
Abstract
Background
Hyperlipidemia associated with obesity is closely related to the development of atherosclerosis. Both n-3 polyunsaturated fatty acids (PUFAs) and long-chain monounsaturated fatty acids (MUFAs; i.e., C20:1 and C22:1 isomers) supplementation modulate risk factors for metabolic syndrome via multiple mechanisms, including the restoration of impaired lipid metabolism. We therefore examined the effects of pollock oil, which contains a considerable amount of n-3 PUFAs as well as long-chain MUFAs, on plasma hyperlipidemia and hepatic steatosis in diet-induced obese mice.
Methods
Male C57BL/6J mice (24-26 g) were divided into two groups (n = 10/group) and were fed a high-fat diet containing 32% lard (control group) or 17% lard plus 15% pollock oil (experimental group) for 6 weeks. For both groups, fat comprised 60% of the total caloric intake.
Results
Although body and liver masses for the two groups did not differ significantly, hepatic lipids concentrations (triglycerides and total cholesterols) were lower (P < 0.05) after pollock oil ingestion. After 2 weeks on the specified diets, plasma lipid levels (total cholesterol, LDL cholesterol, and triglycerides) significantly decreased (P < 0.05) in the experimental group compared with the control group, although plasma HDL cholesterol levels did not differ. At the end of 6 weeks, plasma adiponectin levels increased (P < 0.05), whereas plasma resistin and leptin levels decreased (P < 0.05) in the experimental mice. Increased levels of long-chain MUFAs and n-3 PUFAs in plasma, liver and adipose tissue by ingesting pollock oil were possibly correlated to these favorable changes. Expression of hepatic genes involved in cholesterol metabolism (SREBP2, HMGCR, and ApoB) and lipogenesis (SREPB1c, SCD-1, FAS, and Acac α) was suppressed in the experimental group, and may have favorably affected hyperlipidemia and hepatic steatosis induced by the high-fat diet.
Conclusions
We demonstrated that pollock oil supplementation effectively improved hyperlipidemia, attenuated hepatic steatosis, and downregulated the express of hepatic genes involved in cholesterol and lipid metabolism in mice with diet-induced obesity.
Similar content being viewed by others


Background
Hyperlipidemia, a medical condition characterized by increased blood levels of lipids including cholesterol and triglycerides, is a critical component of metabolism syndrome as well as a possible predisposing factor for atherosclerosis, a leading cause of death worldwide [1, 2]. Lipids that accumulate in the arterial wall as a consequence of hyperlipidemia are oxidized and attract inflammatory monocytes, which differentiate into macrophages that take up the oxidized lipid. Oxidation of low-density lipoproteins (LDLs) in the arterial wall is a major and physiologically relevant mechanism for the pathogenesis of atherosclerosis, and the presence of lipid-loaded macrophage foam cells in the artery intima is a predictor for the development of atherosclerotic lesions. A close relationship exists between dietary fats and dyslipidemia-related events [3]. Although an increased intake of saturated fatty acids is pathogenic for coronary heart disease, numerous studies have demonstrated a protective effect of n-3 polyunsaturated fatty acids (PUFAs) through a variety of mechanisms, including reduction of triglycerides and very-low-density lipoproteins [4]. In addition, we have shown that marine-derived long-chain monounsaturated fatty acids (MUFAs) (i.e., C20:1 and C22:1 isomers) modulate metabolic syndrome by restoring impaired glucose and lipid metabolism [5]. Therefore, fish oils that are rich in both n-3 PUFAs and long-chain MUFAs may help alleviate hypercholesterolemia and hypertriacylglyceridemia.
Alaska pollock (Theragra chalcogramma) is a North Pacific species of the cod family, Gadidae. Pollock oil contains considerable amounts of n-3 PUFAs and long-chain MUFAs [6]. The Alaska pollock fishing industry is the largest in the United States and one of the largest in the world. In recent years, pollock fishing has accounted for ~30% of all U.S. seafood landings by mass [7]. Although pollock oil is used in both the food and feed industries [8], little is known about the relationship between dietary pollock oil and hyperlipidemia. Given the health benefits of n-3 PUFAs and long-chain MUFAs, we examined the effect of dietary pollock oil on hyperlipidemia in mice with diet-induced dyslipidemia.
