
Abstract
Survivin (BIRC5), a member of the inhibitor of apoptosis protein (IAP) family that inhibits caspases and blocks cell death is highly expressed in cancer and is associated with a poorer clinical outcome. Functioning simultaneously during cell division and apoptosis inhibition, survivin plays a pivotal role in determining cell survival. Survivin has consistently been identified by molecular profiling analysis to be associated with higher tumor grade, more advanced disease, abbreviated survival, accelerated rates of recurrence, and chemotherapy and radiation resistance. Survivin's differential expression in cancer compared to normal tissue and its role as a nodal protein in a number of cellular pathways make it a highly flexible therapeutic target, suitable for small-molecule inhibitiors, molecular antagonists, and vaccination-based therapies. By targeting survivin it is hoped that multiple tumor signaling circuitries may be simultaneously disabled. This effect may be applicable to many tumor histologies irrespective of specific genetic makeup. To date, survivin inhibitors have shown modest activity as single agents, but it is anticipated that when given in combination with cytotoxic chemotherapy or monoclonal antibodies they may exhibit enhanced efficacy. This review discusses the complex circuitry of survivin in human cancers and highlights clinical trials involving novel agents that target this important protein.
Similar content being viewed by others

Introduction
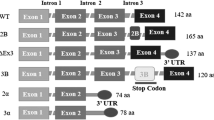
Survivin (BIRC5), is a member of the family of inhibitors of apoptosis proteins (IAPs) [1, 2] of which eight members are known, including X-linked inhibitor of apoptosis (XIAP), cIAP1, cIAP2, NAIP (NLR family, apoptosis inhibitory protein), livin, ILP2 (IAP-like protein 2), BRUCE and survivin [3, 4]. Survivin, the smallest family member, is a 142-amino acid, 16.5 kDa protein encoded by a single gene located on the human 17q25 chromosome, consisting of three introns, and four exons [2, 5, 6] and exists physiologically as a functional homodimer [7, 8]. Alternative splicing of survivin pre-mRNA produces five different mRNAs with the potential to encode up to five distinct proteins, survivin, survivin 2B, survivin ΔEx3, survivin 3B and survivin 2α[9–11]. Survivin has been implicated in both control of cell survival and regulation of mitosis in cancer [5, 12–14]. Survivin is preferentially and highly expressed in cancer cells, with little expression in most normal non-dividing adult tissues (Table 1) [5]. The integral role of survivin in cancer cell division and survival makes it an attractive therapeutic target to inhibit cancer cell growth [1, 2]. It was originally suggested that survivin inhibits cell death induced via the extrinsic and intrinsic apoptotic pathways and confers resistance to apoptosis by directly suppressing caspase activity [14]. Although the exact mechanism of action is unknown, current evidence is that most IAPs, including survivin, block apoptosis by mechanisms other than by direct initiator or effector caspase inhibition [15–17]. Survivin is now thought to function upstream of the effector caspases by inhibiting caspase 9[18], by forming a survivin-hepatitis B X-interacting protein (HBXIP) complex bound to pro-caspase-9 thereby preventing the recruitment of apoptotic protease activating factor 1 (Apaf-1) to the apoptosome [19]. Additionally survivin associates with XIAP enhancing its inhibition of caspase-9 activation [20]. Survivin is inhibited by SMAC/DIABLO (second mitochondria-derived activator of caspases/direct inhibitor of apoptosis binding protein with low pI) which results in the displacement of bound IAPs, which, may then bind to and inhibit caspase function [21, 22].
Some investigators have suggested that the primary function of survivin is in controlling cell division, rather than apoptosis inhibition [23, 24]. Survivin is up-regulated during cell division and is closely associated with centrosomes and mitotic spindle microtubules. It controls chromosome spindle-checkpoint assembly, thereby ensuring normal cell division. Survivin is maximally expressed during the G2M phase of the cell cycle and exists predominantly as a multi-protein complex, known as the chromosomal passenger complex (CPC) [25–27]. By functioning in this complex survivin can facilitate accurate sister chromatid segregation and stabilization of the microtubules in late mitosis [23]. In addition to its direct role in carcinogenesis, survivin may also play a key role in tumor angiogenesis as it is strongly expressed in endothelial cells during the proliferative phase of angiogenesis [12, 28, 29]. Manipulating the survivin pathway may facilitate endothelial cell apoptosis and promote vascular regression during tumor angiogenesis [29]. Increased expression of survivin also appears to be associated with an increased risk of tumor progression and chemoresistance in many tumor types [30–41]. Results of in vitro and in vivo studies have shown that survivin down-modulation reduces tumor-growth and sensitizes tumor cells to chemotherapeutic agents such as taxanes, platinum agents, etoposide, gamma-irradiation, and immunotherapy [42]. As an example, resistance to docetaxel is associated with increased levels of survivin [43], and response is often associated with the degree of expression of the various survivin splice variants [44].
Mechanism of Action of Survivin
Cellular apoptosis is controlled by two pathways. The extrinsic pathway is critical for immune selection and inflammation. It is initiated by the activation of cell death receptors, such as tumor necrosis factor alpha (TNF-α) receptor, located at the cell membrane. The intrinsic pathway is initiated by toxic insult such as radiation or chemotherapy (DNA damaging and anti-microtubule agents) [45]. The two pathways converge at caspase-3 and follow a common process of activating caspases that cause cell death by cleaving essential substrates for cell survival, such as cytoskeletal proteins, DNA repair proteins, and inhibitory subunits of endonucleases [46]. Upon activation of the intrinsic pathway, mitochondrial permeability is increased resulting in the release of both cytochrome C and SMAC/DIABLO. Cytochrome C activates the apoptosome, which in turn activates caspase-mediated proteolysis involved in cell death [47].
SMAC/DIABLO acts as an inhibitor of the IAPs. Upon activation of pro-apoptotic cell signaling, survivin is released from the mitochondria and inhibits caspases-3 and -9. This function requires association with hepatitis B X-interacting protein (HBXIP) and/or with X-linked IAP (XIAP) and is inhibited by SMAC-DIABLO [47]. The regulation of survivin expression and function is complex and occurs at various levels, including transcription, differential splicing, protein degradation, and intracellular sequestration via different ligands [47]. Survivin expression is up-regulated at a transcriptional level by both nuclear factor-κappa β (NF-κB) which in turn, can be activated indirectly by growth factors via the phosphatidylinositol 3-kinase (PI3K)/Akt pathway and by TCF-4/β-catenin pathway [48–50]. The insulin like growth factor-1/mTOR/RAS and Wnt-2 signaling pathways have also been reported to up-regulate survivin via rapid changes in mRNA translation [51–53]. Survivin degradation occurs via the ubiquitin-proteasome pathway in the G1 phase of the cell cycle and is stabilized when bound to heat shock protein 90 (Hsp90) [54]. The survivin protein is closely associated with Cdc2/Cdk1 and it is phosphorylated at the threonine-34 (T34) residue. This phosphorylation stabilizes the protein and allows it to interact with the mitotic spindle and inhibit caspase-9 [55].
There is accumulating evidence that molecular chaperones play a key role in the regulation of survivin. Binding of survivin to the immunophilin aryl hydrocarbon receptor-interacting protein (AIP) [56] or to heat shock protein-90 (Hsp90) [54] maintains its stability against proteasome dependent destruction. Heat shock protein-60 (Hsp60) has also been identified as a molecular chaperone for survivin [57]. Acute ablation of Hsp60 by small interfering RNA (siRNA) has been shown to destabilize the mitochondrial pool of survivin leading to enhanced mitochondrial dysfunction and caspase-dependent apoptosis. This response involves disruption of the Hsp60-p53 complex, which results in p53 stabilization, increased expression of pro-apoptotic Bax, and subsequent Bax-dependent apoptosis [57].
