, Nakamura Y, Pohl T, Grumont R: Genetic approaches in mice to understand Rel/NF-kappaB and IkappaB function: transgenics and knockouts. Oncogene. 1999, 18: 6888-6895. 10.1038/sj.onc.1203236.
Voll RE, Jimi E, Phillips RJ, Barber DF, Rincon M, Hayday AC, Flavell RA, Ghosh S: NF-kappa B activation by the pre-T cell receptor serves as a selective survival signal in T lymphocyte development. Immunity. 2000, 13: 677-689. 10.1016/S1074-7613(00)00067-4.
Acknowledgements
The authors are grateful for the professional service and care of experimental animals provided by the Research Animal Resources Center at the Weill Medical College of Cornell University. We would like to thank Dr Christopher Colon at the Flow Cytometry Core Facility for four-color flow cytometric analyses. We also appreciate Drs. Wenzhi Tian, and Constance Hsia for their critical comments on the manuscript. This work was supported by the funding from the National Institutes of Health. H.C.L. and W. S. P. are Scholars of The Leukemia and Lymphoma Society.
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Feng, B., Cheng, S., Pear, W.S. et al. NF-kB inhibitor blocks B cell development at two checkpoints. Med Immunol 3, 1 (2004). https://doi.org/10.1186/1476-9433-3-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1476-9433-3-1