Methods
Measurement of fatty acid composition of dietary oils
Cameria lard was purchased from Romi Smilfood B. V. (Heerenveen, Netherlands). Pollock oil was obtained from Nippon Suisan Kaisha, Ltd. (Tokyo, Japan) and refined with silica gel and activated clays and then steam-distillation deodorized. All standard and extracted lipids were stored at -20°C until used. Fatty acid compositions of the dietary fats (Table 1) were determined after methylation of samples with 14% (w/v) boron trifluoride/methanol (Sigma Chemical Co., St. Louis, USA.) at 80°C for 30 min. The resulting fatty acid methyl esters were quantified by gas chromatography using an Agilent 6890N Network Gas Chromatograph System (Agilent Technologies Japan, Ltd., Tokyo, Japan) equipped with a split injector, FID detector, and fused silica capillary column (DB-WAX, 30 m × 0.25 mm I.D. × 0.25 μm film thickness, J & W Scientific, Agilent Technologies). Methyl esters were identified by comparison of retention times to those of fatty acid methyl ester standards (Nu-Chek Prep, Inc., Elysian, MN, USA). Pollock oil contains considerable levels of long-chain MUFAs and n-3 PUFAs (C20:1 as well as C22:1 isomers and n-3 PUFAs combined: ~ 45%).
Animals and diets
The Institutional Animal Care and Use Committee at Nihon Bioresearch Inc. (Gifu, Japan) approved this study. Male C57BL/6J mice (5 weeks old) were obtained from Charles River Laboratories Japan Inc. (Yokohama, Japan) and housed at Nihon Bioresearch at 23 ± 1°C under a 12/12 h light-dark cycle. The animals were provided free access to water and standard mouse chow CRF-1 (Oriental Yeast Co. Ltd., Tokyo, Japan) for a 1-week acclimatization period.
After acclimatization, mice were randomly assigned to one of two groups for the 6-week feeding experiment. The control group (n = 10) was fed a high-fat diet containing 32% lard (D12492 Rodent Diet with 60 kcal% Fat; Research Diets, Inc., NJ, USA) and the experimental group was fed a diet supplemented with pollock oil (17% lard plus 15% pollock oil). To control for total fat intake, the total fat content in both diets corresponded to 60% of the caloric intake. The compositions of the diets are listed in Table 2. Body mass and food intake were monitored throughout the study. At the end of the intervention period, mice were anesthetized with 4% sodium pentobarbital (Dainippon Sumitomo Pharma, Osaka, Japan), and blood was collected by abdominal vein puncture. Plasma was obtained by centrifugation at 1000 g for 15 min and stored at -80°C until analyses. Vital organs were removed and weighed after a short wash in cold phosphate-buffered saline, pH 7.4. Mesenteric white adipose tissue (WAT) and Livers were kept at -80°C for the further lipid extraction and quantitative polymerase chain reaction (QPCR) analysis.
Lipid extraction and fatty acid analysis
The fatty acid compositions of plasma, liver and WAT in the C57BL/6J mice were determined as described before [5]. Lipids were extracted by homogenizing the tissue samples in a methanol/hexane solution (4:1 v/v) added with butylated hydroxytoluene (BHT, 50 μg/mL) as an antioxidant. The samples were methylated with acetyl chloride, and the fatty acid methyl esters were separated and quantified by gas chromatography. Identification of the methyl esters were made by comparison of retention times of standard fatty acids.