Survivin as a Regulator of Cell Division
Survivin plays a central role in cell division, where its expression is coordinated within the cell cycle [2, 47]. Survivin levels increase in G1 and peak in the G2M phase. During mitosis, survivin functions as a regulator of microtubule dynamics and as part of the chromosomal passenger complex (CPC). Survivin functions both at the centrosomes and the microtubules of the metaphase and anaphase spindle providing stabilization and ensuring accurate separation of sister chromatids [2, 58, 59]. Survivin also localizes to the kinetochores, the mid-region or centromeric portion of the metaphase chromosomes. Here survivin is associated with regulators of cytokinesis, such as Aurora B kinase, the inner centromere protein antigens (INCENP), and Borealin/Dasra [60, 61]. This supports the hypothesis that survivin acts as a subunit of the CPC which is required for proper chromosome segregation and cytokinesis [62]. It follows that if survivin is removed from the system, the kinetochore-microtubule system is not properly formed, cell division is either halted or improperly completed, and ultimately cell death occurs. Survivin is also bound to microtubules located in the mitotic apparatus during cell division. Through its association with cyclin-dependent kinase 1 (CDK1), microtubule-bound survivin becomes phosphorylated on Thr34 [1]. This leads to stabilization of the protein and efficient counter-activation of apoptosis in dividing cells. Elimination of survivin leads to apoptosis of dividing cells.
Survivin Expression in Cancer Cells
Survivin is undetectable in most non-proliferating adult tissues. Exceptions are CD34+ hematopoietic stem cells, placenta, basal cells of the colonic epithelium, and thymus [5]. On the other hand, survivin is over-expressed in a wide variety of cancers (see Table 1) [5]. Overexpression is correlated with advanced disease, accelerated time to recurrence, reduced survival, and resistance to therapy [38]. Survivin has been identified as one of 16 genes within an mRNA expression signature that, in patients with breast cancer, correlates with a poor response to tamoxifen, but a good response to chemotherapy [63]. Furthermore, inhibition of in vitro or in vivo survivin expression by antisense oligonucleotides, or inhibition of survivin function by in vitro inhibition of CDK1-mediated phosphorylation leads to cell death [58]. This is particularly apparent when CDK1 inhibition is combined with taxanes [58]. Recently the mTOR pathway, which can act as a sensor network in stressed conditions, has been implicated in elevating survivin levels [51]. In prostate cancer cells, insulin-like growth factor-1 (IGF-1)-mediated mTOR pathway activation positively modulated survivin levels by increasing the translation of a survivin mRNA pool [51].
The role of survivin in malignant melanoma
Genes that are highly expressed in aggressive melanomas includes many with roles in cell cycle regulation/proliferation, DNA replication/repair, and apoptosis pathway-related genes. Survivin is variably expressed in the cytoplasm across the spectrum of melanocytic lesions, with nuclear expression detectable in a subset of malignant melanomas, but not in benign or dysplastic nevi [64]. Nuclear expression has been reported to be an independent predictor for disease recurrence and decreased overall survival in patients with early stage cutaneous melanoma [65]. Patients with nuclear immunoreactivity for survivin showed an increased risk of melanoma recurrence during the first three post-operative years and an increased risk of death. Cytoplasmic survivin staining has not demonstrated a correlation with patient survival [65]. This suggests that the nuclear localization of survivin may play a role in the transformation process. Chen and co-workers demonstrated the up-regulation of survivin in primary melanomas as compared to benign nevi [66]. Similarly, Nasr et al found immune-histochemistry (IHC) nuclear staining for survivin was present in 12 of 18 cases (67%) of malignant melanoma with an average index of 7% (range 0%-15%), and no nuclear staining was present in any benign lesions examined [67]. Expression of survivin, bcl-2, bax, and bcl-X in sentinel lymph node (SLN) biopsies was assessed in 36 patients with stage I and II melanoma using RT-PCR and Southern blotting and correlated to overall survival. Survivin expression correlated with the outcome of patients in a statistically significant way (P < 0.005), whereas the expression of bcl-2, bax, and bcl-X, did not seem to correlate to progression of disease [68]. In this study, 61.5% of patients expressing survivin in the SLN progressed or died of melanoma. In patients negative for survivin expression, 100% were disease-free at a median follow-up time of 52.9 months suggesting that survivin gene expression in SLNs may be a useful prognostic indicator. There was no correlation between bcl-2, bax, and bcl-X gene expression and outcome [68]. Survivin expression has been studied in a limited number of non-cutaneous melanomas. In uveal melanomas, expression of survivin was reported to be low and did not correlate with outcome, resistance of the tumor to brachytherapy, or the presence of liver metastases [69]. Survivin expression has also been reported in a case of esophageal melanoma [70], but its impact is unclear. A phase II study investigated YM155 (section 5.1), a small molecule survivin suppressor as a single agent first line treatment in 34 patients with metastatic melanoma. One patient had a complete response, one had a partial response, and 11 had stable disease97.
The role of survivin in solid tumors
Survivin expression has been detected in a wide variety of benign and pre-neoplastic lesions including polyps of the colon, breast adenomas, Bowen's disease and hypertrophic actinic keratosis [71]. Survivin has been shown to play an important role in colorectal tumorigenesis by stimulating the transition of adenomas with mild dysplasia into highly dysplastic lesions [72]. This transition was associated with a significant decrease in the apoptotic index (AI) and significant increases in the Ki-67 labeling index (LI) and microvessel density (MVD) (P < 0.001). The expression of survivin inversely correlated with AI and was positively correlated with Ki-67 LI and MVD (P < 0.001). In esophageal cancer, survivin expression has been correlated with a poor prognosis whereby the median survival for patients with high levels of survivin expression was reduced compared to patients with low expression levels [73]. In this study the median survival for patients with advanced esophageal cancer and with high levels of survivin expression was 9.0 months compared to 30.0 months for those patients with low survivin expression (p = 0.0023) suggesting that survivin expression may provide prognostic information. Similarly patients with advanced gastric cancer with survivin-positive tumors have a significantly lower 5-year survival rate compared to patients with survivin-negative tumors (P < 0.05) [74]. In non-small cell lung cancer (NSCLC), survivin is over-expressed in approximately 80% of tumors and its presence is associated with a reduced survival in patients with resected neoplasms [75, 76].
Expression of survivin also correlates with resistance to therapy. In prostate cancer cells treated with docetaxel, apoptosis is induced by the binding of survivin to SMAC/DIABLO and to the mitotic spindle [77]. Nakahara et al have reported that YM155 (Section 5.1) induced regression of established hormone refractory prostate cancer (HRPC) xenografts [78]. In vitro, docetaxel resistance was reversed in gastric cancer cells when messenger ribonucleic acid (mRNA) expression of survivin was down-regulated by gambogic acid, a survivin inhibitor [43].
Preliminary results are available for a phase II trial investigating 168-hour continuous iv infusion of YM155 (Section 5.1) in patients with metastatic HRPC who have received prior taxane therapy. The primary endpoint of this trial is PSA response rate. Data on 32 patients has been reported and 2 PSA responders were seen indicating YM155 may have some activity in HRPC [79]. An ongoing study is investigating the combination of YM155 and docetaxel in patients with HRPC and preliminary data appear promising. Survivin over-expression has been associated with platinum and taxane-based chemotherapy resistance in a number of solid tumors [80, 81]. It is postulated that vascular endothelial growth factor (VEGF) induced angiogenesis may lead to a transient increase in survivin levels [82] suggesting a role for combining survivin inhibitors with anti-VEGF strategies.