Determination of plasma lipid levels
Blood samples were taken from the retro-orbital venous plexus of each mouse at the end of weeks 0, 2, 4, and 6. Plasma concentrations of triglyceride (TG), total cholesterol (TC), and high-density lipoprotein (HDL) cholesterol were measured using Triglycerol E-Test, Cholesterol E-Test, and HDL-cholesterol E-Test kits (Wako Pure Chemical Industries, Ltd., Osaka, Japan), respectively. The concentration of LDL cholesterol was calculated as [LDL cholesterol] = [TC] - [HDL cholesterol] - [TG] × 0.2.
Determination of plasma adipokine levels
Plasma concentrations of adiponectin, resistin, and leptin were determined at the end of the 6-week period using the Mouse Adiponectin ELISA kit (Otsuka Pharmaceutical Co., Ltd., Tokyo, Japan), Mouse Resistin ELISA kit (Shibayagi Co. Ltd., Gunma, Japan), and Mouse Leptin ELISA kit (Morinaga Institute of Biological Science, Inc., Yokohama, Japan), respectively.
Determination of hepatic lipid levels
Total hepatic lipids were extracted from liver samples as described [9]. Extracted lipids were dried under vacuum (Concentrator Plus 5305, Eppendorf Inc., NY, USA) and then dissolved in 2-propanol containing 10% (w/w) Triton X-100. Triglyceride and total cholesterol concentrations were determined using the above-mentioned commercial enzyme kits (Wako).
Determination of mRNA expression by QPCR
Total RNA was isolated from liver samples using TRIzol reagent (Qiagen, Valencia, CA, USA) according to the manufacturer's protocol. First-strand cDNA was generated from total RNA (1 μg) using the PrimeScript II 1st strand cDNA Synthesis kit (TaKaRa Bio, Otsu, Japan). The resulting cDNA was used for QPCR amplification and specific sequence detection on an Applied Biosystems 7300 Real-Time PCR System (Life Technologies Ltd., Tokyo, Japan). The PCR cycling parameters were 30 s at 95°C; 40 cycles of 5 s at 95°C, 34 s at 60°C; and a final melting curve of 15 s at 95°C, 1 min at 60°C, 15 s at 95°C. Gene expression was scaled to the expression of the housekeeping gene encoding 18S ribosomal RNA. PCR reactions contained forward and reverse primers (10 μM each) and SYBR Premix Ex Taq (TaKaRa Bio). The targeted genes, their corresponding proteins, and the respective sense and antisense PCR primers were: SREBP2 (sterol regulatory element binding protein 2), 5'- TGGGCGATGAGCTGACTCT-3' and 5'- ACTGTAGCATCTCGTCGATGT-3'; HMGCR (3-hydroxy-3-methylglutaryl-coenzyme A reductase), 5'- TGTTCACCGGCAACAACAAGA-3' and 5'-CCGCGTTATCGTCAGGATGA-3'; ApoB (apolipoprotein B), 5'-TTGGCAAACTGCATAGCATCC-3' and 5'-TCAAATTGGGACTCTCCTTTAGC-3'; ApoA (apolipoprotein A), 5'- GGCACGTATGGCAGCAAGAT-3' and 5'-CCAAGGAGGAGGATTCAAACTG-3'; SREBP1c (sterol regulatory element binding protein 1c), 5'-GATGTGCGAACTGGACACAG-3' and 5'-CATAGGGGGCGTCAAACAG-3'; SCD-1 (stearoyl-coenzyme A desaturase-1), 5'-TTCTTGCGATACACTCTGGTGC-3' and 5'-CGGGATTGAATGTTCTTGTCGT-3'; FAS (fatty acid synthase), 5'-TTCTTGCGATACACTCTGGTGC-3' and 5'-CGGGATTGAATGTTCTTGTCGT-3'; Acacα (acetyl-coenzyme A carboxylase alpha), 5'-GATGAACCATCTCCGTTGGC-3' and 5'-CCCAATTATGAATCGGGAGTGC-3'.
Statistical analysis
Results are expressed as mean ± standard error of the mean. Statistical differences between two groups were analyzed by Student's t-test and were considered significant at P < 0.05.
Results
Effect of pollock oil on body and organ mass
Table 3 lists body and vital organ masses for diet-induced obese C57BL/6J mice in the control (lard) and experimental (pollock oil) groups. After the 6-week feeding period, there were no significant differences in body, liver, white as well as brown adipose tissue, and skeleton muscle masses between the two groups.
Effect of pollock oil on plasma lipid levels
Compared to control, total cholesterol plasma levels for the experimental group were reduced by 27% (P < 0.01), 17% (P < 0.01), and 30% (P < 0.001) at the end of weeks 2, 4, and 6, respectively (Table 4). Pollock oil intake also significantly reduced plasma LDL cholesterol levels by 33% (P < 0.05), 23% (P < 0.01), and 38% (P < 0.001), and reduced plasma triglyceride levels by 40% (P < 0.01), 50% (P < 0.01), and 46% (P < 0.01) at the end of weeks 2, 4, and 6, respectively. No significant differences in plasma HDL cholesterol levels were detected.
Effect of pollock oil on fatty acid compositions of plasma, liver and WAT
Plasma, liver and mesenteric WAT fatty acid compositions in the control and pollock oil group are shown in Table 5 and Table 6. Although total saturated fatty acid levels did not differ between the control and experimental group, pollock oil ingestion markedly (P < 0.05) increased long-chain MUFA (i.e., C20:1 and C22:1 isomers combined) levels 3-, 1.2- and 5-fold in plasma, liver and WAT, respectively. Intake of pollock oil also significantly (P < 0.05) increased total n-3 PUFA levels 3-, 2- and 7-fold in plasma, liver and WAT, respectively. In contrast, total n-6 PUFA levels were significantly (P < 0.05) decreased by 50%, 31% and 14% in plasma, liver and WAT, respectively in the pollock oil group as compared to the control.
Effect of pollock oil on plasma adipocytokine levels
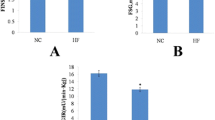
To determine if pollock oil could relieve the adipokine dysregulation caused by excessive total caloric intake, we measured the adipokine plasma levels. Intake of pollock oil increased the plasma concentration of adiponectin by 15% (P < 0.05) in the experimental group compared to the control group (Figure 1A). Plasma resistin and leptin concentrations were reduced by 14% (P < 0.05) and 41% (P < 0.05), respectively (Figure 1B and 1C).

Effect of pollock oil on plasma adipokine levels in mice fed a high-fat diet. Plasma levels of adiponectin (A), resistin (B), and leptin (C) in C57BL/6J mice fed for 6 weeks with a diet containing 32% lard (lard diet) or 17% lard plus 15% pollock oil (PO diet). Values are mean ± SE (n = 10). *P < 0.05.
Effect of pollock oil on hepatic steatosis
Because obesity can trigger hepatic steatosis, which is associated with hyperlipidemia, we measured the levels of hepatic lipids to determine if pollock oil suppressed hepatic lipid accumulation. Total hepatic lipid, triglyceride, and total cholesterol levels were reduced by 21% (P < 0.01), 40% (P < 0.001), and 12% (P < 0.05), respectively, in the experimental group compared with the control group (Figure 2).

Effect of pollock oil on hepatic steatosis in mice fed a high-fat diet. Total hepatic lipid (A), triglyceride (B), and total cholesterol (C) levels in C57BL/6J mice fed for 6 weeks with a diet containing 32% lard (lard diet) or 17% lard plus 15% pollock oil (PO diet). Values are mean ± SE (n = 10). *P < 0.05; **P < 0.01; ***P < 0.001.