The role of survivin in hematological malignancies
Survivin expression has been observed in the majority of malignant lymphoproliferative disorders with the exception of chronic lymphocytic leukemia in which it is expressed at levels significantly lower than that found in normal lymphocytes [83]. Different techniques for detecting survivin expression have been used including immunohistochemistry (IHC), immunoblotting, and messenger RNA (mRNA) detection; however, no studies comparing these methods have been performed. Three trials have utilized IHC to determine expression of survivin in diffuse large B cell lymphoma (DLBCL) and one study used mRNA expression. The largest study included 222 patients from four separate randomized trials [31]; however, the interpretation of the effect of survivin expression on outcome is complicated by the fact that different chemotherapy regimens were used in the different trials. Although most regimens contained an anthracycline, usually doxorubicin, or mitoxantrone, some regimens did not. In this report, tumors were characterized as being positive for survivin expression if cytoplasmic staining was demonstrated in 70-90% of cells, and negative if less than 5% of the tumor showed staining. Survivin expression was observed in 60% of the patients' tumors and was found to confer an inferior outcome. Patients whose tumors did not show expression of survivin had a 5-year survival of 54%, whereas only 40% of survivin-positive cases were alive at five years (P = 0.02). The complete remission rate was lower in the survivin-expressing group, but not significantly different from the survivin-negative cases at 61% versus 68%, respectively (P = 0.29). This resulted in a lower event-free survival in the survivin-positive cases, but the difference did not achieve statistical significance (p = 0.09). The survival difference was examined in a multivariate analysis incorporating the International Prognostic Index (IPI) and was found to be independent of the IPI score. In two other reports that included 60 and 39 patients, respectively, the nuclear localization of survivin was associated with an inferior outcome in one, but not the other [84, 85]. The relatively small numbers of patients in these studies may have accounted for this disparity. The statistical significance was lost when the patients in the larger study were grouped according to whether they represented a germinal center or non-germinal center DLBCL. There was a trend towards a worse outcome in both subgroups, but the difference did not achieve significance.
Survivin expression has also been evaluated using an RNAse protection assay [86]. Here the authors reported that survivin mRNA expression was increased in 80% of the tumors examined, but it did not impact on overall survival. Survivin protein expression was not simultaneously examined and it is possible that translational regulation could have resulted in a disparity in the mRNA and protein expression levels. A recent report examining the relationship between survivin mRNA and protein levels in patients with HTLV-1-associated adult T cell leukemia/lymphoma (ATLL) found a strong correlation between mRNA levels and protein expression [87].
There has been increasing interest in survivin as a potential therapeutic target in lymphoma. Antisense oligonucleotides (ASO) have been shown to inhibit survivin expression in a number of DLBCL cell lines. These ASO inhibited cell growth in vitro and slowed tumor growth in animals [88].
Clinical studies of investigational survivin inhibitors are ongoing in a number of hematological malignancies. There was great enthusiasm for the small molecule imidazolium-based survivin transcriptional inhibitor YM155 (Section 5.1) following the phase I trial as responses were observed in three of five patients with relapsed B cell lymphoma, including two patients with DLBCL [89]. These results were not reproduced in the phase II trial that included 41 patients with relapsed and refractory DLBCL [90]. At the time of the abstract submission, only one of the first 25 patients enrolled and evaluated for tumor response showed an objective response. YM155 was well tolerated with grade 3 or 4 anemia (16%), neutropenia (8%), fatigue (8%), and deep venous thrombosis (8%) as the only events that occurred in more than 4% of the patients. Due to the potential for synergism of YM155 with other agents, additional trials are underway or planned combining YM155 with chemotherapy or monoclonal antibodies including rituximab and alemtuzumab.
Survivin expression may have even greater prognostic significance in patients with T cell lymphoma [50, 87, 91, 92]. High levels of expression have been reported in patients with HTLV-1-associated ATLL. Survivin levels were greater in the aggressive acute subtype of the disease compared with the more indolent chronic form of ATLL. ATLL patients whose tumors expressed high levels of survivin had an inferior outcome, with a median survival time of 6.4 months compared to 18 months in those patients with low survivin levels [93]. These results have been reproduced in a number of other studies and are similar to the results seen in DLBCL. Inhibition of survivin expression in primary ATL cells using shRNA leads to decreases in tumor cell viability.
Anaplastic large cell lymphoma (ALCL), another relatively uncommon T cell neoplasm, is separated into two prognostic groups based on the expression of the anaplastic large cell kinase gene (ALK). The expression of ALK is most frequently the result of a translocation between chromosomes 2 and 5, resulting in a unique fusion of the ALK and nucleophosmin genes, respectively. ALK-positive ALCL has a superior clinical outcome with about 80% of patients cured with current chemotherapy. In contrast, patients with ALK-negative ALCL demonstrate a worse prognosis with a 50% 5-year survival. Survivin expression was examined in ALCL patients by IHC and was found to be frequently positive with primarily cytoplasmic localization [92]. Approximately 63% of ALK-positive ALCL tumors expressed survivin compared with 47% of the ALK-negative ALCL. Absence of survivin expression in both subtypes conferred an improved prognosis with 100% of the ALK-positive, survivin-negative patients alive at 5 years, and 89% of the ALK-negative, survivin-negative patients alive at 5-years. In contrast only 34% of ALK-positive, survivin-positive patients were alive at 5 years. For the ALK-negative group, the 5-year overall survival was 60% for patients with survivin-positive tumors versus 92% for patients with survivin-negative tumors (P = .04). Survivin expression remained an independent adverse prognostic marker in a multivariate analysis that included IPI score.
Current therapeutic approaches
Transcriptional repressors
YM155 (1-(2-Methoxyethyl)-2-methyl-4,9-dioxo-3-(pyrazin-2-ylmethyl)-4,9-ihydro-H-naphtho[2,3-d]imidazolium bromide) (Astellas Pharma Inc) is a small-molecule suppressor of survivin. YM155 interacts specifically with the 269 base pair survivin core promoter region and functions in a cell cycle independent manner as a transcriptional inhibitor. YM155 has demonstrated potent anti-proliferative activity against various cancer cell lines and preferentially induces cell death in tumor cells. Preclinical data has also shown that it has the ability to serve as a radio-sensitizing agent and to potentiate the antitumor activity of various cytotoxic agents including carboplatin and paclitaxel [94, 95]. In preclinical experiments, YM155 has shown promising activity in a wide variety of human tumor xenograft models including non-small cell lung cancer (NSCLC) [96].
YM155 has been investigated in six phase I or II clinical trials in solid tumors and non-Hodgkins lymphoma. It has been shown to be well tolerated with the most common toxicities being reported as grades 1-2 in severity and consisting primarily of stomatitis, pyrexia and nausea [97–100]. In a phase I study conducted in the United States, 41 patients received YM155 at doses ranging from 1.8 to 6.0 mg/m2/d by a 168-hour (7 days) continuous intravenous infusion every 3 weeks. The maximum tolerated dose (MTD) was determined as 4.8 mg/m2. In a second phase I trial conducted in Japan [101], patients with advanced refractory solid tumors were treated with escalating doses of YM155 administered by continuous i.v. infusion for 168-hours in 21-day cycles. Of the 34 patients enrolled, 33 (median age, 59 years) received at least 1 dose of YM155 and the MTD was determined to be 8.0 mg/m2/d. The most common adverse reactions judged to be related to YM155 were microalbuminuria, fever, injection-site phlebitis, fatigue, and decreased hemoglobin/anemia, serum albumin, and lymphocyte count.