Effect of pollock oil on mRNA expression of genes involved in hepatic lipid metabolism
The pollock oil-supplemented diet led to a 66% reduction (P < 0.001) in mRNA expression of SREBP2, a gene that encodes a transcription factor involved mainly in regulation of cholesterol synthesis (Figure 3A). It also reduced expression of the cholesterogenic gene HMGCR by 51% (P < 0.05) and of ApoB by 23% (P < 0.05), although it had no significant effect on expression of ApoA. Furthermore, intake of pollock oil caused a 24% (P < 0.05) reduction in mRNA expression of SREBP1c, which encodes for the lipogenic transcription factor SREBP1c, and also reduced expression of the downstream lipogenic genes SCD-1, FAS, and Acac α by 68% (P < 0.001), 69% (P < 0.001), and 33% (P < 0.05), respectively (Figure 3B).

Effect of pollock oil on transcription of genes related to hepatic cholesterol and lipid metabolism. mRNA levels for genes involved in cholesterol (A) and lipid (B) metabolism in C57BL/6J mice fed for 6 weeks with a diet containing 32% lard (lard diet) or 17% lard plus 15% pollock oil (PO diet). Values are mean ± SE (n = 10). *P < 0.05; ***P < 0.001.
Discussion
People with obesity have an increased risk of cardiovascular disease, which is a major cause of their increased mortality. Although multiple factors are thought to contribute to these elevated risks, one main determinant is the adverse effect of obesity on lipoprotein levels [10]. For the study reported herein, we examined the effect of pollock oil on dyslipidemia in diet-induced obese mice, and showed that inclusion of pollock oil in a high-fat diet significantly decreased plasma levels of total and LDL cholesterol and triglyceride.
To identify possible mechanisms underlying this reduction, we measured plasma adipokine levels. Adiponectin, an adipocyte-derived hormone, is believed to play an important role in regulating hyperglycemia, hyperlipidemia, and endothelial dysfunction in humans, all of which probably contribute to certain markedly increased risks associated with obesity-related disorders, e.g., atherosclerosis and diabetes [11]. Adiponectin levels correlate negatively with serum triglyceride levels in non-diabetic subjects and type-2 diabetics, and hypoadiponectinemia is associated with smaller LDL particle size, indicating a link between adiponectin and dyslipidemia [12–15]. Furthermore, in vitro data indicate that adiponectin is an anti-inflammatory and anti-proliferative mediator that can modulate atherosclerosis progression [16–18]. Conversely, increased levels of proinflammatory adipokines (e.g., tumor necrosis factor α, resistin, monocyte chemotactic protein 1, and interleukin 8) have been associated with increased serum lipids (e.g., triglycerides, and total and LDL cholesterol), which increase monocyte recruitment and adherence to arterial walls, causing wall remodeling [19–22]. In addition, leptin, the first adipocytokine discovered, has been associated with development of obesity, as ob/ob leptin-deficient mice are markedly obese [23]. However, most obese humans have increased blood leptin concentrations, likely reflecting resistance to the action(s) of leptin [24]. Leptin is also associated with increased insulin resistance, which can cause hyperlipidemia [25]. Therefore, the decreased plasma lipid concentrations observed following pollock oil ingestion were possibly associated with elevated plasma adiponectin levels and reduced plasma proinflammatory adipokine levels. As endogenous ligands for peroxisome proliferator-activated receptors, n-3 PUFAs regulate the expression of genes encoding key proteins involved in metabolism [26, 27]. Intake of fish oil enriched in n-3 PUFAs has been reported to increase plasma adiponectin levels and lower proinflammatory adipokine levels [28, 29]. Furthermore, we have shown that a diet rich in long-chain MUFAs modulates adipokine profiles [5]. Fatty acid composition analyses revealed that pollock oil ingestion significantly increased long-chain MUFA and n-3 PUFA levels in plasma and vital organs. Therefore, the favorable changes in the plasma adipokine profile might be attributed to a combined effect of n-3 PUFAs and long-chain MUFAs found abundantly in pollock oil.
Ingestion of pollock oil also decreased hepatic lipid levels in the experimental group, indicating that pollock oil inhibited hepatic steatosis triggered by obesity. The liver is the most important organ in energy metabolism, and is vital to the production and catabolism of plasma lipoproteins and endogenous lipids [30, 31]. The improvement in fatty filtration in the livers of pollock oil-fed mice was possibly related to the decreased plasma lipid levels. To characterize how pollock oil decreased hepatic lipid accumulation, we also determined the hepatic mRNA levels of genes related to lipid metabolism. In the liver, sterol regulatory element-binding proteins (SREBPs) are key transcription factors that regulate the levels of lipids produced for export into the plasma as lipoproteins and into the bile as micelles. The SREBP family includes SREBP1a, 1c, and 2 [32]. SREBP1c regulates transcription of genes involved in fatty acid metabolism (e.g., SCD-1, FAS, and Acac α), and SREBP2 regulates transcription of cholesterol-related genes (e.g., HMGCR, which encodes the rate-limiting enzyme of cholesterol biosynthesis). Increased SREBP activity causes cholesterol and fatty acid accumulation and downregulates the SREBP-cleavage-activating protein (SCAP)/SREBP pathway by feedback inhibition [33]. Our data show that, concomitant with downregulation of SREBP1c and SREBP2 mRNA, intake of pollock oil suppressed transcription of genes targeted by SREBP. Furthermore, pollock oil intake inhibited hepatic ApoB transcription. Apolipoprotein B is central in lipoprotein metabolism [34], serving as a structural and functional component of triglyceride-rich very-low-density lipoproteins and their metabolic products, e.g., intermediate-density lipoprotein and LDL. Downregulation of ApoB expression may therefore decrease circulating LDL cholesterol levels. Notably, pollock oil ingestion did not alter mRNA expression of ApoA, the structural component of HDL, which may reflect the fact that plasma HDL cholesterol levels were the same in the control and experimental groups. Collectively, the observed decreases in mRNA levels suggest that the improvements in hepatic lipid levels and associated hyperlipidemia found in the experimental group may be partially associated with decreased de novo cholesterol, lipid, and ApoB synthesis.
Conclusions
Our study showed that ingestion of pollock oil ameliorated hypercholesterolemia and hypertriacylglyceridemia in diet-induced obese mice. The hypolipidemic effect of pollock oil was possibly related to an increase in plasma adiponectin concentrations and a decrease in the plasma levels of proinflammatory adipokines. Downregulation of mRNA expression of lipogenic genes and genes involved in cholesterol metabolism positively affected hepatic lipid accumulation, which likely led to an improved plasma lipid profile.
Abbreviations
- Acacα:
-
acetyl-coenzyme A carboxylase alpha
- ApoA:
-
apolipoprotein A
- ApoB:
-
apolipoprotein B
- FAS:
-
fatty acid synthase
- HDL:
-
high-density lipoprotein
- HMGCR:
-
3-hydroxy-3-methylglutaryl-coenzyme A reductase
- LDL:
-
low-density lipoprotein
- MUFA:
-
monounsaturated fatty acids
- PUFA:
-
polyunsaturated fatty acids
- QPCR:
-
quantitative polymerase chain reaction
- SCD-1:
-
stearoyl-coenzyme A desaturase-1
- SREBP:
-
sterol regulatory element binding protein
- WAT:
-
white adipose tissue.
References
Holvoet P: Relations between metabolic syndrome, oxidative stress and inflammation and cardiovascular disease. Verh K Acad Geneeskd Belg. 2008, 70: 193-219.
Meydani M: Vitamin E and atherosclerosis: beyond prevention of LDL oxidation. J Nutr. 2001, 131: 366S-368S.
Lairon D, Defoort C: Effects of nutrients on postprandial lipemia. Curr Vasc Pharmacol. 2011, 9: 309-312. 10.2174/157016111795495576
Connor WE: n-3 Fatty acids and heart disease. Nutrition and disease update: heart disease. Edited by: Kritchevsky D, Carroll KK. 1994, 7-42. Champaign, IL: American Oil Chemists' Society,
Yang ZH, Miyahara H, Mori T, Doisaki N, Hatanaka A: Beneficial effects of dietary fish-oil-derived monounsaturated fatty acids on metabolic syndrome risk factors and insulin resistance in mice. J Agric Food Chem. 2011, 59: 7482-7489. 10.1021/jf201496h
Oliveira ACM, Bechtel PJ: Lipid composition of Alaska pink salmon (Oncorhynchus gorbuscha) and Alaska walleye pollock (theragra chalcogramma). J Aquat Food Prod Tech. 2005, 14: 73-91. 10.1300/J030v14n01_07.