As monotherapy, YM155 has shown modest antitumor activity in a phase II trial in patients with advanced NSCLC who had failed one or two prior chemotherapies [98, 102]. Two partial responses were observed in 37 patients (5.4%) enrolled in this study, where YM155 was administered as a 7 day continuous infusion at 4.8 mg/m2/day, repeated every 3 weeks. Fourteen patients (37.8%) achieved stable disease resulting in a disease control rate of 43.2% (95% CI, 27.1% to 60.5%). Median PFS was 1.7 months (95% CI, 1.3 to 2.8 months). Median overall survival was 6.6 months (95% CI, 4 to 12.2 months), with a 1-year survival rate of 35.1%. Treatment was well tolerated with the majority of treatment discontinuations not YM155 related. A phase I/II study evaluating the combination of YM155 in combination with carboplatin and paclitaxel has now been initiated. YM155 is also being investigated in malignant melanoma, HER-2/neu-negative breast cancer, and in combination with docetaxel in hormone refractory prostate cancer (see Table 2).
Terameprocol (EM-1421, Erimos Pharmaceuticals) is a semi-synthetic small molecule with antitumor activity occurring via selective targeting of Sp1-regulated proteins, including survivin and cyclin dependent kinases (cdc2) that control cell cycle and apoptosis [103, 104]. Terameprocol is in clinical development as a site-specific transcription inhibitor in solid refractory tumors and in leukemias.
mRNA Inhibitors
LY2181308 (ISIS/Eli Lilly Pharmaceuticals), a novel modified ASO, is a specific inhibitor of survivin. In a phase I trial in which 24 patients were enrolled, LY2181308 showed a safety and pharmacokinetic (PK) profile consistent with previously described ASOs. Side effects were mild to moderate and no grade 3 or 4 toxicities were described at the MTD of 750 mg [105]. Additionally, a pharmacodynamic study has been conducted in 34 patients, including 22 patients with available pre- and post-treatment biopsies. IHC indicated that survivin expression was reduced in the nucleus and cytoplasm in 11 of 17, and 5 of 14 evaluable pairs, respectively. Gene expression analysis indicated a reduction in survivin expression of 20-50% in 11 of 15 evaluable pairs. Analysis of fresh tumor from endobronchial sampling revealed that 2 of 3 NSCLC patients had a near-complete elimination of survivin-positive cells accompanied by an increase in the fraction of cells with a sub-G1 DNA content, consistent with cell death [106]. Furthermore, a human micro-dosing imaging PK study of an ASO with LY2181308 using carbon-11 radiolabeled LY2181308 ([11C]LY2181308) was conducted. In this study pharmacokinetic analysis confirmed that biologically active human tumor drug concentrations of [11C]LY2181308 can be achieved. LY2181308 therapy saturated normal tissue kinetics and increased tumor uptake of [11C]LY2181308 [107]. LY2181308 is currently being evaluated in phase II trials for relapsed and refractory acute myeloid leukemia, prostate cancer and NSCLC (see Table 2). Other approaches for inactivating specific mRNAs that remain predominantly in the preclinical setting involve the use of hammerhead ribozymes [108, 109] and small interfering RNAs [110, 111].
Small molecule survivin inhibitors
Shepherdin is a small molecule inhibitor in early stage clinical development that acts as an antagonist of the survivin-Hsp 90 complex [112]. This molecule is a 5 amino acid peptide that antagonizes the binding between survivin and Hsp-90 as well as acting as a global inhibitor of Hsp90 function via competition with ATP.
Immunotherapy
Due to its differential expression by tumors, it has been hypothesized that cancer patients may recognize survivin as a "non-self" protein and mount an immune response against it [113]. Phase I trials using survivin-directed autologous cytotoxic T lymphocytes acitvated with survivin-primed dendritic cells or survivin peptides have been performed. Treatment was well tolerated with limited toxicity [114, 115]. Wobser et al, described an early example of survivin-based vaccination therapy in a patient with metastatic pancreatic cancer that achieved a complete regression of liver metastasis after receiving a HLA-A2 restricted survivin peptide plus adjuvant [116]. Recently a phase I-II trial of vaccination with HLA-A*0201-restricted peptides from prostate specific membrane antigen (PSMA) and survivin was carried out in 20 prostate cancer patients with biochemical failure after surgery or radiotherapy. The vaccine consisted of two peptides from PSMA (PSMA4-12 and PSMA711-719) and one from survivin (SVV96-104/97M) given by 4 biweekly (priming) and then 4 monthly administrations (boosting). To selectively eliminate regulatory T cells (Tregs) and possibly enhance immunization, the vaccinations were preceded by cyclophosphamide 300 mg/kq, i.v. The vaccine was well tolerated, with 14/20 patients exhibiting a significant although transient PSA decrease. Most patients (19/20) showed a significant increase of SVV96-104/97M-specific T cells, while a response to PSMA was achieved in about half of the patients. Increments of HLA-A*0201/SVV96-104/97M or PSMA711-719 multimer+ CD8+ T cells were induced in 50% and 35% of patients, respectively. However, vaccination-induced Ag-specific T cells displayed limited cross-reactivity with HLA-A*0201+ prostate cancer cells. These results indicate that the vaccine-induced Ag-specific CD8+ T cells had a reduced ability to cross-recognize prostate cancer cells which could explain why PSA control was achieved only transiently [117].
The tolerability, immunogenicity, and clinical efficacy of three survivin peptides restricted to HLA A1, A2 and B35 were addressed in a phase I/II trial which included 79 patients with metastatic melanoma (n = 61), pancreatic (n = 8), cervical (n = 5), colorectal (n = 2), adrenal gland (n = 2), and Merkel cell carcinoma (n = 1) who failed to respond to systemic standard therapy. Peptide vaccination was safe and vaccine-specific immune responses were induced in 50% of the patients. Objective responses and control of disease were essentially restricted to immunological responders. Three complete responses and three partial responses were observed (OR = 7.6%) with the duration of response ranging from 3 to 36+ months [118]. Due to these encouraging results, ongoing Phase II studies are investigating survivin based immunotherapy in patients with advanced pancreatic, colon and cervical carcinomas as well as in melanoma.
Survivin as a radiosensitizer
Survivin is known to play an important role in the sensitivity of cells to radiation with high survivin levels corresponding to reduced radiation sensitivity [119, 120]. In a number of preclinical studies inhibition of survivin expression has been shown to sensitize tumor cells to ionizing radiation [121–124]. Although apoptosis does not play a major role in the response of solid tumors to radiation as a single agent, inhibition of survivin prior to irradiation leads to an increase in apoptosis and reduced tumor cell survival [121]. In addition, survivin inhibition interferes with normal cell cycle progression and DNA repair [125, 126]. Consistent with these observations, a number of small molecule inhibitors of survivin have been shown to enhance the cytotoxic effects of radiation [127]. Collectively, these findings provide a strong rationale for the clinical evaluation of survivin inhibitors administered either concurrently or sequentially with therapeutic radiation.
Conclusion
Over the last decade it has become increasingly clear that inhibitor of apoptosis proteins play an integral role in maintaining cellular homeostasis. In particular, one of these proteins, survivin serves many functions involved in cell survival including complex intracellular signaling, stabilizing mitosis and facilitating cellular adaptation. Much remains to be learned regarding the biology of survivin and other IAPs in terms of how these molecules intersect with other pathways. Survivin is a highly expressed in many different tumors and its expression correlates with advanced disease, poorer survival, and chemotherapy and radiation resistance. Because of the role it plays, survivin is of increasing interest as a potential therapeutic target in cancer. Survivin antagonists may function not as single protein inhibitors, but rather as global pathway inhibitors that may disable multiple signaling circuits in tumors. Clinical trials have highlighted the problems with attempts to correlate survivin expression with clinical outcome. Small sample numbers, non-uniform treatments, the presence of multiple alternatively spliced survivin mRNAs with differing effects on apoptosis and the different methods of detection of survivin, all lead to difficulty in trial interpretation. Further efforts are required to achieve a greater understanding of the biology of survivin and the other IAPs and more effectively exploit strategies that target this protein in cancer.
Conflict of interest
The authors declare that they have no competing interests.
Abbreviations
- ALCL:
-
anaplastic large cell lymphoma
- ALK:
-
anaplastic lymphoma kinase
- ATLL:
-
adult T cell leukemia/lymphoma
- HTLV-1:
-
human T cell lymphotrophic virus-1
- IAP:
-
inhibitor of apoptosis protein
- NSCLC:
-
non-small cell lung cancer
- VEGF:
-
vascular endothelial growth factor
- DLBCL:
-
diffuse large B cell lymphoma
- SMAC:
-
second mitochondria-derived activator of caspases
- DIABLO:
-
direct inhibitor of apoptosis binding protein with low pI
- mRNA:
-
messenger ribonucleic acid
- PK:
-
phramacokinetics
- TNF-α:
-
tumor necrosis factor alpha
- HBXIP:
-
hepatitis B X-interacting protein
- XIAP:
-
X-linked IAP
- Hsp90:
-
heat shock protein 90
- INCENP:
-
inner centromere protein antigens
- CDK1:
-
cyclin-dependent kinase 1
- CPC:
-
chromosomal passenger complex.
References
Altieri DC: Survivin, cancer networks and pathway-directed drug discovery. Nat Rev Cancer. 2008, 8: 61-70. 10.1038/nrc2293
Li F, Ambrosini G, Chu EY, Plescia J, Tognin S, Marchisio PC, Altieri DC: Control of apoptosis and mitotic spindle checkpoint by survivin. Nature. 1998, 396: 580-584. 10.1038/25141
Hunter AM, LaCasse EC, Korneluk RG: The inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis. 2007, 12: 1543-1568. 10.1007/s10495-007-0087-3
Schimmer AD: Inhibitor of apoptosis proteins: translating basic knowledge into clinical practice. Cancer Res. 2004, 64: 7183-7190. 10.1158/0008-5472.CAN-04-1918
Ambrosini G, Adida C, Altieri DC: A novel anti-apoptosis gene, survivin, expressed in cancer and lymphoma. Nat Med. 1997, 3: 917-921. 10.1038/nm0897-917
Li F, Altieri DC: The cancer antiapoptosis mouse survivin gene: characterization of locus and transcriptional requirements of basal and cell cycle-dependent expression. Cancer Res. 1999, 59: 3143-3151.
Verdecia MA, Huang H, Dutil E, Kaiser DA, Hunter T, Noel JP: Structure of the human anti-apoptotic protein survivin reveals a dimeric arrangement. Nat Struct Biol. 2000, 7: 602-608. 10.1038/77929
Chantalat L, Skoufias DA, Kleman JP, Jung B, Dideberg O, Margolis RL: Crystal structure of human survivin reveals a bow tie-shaped dimer with two unusual alpha-helical extensions. Mol Cell. 2000, 6: 183-189.
Mahotka C, Wenzel M, Springer E, Gabbert HE, Gerharz CD: Survivin-deltaEx3 and survivin-2B: two novel splice variants of the apoptosis inhibitor survivin with different antiapoptotic properties. Cancer Res. 1999, 59: 6097-6102.
Caldas H, Honsey LE, Altura RA: Survivin 2alpha: a novel Survivin splice variant expressed in human malignancies. Mol Cancer. 2005, 4: 11- 10.1186/1476-4598-4-11
Badran A, Yoshida A, Ishikawa K, Goi T, Yamaguchi A, Ueda T, Inuzuka M: Identification of a novel splice variant of the human anti-apoptopsis gene survivin. Biochem Biophys Res Commun. 2004, 314: 902-907. 10.1016/j.bbrc.2003.12.178
Blanc-Brude OP, Mesri M, Wall NR, Plescia J, Dohi T, Altieri DC: Therapeutic targeting of the survivin pathway in cancer: initiation of mitochondrial apoptosis and suppression of tumor-associated angiogenesis. Clin Cancer Res. 2003, 9: 2683-2692.
Pennati M, Folini M, Zaffaroni N: Targeting survivin in cancer therapy: fulfilled promises and open questions. Carcinogenesis. 2007, 28: 1133-1139. 10.1093/carcin/bgm047
Tamm I, Wang Y, Sausville E, Scudiero DA, Vigna N, Oltersdorf T, Reed JC: IAP-family protein survivin inhibits caspase activity and apoptosis induced by Fas (CD95), Bax, caspases, and anticancer drugs. Cancer Res. 1998, 58: 5315-5320.
Banks DP, Plescia J, Altieri DC, Chen J, Rosenberg SH, Zhang H, Ng SC: Survivin does not inhibit caspase-3 activity. Blood. 2000, 96: 4002-4003.
Li C, Wu Z, Liu M, Pazgier M, Lu W: Chemically synthesized human survivin does not inhibit caspase-3. Protein Sci. 2008, 17: 1624-1629. 10.1110/ps.036145.108
Eckelman BP, Salvesen GS, Scott FL: Human inhibitor of apoptosis proteins: why XIAP is the black sheep of the family. EMBO Rep. 2006, 7: 988-994. 10.1038/sj.embor.7400795
Dohi T, Beltrami E, Wall NR, Plescia J, Altieri DC: Mitochondrial survivin inhibits apoptosis and promotes tumorigenesis. J Clin Invest. 2004, 114: 1117-1127.
Marusawa H, Matsuzawa S, Welsh K, Zou H, Armstrong R, Tamm I, Reed JC: HBXIP functions as a cofactor of survivin in apoptosis suppression. EMBO J. 2003, 22: 2729-2740. 10.1093/emboj/cdg263
Dohi T, Okada K, Xia F, Wilford CE, Samuel T, Welsh K, Marusawa H, Zou H, Armstrong R, Matsuzawa S, Salvesen GS, Reed JC, Altieri DC: An IAP-IAP complex inhibits apoptosis. J Biol Chem. 2004, 279: 34087-34090. 10.1074/jbc.C400236200
Verhagen AM, Ekert PG, Pakusch M, Silke J, Connolly LM, Reid GE, Moritz RL, Simpson RJ, Vaux DL: Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to and antagonizing IAP proteins. Cell. 2000, 102: 43-53. 10.1016/S0092-8674(00)00009-X
Du C, Fang M, Li Y, Li L, Wang X: Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell. 2000, 102: 33-42. 10.1016/S0092-8674(00)00008-8
Yang D, Welm A, Bishop JM: Cell division and cell survival in the absence of survivin. Proc Natl Acad Sci USA. 2004, 101: 15100-15105. 10.1073/pnas.0406665101
Okada H, Mak TW: Pathways of apoptotic and non-apoptotic death in tumour cells. Nat Rev Cancer. 2004, 4: 592-603. 10.1038/nrc1412
Vader G, Kauw JJ, Medema RH, Lens SM: Survivin mediates targeting of the chromosomal passenger complex to the centromere and midbody. EMBO Rep. 2006, 7: 85-92. 10.1038/sj.embor.7400562
Ruchaud S, Carmena M, Earnshaw WC: The chromosomal passenger complex: one for all and all for one. Cell. 2007, 131: 230-231. 10.1016/j.cell.2007.10.002
Li F, Altieri DC: Transcriptional analysis of human survivin gene expression. Biochem J. 1999, 344 (Pt 2): 305-311. 10.1042/0264-6021:3440305
Harfouche R, Hassessian HM, Guo Y, Faivre V, Srikant CB, Yancopoulos GD, Hussain SN: Mechanisms which mediate the antiapoptotic effects of angiopoietin-1 on endothelial cells. Microvasc Res. 2002, 64: 135-147. 10.1006/mvre.2002.2421
Mesri M, Morales-Ruiz M, Ackermann EJ, Bennett CF, Pober JS, Sessa WC, Altieri DC: Suppression of vascular endothelial growth factor-mediated endothelial cell protection by survivin targeting. Am J Pathol. 2001, 158: 1757-1765. 10.1016/S0002-9440(10)64131-4
Kawasaki H, Altieri DC, Lu CD, Toyoda M, Tenjo T, Tanigawa N: Inhibition of apoptosis by survivin predicts shorter survival rates in colorectal cancer. Cancer Res. 1998, 58: 5071-5074.