Bailey KM, Powers DM, Quattro JM, Villa G, Nishimura A, Traynor JJ, Walters G: Population Ecology and Structural Dynamics of Walleye Pollock (Theragra chalcogramma). Dynamics of the Bering Sea. 1999, 581-614.
Bimbo AP: Technology of production and industrial utilization of marine oil. Marine biogenic lipids, fats, and oil. Edited by: RG Ackman. 1989, II: 401-433. Boca, Raton, Florida: CRC Press, Inc,
Folch J, Lees M, Stanley GHS: A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem. 1956, 226: 497-509.
National Heart, Lung, and Blood Institute: Clinical guidelines on the identification, evaluation, and treatment of overweight and obesity in adults-the evidence report. Obes Res. 1998, 6 (Suppl 2): 51S-209S.
Chandran M, Phillips SA, Ciaraldi T, Henry RR: Adiponectin: more than just another fat cell hormone?. Diabetes Care. 2003, 26: 2442-2450. 10.2337/diacare.26.8.2442
Matsubara M, Maruoka S, Katayose S: Decreased plasma adiponectin concentrations in women with dyslipidemia. J Clin Endocrinol Metab. 2002, 87: 2764-2769. 10.1210/jc.87.6.2764
Kazumi T, Kawaguchi A, Hirano T, Yoshino G: Serum adiponectin is associated with high-density lipoprotein cholesterol, triglycerides, and low density lipoprotein particle size in young healthy men. Metab Clin Exp. 2004, 53: 589-593. 10.1016/j.metabol.2003.12.008
Hulthe J, Hulten LM, Fagerberg B: Low adipocyte-derived plasma protein adiponectin concentrations are associated with the metabolic syndrome and small dense low-density lipoprotein particles: atherosclerosis and insulin resistance study. Metab Clin Exp. 2003, 52: 1612-1614. 10.1016/S0026-0495(03)00313-5
Schulze MB, Rimm EB, Shai I, Rifai N, Hu FB: Relationship between adiponectin and glycemic control, blood lipids, and inflammatory markers in men with type 2 diabetes. Diabetes Care. 2004, 27: 1680-1687. 10.2337/diacare.27.7.1680
Ouchi N, Kihara S, Arita Y, Nishida M, Matsuyama A, Okamoto Y, Ishigami M, Kuriyama H, Kishida K, Nishizawa H, Hotta K, Muraguchi M, Ohmoto Y, Yamashita S, Funahashi T, Matsuzawa Y: Adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation. 2001, 103: 1057-1063.
Yokota T, Oritani K, Takahashi I, Ishikawa J, Matsuyama A, Ouchi N, Kihara S, Funahashi T, Tenner AJ, Tomiyama Y, Matsuzawa Y: Adiponectin, a new member of the family of soluble defence collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood. 2000, 96: 1723-1732.
Kumada M, Kihara S, Ouchi N, Kobayashi H, Okamoto Y, Ohashi K, Maeda K, Nagaretani H, Kishida K, Maeda N, Nagasawa A, Funahashi T, Matsuzawa Y: Adiponectin specifically increased tissue inhibitor of metalloproteinase-1 through interleukin-10 expression in human macrophages. Circulation. 2004, 109: 2046-2049. 10.1161/01.CIR.0000127953.98131.ED
Jovinge S, Hamsten A, Tornvall P, Proudler A, Bavenholm P, Ericsson CG, Godsland I, de Faire U, Nilsson J: Evidence for a role of tumor necrosis factor alpha in disturbances of triglyceride and glucose metabolism predisposing to coronary heart disease. Metabolism. 1998, 47: 113-118. 10.1016/S0026-0495(98)90203-7
Nelken NA, Coughlin SR, Gordon D, Wilcox JN: Monocyte chemoattractant protein-1 in human atheromatous plaques. J Clin Invest. 1991, 88: 1121-1127. 10.1172/JCI115411
Stapleton PA, Goodwill AG, James ME, D'Audiffret AC, Frisbee JC: Differential Impact of Familial Hypercholesterolemia and Combined Hyperlipidemia on Vascular Wall and Network Remodeling in Mice. Microcirculation. 2010, 17: 47-58. 10.1111/j.1549-8719.2009.00003.x
Tilg H, Moschen AR: Adipocytokines: mediators linking adipose tissue, inflammation and immunity. Nat Rev Immunol. 2006, 6: 772-783. 10.1038/nri1937
Ingalls AM, Dickie MM, Snell GD: Obese, a new mutation in the house mouse. J Hered. 1950, 41: 317-318.
Martin SS, Qasim A, Reilly MP: Leptin resistance: a possible interface of inflammation and metabolism in obesity-related cardiovascular disease. J Am Coll Cardiol. 2008, 52: 1201-1210. 10.1016/j.jacc.2008.05.060
Koh KK, Park SM, Quon MJ: Leptin and cardiovascular disease: response to therapeutic interventions. Circulation. 2008, 117: 3238-3249. 10.1161/CIRCULATIONAHA.107.741645
Berger J, Moller DE: The mechanisms of action of PPARs. Annu Rev Med. 2002, 53: 409-435. 10.1146/annurev.med.53.082901.104018
Huss JM, Kelly DP: Nuclear receptor signaling and cardiac energetics. Circ Res. 2004, 95: 568-578. 10.1161/01.RES.0000141774.29937.e3
Das UN, Ramos EJ, Meguid MM: Metabolic alterations during inflammation and its modulation by central actions of omega-3 fatty acids. Curr Opin Clin Nutr Metab Care. 2003, 6: 413-419.
Browning LM: n-3 Polyunsaturated fatty acids, inflammation and obesity-related disease. Proc Nutr Soc. 2003, 62: 447-453. 10.1079/PNS2003252
Shepherd J: Lipoprotein metabolism: an overview. Ann Acad Med Singapore. 1992, 21: 106-113.
Tietge UJ, Boker KH, Bahr MJ, Weinberg S, Pichlmayr R, Schmidt HH, Manns MP: Lipid parameters predicting liver function in patients with cirrhosis and after liver transplantation. Hepatogastroenterology. 1998, 45: 2255-2260.
Osborne TF: Sterol regulatory element-binding proteins (SREBPs): key regulators of nutritional homeostasis and insulin action. J Biol Chem. 2000, 275: 32379-32382.
Goldstein JL, Rawson RB, Brown MS: Mutant mammalian cells as tools to delineate the sterol regulatory element-binding protein pathway for feedback regulation of lipid synthesis. Arch Biochem Biophys. 2002, 397: 139-148. 10.1006/abbi.2001.2615
Burnett JR, Barrett PHR: Apolipoprotein B metabolism: tracer kinetics, models, and metabolic studies. Crit Rev Clin Lab Sci. 2002, 39: 89-137. 10.1080/10408360208951113
Acknowledgements
We thank Mr. Nobushige Doisaki and Ms. Kiyomi Furihata in Nippon Suisan Kaisha, Ltd. for technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
ZHY participated in the planning of the study, data analysis, and manuscript preparation. HM participated in experimental work. JT, AH and MK participated in the planning and organization of the study. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Yang, ZH., Miyahara, H., Takeo, J. et al. Pollock oil supplementation modulates hyperlipidemia and ameliorates hepatic steatosis in mice fed a high-fat diet. Lipids Health Dis 10, 189 (2011). https://doi.org/10.1186/1476-511X-10-189
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1476-511X-10-189