Adida C, Haioun C, Gaulard P, Lepage E, Morel P, Briere J, Dombret H, Reyes F, Diebold J, Gisselbrecht C, Salles G, Altieri DC, Molina TJ: Prognostic significance of survivin expression in diffuse large B-cell lymphomas. Blood. 2000, 96: 1921-1925.
Takai N, Miyazaki T, Nishida M, Nasu K, Miyakawa I: Survivin expression correlates with clinical stage, histological grade, invasive behavior and survival rate in endometrial carcinoma. Cancer Lett. 2002, 184: 105-116. 10.1016/S0304-3835(02)00190-8
Tanaka K, Iwamoto S, Gon G, Nohara T, Iwamoto M, Tanigawa N: Expression of survivin and its relationship to loss of apoptosis in breast carcinomas. Clin Cancer Res. 2000, 6: 127-134.
Span PN, Sweep FC, Wiegerinck ET, Tjan-Heijnen VC, Manders P, Beex LV, de Kok JB: Survivin is an independent prognostic marker for risk stratification of breast cancer patients. Clin Chem. 2004, 50: 1986-1993. 10.1373/clinchem.2004.039149
Ferrandina G, Legge F, Martinelli E, Ranelletti FO, Zannoni GF, Lauriola L, Gessi M, Gallotta V, Scambia G: Survivin expression in ovarian cancer and its correlation with clinico-pathological, surgical and apoptosis-related parameters. Br J Cancer. 2005, 92: 271-277.
Lee JP, Chang KH, Han JH, Ryu HS: Survivin, a novel anti-apoptosis inhibitor, expression in uterine cervical cancer and relationship with prognostic factors. Int J Gynecol Cancer. 2005, 15: 113-119. 10.1111/j.1048-891X.2005.15011.x
Lu CD, Altieri DC, Tanigawa N: Expression of a novel antiapoptosis gene, survivin, correlated with tumor cell apoptosis and p53 accumulation in gastric carcinomas. Cancer Res. 1998, 58: 1808-1812.
Adida C, Berrebi D, Peuchmaur M, Reyes-Mugica M, Altieri DC: Anti-apoptosis gene, survivin, and prognosis of neuroblastoma. Lancet. 1998, 351: 882-883. 10.1016/S0140-6736(05)70294-4
Islam A, Kageyama H, Takada N, Kawamoto T, Takayasu H, Isogai E, Ohira M, Hashizume K, Kobayashi H, Kaneko Y, Nakagawara A: High expression of Survivin, mapped to 17q25, is significantly associated with poor prognostic factors and promotes cell survival in human neuroblastoma. Oncogene. 2000, 19: 617-623. 10.1038/sj.onc.1203358
Sarela AI, Macadam RC, Farmery SM, Markham AF, Guillou PJ: Expression of the antiapoptosis gene, survivin, predicts death from recurrent colorectal carcinoma. Gut. 2000, 46: 645-650. 10.1136/gut.46.5.645
Vischioni B, van der Valk P, Span SW, Kruyt FA, Rodriguez JA, Giaccone G: Nuclear localization of survivin is a positive prognostic factor for survival in advanced non-small-cell lung cancer. Ann Oncol. 2004, 15: 1654-1660. 10.1093/annonc/mdh436
Zaffaroni N, Daidone MG: Survivin expression and resistance to anticancer treatments: perspectives for new therapeutic interventions. Drug Resist Updat. 2002, 5: 65-72. 10.1016/S1368-7646(02)00049-3
Wang T, Wei J, Qian X, Ding Y, Yu L, Liu B: Gambogic acid, a potent inhibitor of survivin, reverses docetaxel resistance in gastric cancer cells. Cancer Lett. 2008, 262: 214-222. 10.1016/j.canlet.2007.12.004
Meng H, Tanigawa N, Hao CY, Dai DJ, Lu CD, Ji JF: Chemoresponse to docetaxel correlates with expression of the survivin splicing variants in patients with gastric cancer. Hepatogastroenterology. 2007, 54: 1934-1940.
Adams JM, Cory S: Life-or-death decisions by the Bcl-2 protein family. Trends Biochem Sci. 2001, 26: 61-66. 10.1016/S0968-0004(00)01740-0
Cryns V, Yuan J: Proteases to die for. Genes Dev. 1998, 12: 1551-1570. 10.1101/gad.12.11.1551
Mita AC, Mita MM, Nawrocki ST, Giles FJ: Survivin: key regulator of mitosis and apoptosis and novel target for cancer therapeutics. Clin Cancer Res. 2008, 14: 5000-5005. 10.1158/1078-0432.CCR-08-0746
Van Antwerp DJ, Martin SJ, Verma IM, Green DR: Inhibition of TNF-induced apoptosis by NF-kappa B. Trends Cell Biol. 1998, 8: 107-111. 10.1016/S0962-8924(97)01215-4
Kim PJ, Plescia J, Clevers H, Fearon ER, Altieri DC: Survivin and molecular pathogenesis of colorectal cancer. Lancet. 2003, 362: 205-209. 10.1016/S0140-6736(03)13910-4
Pise-Masison CA, Radonovich M, Dohoney K, Morris JC, O'Mahony D, Lee MJ, Trepel J, Waldmann TA, Janik JE, Brady JN: Gene expression profiling of ATL patients: compilation of disease-related genes and evidence for TCF4 involvement in BIRC5 gene expression and cell viability. Blood. 2009, 113: 4016-4026. 10.1182/blood-2008-08-175901
Vaira V, Lee CW, Goel HL, Bosari S, Languino LR, Altieri DC: Regulation of survivin expression by IGF-1/mTOR signaling. Oncogene. 2007, 26: 2678-2684. 10.1038/sj.onc.1210094
Sommer KW, Schamberger CJ, Schmidt GE, Sasgary S, Cerni C: Inhibitor of apoptosis protein (IAP) survivin is upregulated by oncogenic c-H-Ras. Oncogene. 2003, 22: 4266-4280. 10.1038/sj.onc.1206509
You L, He B, Xu Z, Uematsu K, Mazieres J, Mikami I, Reguart N, Moody TW, Kitajewski J, McCormick F, Jablons DM: Inhibition of Wnt-2-mediated signaling induces programmed cell death in non-small-cell lung cancer cells. Oncogene. 2004, 23: 6170-6174. 10.1038/sj.onc.1207844
Fortugno P, Beltrami E, Plescia J, Fontana J, Pradhan D, Marchisio PC, Sessa WC, Altieri DC: Regulation of survivin function by Hsp90. Proc Natl Acad Sci USA. 2003, 100: 13791-13796. 10.1073/pnas.2434345100
O'Connor DS, Grossman D, Plescia J, Li F, Zhang H, Villa A, Tognin S, Marchisio PC, Altieri DC: Regulation of apoptosis at cell division by p34cdc2 phosphorylation of survivin. Proc Natl Acad Sci USA. 2000, 97: 13103-13107.
Kang BH, Altieri DC: Regulation of survivin stability by the aryl hydrocarbon receptor-interacting protein. J Biol Chem. 2006, 281: 24721-24727. 10.1074/jbc.M603175200
Ghosh JC, Dohi T, Kang BH, Altieri DC: Hsp60 regulation of tumor cell apoptosis. J Biol Chem. 2008, 283: 5188-5194. 10.1074/jbc.M705904200
O'Connor DS, Wall NR, Porter AC, Altieri DC: A p34(cdc2) survival checkpoint in cancer. Cancer Cell. 2002, 2: 43-54.
Altieri DC: The case for survivin as a regulator of microtubule dynamics and cell-death decisions. Curr Opin Cell Biol. 2006, 18: 609-615. 10.1016/j.ceb.2006.08.015
Bolton MA, Lan W, Powers SE, McCleland ML, Kuang J, Stukenberg PT: Aurora B kinase exists in a complex with survivin and INCENP and its kinase activity is stimulated by survivin binding and phosphorylation. Mol Biol Cell. 2002, 13: 3064-3077. 10.1091/mbc.E02-02-0092
Gassmann R, Carvalho A, Henzing AJ, Ruchaud S, Hudson DF, Honda R, Nigg EA, Gerloff DL, Earnshaw WC: Borealin: a novel chromosomal passenger required for stability of the bipolar mitotic spindle. J Cell Biol. 2004, 166: 179-191. 10.1083/jcb.200404001
Lens SM, Vader G, Medema RH: The case for Survivin as mitotic regulator. Curr Opin Cell Biol. 2006, 18: 616-622. 10.1016/j.ceb.2006.08.016
Span PN, Tjan-Heijnen VC, Manders P, van Tienoven D, Lehr J, Sweep FC: High survivin predicts a poor response to endocrine therapy, but a good response to chemotherapy in advanced breast cancer. Breast Cancer Res Treat. 2006, 98: 223-230. 10.1007/s10549-005-9153-0
Ding Y, Prieto VG, Zhang PS, Rosenthal S, Smith KJ, Skelton HG, Diwan AH: Nuclear expression of the antiapoptotic protein survivin in malignant melanoma. Cancer. 2006, 106: 1123-1129. 10.1002/cncr.21727
Piras F, Murtas D, Minerba L, Ugalde J, Floris C, Maxia C, Colombari R, Perra MT, Sirigu P: Nuclear survivin is associated with disease recurrence and poor survival in patients with cutaneous malignant melanoma. Histopathology. 2007, 50: 835-842. 10.1111/j.1365-2559.2007.02695.x
Chen N, Gong J, Chen X, Meng W, Huang Y, Zhao F, Wang L, Zhou Q: Caspases and inhibitor of apoptosis proteins in cutaneous and mucosal melanoma: expression profile and clinicopathologic significance. Hum Pathol. 2009, 40: 950-956. 10.1016/j.humpath.2008.12.001
Nasr MR, El-Zammar O: Comparison of pHH3, Ki-67, and survivin immunoreactivity in benign and malignant melanocytic lesions. Am J Dermatopathol. 2008, 30: 117-122. 10.1097/DAD.0b013e3181624054
Gradilone A, Gazzaniga P, Ribuffo D, Scarpa S, Cigna E, Vasaturo F, Bottoni U, Innocenzi D, Calvieri S, Scuderi N, Frati L, Agliano AM: Survivin, bcl-2, bax, and bcl-X gene expression in sentinel lymph nodes from melanoma patients. J Clin Oncol. 2003, 21: 306-312. 10.1200/JCO.2003.08.066
Ben Simon GJ, Abulafia A, Pe'er J: Apoptosis inhibitor, survivin, in posterior uveal melanoma: comparison among primary tumors, tumors resistant to brachytherapy, tumors with liver metastases, and liver metastases. Curr Eye Res. 2006, 31: 251-257. 10.1080/02713680600557014
Mamori S, Kahara F, Ohnishi K, Takeda A, Higashida A, Ashida C, Yamada H: Survivin expression in primary malignant melanoma of the esophagus. Scand J Gastroenterol. 2009, 44: 1497-1498. 10.3109/00365520903330310
Altieri DC: Survivin, versatile modulation of cell division and apoptosis in cancer. Oncogene. 2003, 22: 8581-8589. 10.1038/sj.onc.1207113
Kawasaki H, Toyoda M, Shinohara H, Okuda J, Watanabe I, Yamamoto T, Tanaka K, Tenjo T, Tanigawa N: Expression of survivin correlates with apoptosis, proliferation, and angiogenesis during human colorectal tumorigenesis. Cancer. 2001, 91: 2026-2032. 10.1002/1097-0142(20010601)91:11<2026::AID-CNCR1228>3.0.CO;2-E
Kato J, Kuwabara Y, Mitani M, Shinoda N, Sato A, Toyama T, Mitsui A, Nishiwaki T, Moriyama S, Kudo J, Fujii Y: Expression of survivin in esophageal cancer: correlation with the prognosis and response to chemotherapy. Int J Cancer. 2001, 95: 92-95. 10.1002/1097-0215(20010320)95:2<92::AID-IJC1016>3.0.CO;2-9
Song KY, Jung CK, Park WS, Park CH: Expression of the antiapoptosis gene Survivin predicts poor prognosis of stage III gastric adenocarcinoma. Jpn J Clin Oncol. 2009, 39: 290-296. 10.1093/jjco/hyp020
Shinohara ET, Gonzalez A, Massion PP, Chen H, Li M, Freyer AS, Olson SJ, Andersen JJ, Shyr Y, Carbone DP, Johnson DH, Hallahan DE, Lu B: Nuclear survivin predicts recurrence and poor survival in patients with resected nonsmall cell lung carcinoma. Cancer. 2005, 103: 1685-1692. 10.1002/cncr.20951
Monzo M, Rosell R, Felip E, Astudillo J, Sanchez JJ, Maestre J, Martin C, Font A, Barnadas A, Abad A: A novel anti-apoptosis gene: Re-expression of survivin messenger RNA as a prognosis marker in non-small-cell lung cancers. J Clin Oncol. 1999, 17: 2100-2104.
Kim JY, Chung JY, Lee SG, Kim YJ, Park JE, Yoo KS, Yoo YH, Park YC, Kim BG, Kim JM: Nuclear interaction of Smac/DIABLO with Survivin at G2/M arrest prompts docetaxel-induced apoptosis in DU145 prostate cancer cells. Biochem Biophys Res Commun. 2006, 350: 949-954. 10.1016/j.bbrc.2006.09.143
Nakahara T, Takeuchi M, Kinoyama I, Minematsu T, Shirasuna K, Matsuhisa A, Kita A, Tominaga F, Yamanaka K, Kudoh M, Sasamata M: YM155, a novel small-molecule survivin suppressant, induces regression of established human hormone-refractory prostate tumor xenografts. Cancer Res. 2007, 67: 8014-8021. 10.1158/0008-5472.CAN-07-1343
Karavasilis V, Mita A, Hudes G, Quinn D, Ferrari A, Kocak I, Keating A, Bartes P, Charleston D, de Bono J: Phase II monotherapy study of YM155, a novel survivin suppressant, administered by 168-hour continuous infusion in previously treated hormone refractory prostate cancer (HRPC). J Clin Oncol (Meeting Abstracts). 2007, 25: 5135-
Nomura T, Yamasaki M, Nomura Y, Mimata H: Expression of the inhibitors of apoptosis proteins in cisplatin-resistant prostate cancer cells. Oncol Rep. 2005, 14: 993-997.
Zaffaroni N, Pennati M, Colella G, Perego P, Supino R, Gatti L, Pilotti S, Zunino F, Daidone MG: Expression of the anti-apoptotic gene survivin correlates with taxol resistance in human ovarian cancer. Cell Mol Life Sci. 2002, 59: 1406-1412. 10.1007/s00018-002-8518-3
Tran J, Master Z, Yu JL, Rak J, Dumont DJ, Kerbel RS: A role for survivin in chemoresistance of endothelial cells mediated by VEGF. Proc Natl Acad Sci USA. 2002, 99: 4349-4354. 10.1073/pnas.072586399
de Graaf AO, van Krieken JH, Tonnissen E, Wissink W, van de Locht L, Overes I, Dolstra H, de Witte T, van der Reijden BA, Jansen JH: Expression of C-IAP1, C-IAP2 and SURVIVIN discriminates different types of lymphoid malignancies. Br J Haematol. 2005, 130: 852-859. 10.1111/j.1365-2141.2005.05690.x
Watanuki-Miyauchi R, Kojima Y, Tsurumi H, Hara T, Goto N, Kasahara S, Saio M, Moriwaki H, Takami T: Expression of survivin and of antigen detected by a novel monoclonal antibody, T332, is associated with outcome of diffuse large B-cell lymphoma and its subtypes. Pathol Int. 2005, 55: 324-330. 10.1111/j.1440-1827.2005.01832.x
Liu L, Zhang M, Zou P: Expression of PLK1 and survivin in diffuse large B-cell lymphoma. Leuk Lymphoma. 2007, 48: 2179-2183. 10.1080/10428190701615918
Kuttler F, Valnet-Rabier MB, Angonin R, Ferrand C, Deconinck E, Mougin C, Cahn JY, Fest T: Relationship between expression of genes involved in cell cycle control and apoptosis in diffuse large B cell lymphoma: a preferential survivin-cyclin B link. Leukemia. 2002, 16: 726-735. 10.1038/sj.leu.2402427
Che XF, Zheng CL, Owatari S, Mutoh M, Gotanda T, Jeung HC, Furukawa T, Ikeda R, Yamamoto M, Haraguchi M, Arima N, Akiyama S: Overexpression of survivin in primary ATL cells and sodium arsenite induces apoptosis by down-regulating survivin expression in ATL cell lines. Blood. 2006, 107: 4880-4887. 10.1182/blood-2005-08-3423
Ansell SM, Arendt BK, Grote DM, Jelinek DF, Novak AJ, Wellik LE, Remstein ED, Bennett CF, Fielding A: Inhibition of survivin expression suppresses the growth of aggressive non-Hodgkin's lymphoma. Leukemia. 2004, 18: 616-623. 10.1038/sj.leu.2403281
Tolcher AW, Mita A, Lewis LD, Garrett CR, Till E, Daud AI, Patnaik A, Papadopoulos K, Takimoto C, Bartels P, Keating A, Antonia S: Phase I and pharmacokinetic study of YM155, a small-molecule inhibitor of survivin. J Clin Oncol. 2008, 26: 5198-5203. 10.1200/JCO.2008.17.2064
Cheson BD, Vose JM, Bartlett NL, Lopez A, Van der Jagt RH, Tolcher AW, Weisenburger DD, Seiz AL, Shamsili S, Keating AT: Safety and efficacy of YM155 in diffuse large B-cell lymphoma (DLBCL). J Clin Oncol (Meeting Abstracts). 2009, 27: 8502-
Xing Z, Conway EM, Kang C, Winoto A: Essential role of survivin, an inhibitor of apoptosis protein, in T cell development, maturation, and homeostasis. J Exp Med. 2004, 199: 69-80. 10.1084/jem.20031588
Schlette EJ, Medeiros LJ, Goy A, Lai R, Rassidakis GZ: Survivin expression predicts poorer prognosis in anaplastic large-cell lymphoma. J Clin Oncol. 2004, 22: 1682-1688. 10.1200/JCO.2004.10.172
Kamihira S, Yamada Y, Hirakata Y, Tomonaga M, Sugahara K, Hayashi T, Dateki N, Harasawa H, Nakayama K: Aberrant expression of caspase cascade regulatory genes in adult T-cell leukaemia: survivin is an important determinant for prognosis. Br J Haematol. 2001, 114: 63-69. 10.1046/j.1365-2141.2001.02902.x
Iwasa T, Okamoto I, Suzuki M, Nakahara T, Yamanaka K, Hatashita E, Yamada Y, Fukuoka M, Ono K, Nakagawa K: Radiosensitizing Effect of YM155, a Novel Small-Molecule Survivin Suppressant, in Non-Small Cell Lung Cancer Cell Lines. Clin. Cancer Res. 2008, 14: 6496-6504. 10.1158/1078-0432.CCR-08-0468. 10.1158/1078-0432.CCR-08-0468
Yamanaka K, Nakahara T, Kita A, Takeuchi M, Kinoyama I, Matsuhisa A, Koutoku H, Sasamata M: Concomitant and sequential administration of YM155 and docetaxel enhances apoptosis and tumor regression in human tumor xenograft models. Molecular Cancer. Therapeutics. 2007, 6: 3520S-3521S.
Yamanaka K, Nakahara T, Kita A, Miyoshi S, Noda A, Takeuchi M, Kinoyama I, Koutoku H, Nishimura S, Sasamata M: Therapeutic potential of YM155 alone and in combination with chemotherapeutics against human non-small cell lung cancer in carcinoma xenograft models, in, Pergamon-Elsevier Science Ltd. 2008, 112-112.
Alexandrescu DT, Gonzales R, Lewis K, Samlowski W, Cranmer L, Catlett J, Kirkwood J, Whitman E, Lawson D, Bartels P, Drake T, Keating A: A phase II study of YM155 administered as 168 hour continuous infusion in stage IV and unresectable stage III melanoma. Molecular Cancer Therapeutics. 2007, 6: 3385S-3385S.
Giaccone G, Zatloukal P, Roubec J, Musil J, Kuta M, van Klaveren RJ, Chaudhary S, Gunther A, Shamsili S, Kazei D, Verbeeck F: A phase II, monotherapy study of YM155, a novel survivin suppressant, in previously treated advanced stage non-small cell lung cancer (NSCLC). Molecular Cancer Therapeutics. 2007, 6: 3597S-3597S.
Papadopoulos K, Mita A, Antonio S, Lewis LD, Reddy NJ, Till E, Mahany JJ, Keating A, Seiz A, Bartels P, Tolcher A: Efficacy and safety of YM155 in relapsed/refractory non-Hodgkin's lymphoma patients enrolled in a phase I study. Blood. 2007, 110: 3422-
Tolcher AW, Mita A, Lewis LD, Garrett CR, Till E, Daud AI, Patnaik A, Papadopoulos K, Takimoto C, Bartels P, Keating A, Antonia S: Phase I and Pharmacokinetic Study of YM155, a Small-Molecule Inhibitor of Survivin. J Clin Oncol. 2008, 26: 5198-5203. 10.1200/JCO.2008.17.2064
Satoh T, Okamoto I, Miyazaki M, Morinaga R, Tsuya A, Hasegawa Y, Terashima M, Ueda S, Fukuoka M, Ariyoshi Y, Saito T, Masuda N, Watanabe H, Taguchi T, Kakihara T, Aoyama Y, Hashimoto Y, Nakagawa K: Phase I study of YM155, a novel survivin suppressant, in patients with advanced solid tumors. Clin Cancer Res. 2009, 15: 3872-3880. 10.1158/1078-0432.CCR-08-1946
Giaccone G, Zatloukal P, Roubec J, Floor K, Musil J, Kuta M, van Klaveren RJ, Chaudhary S, Gunther A, Shamsili S: Multicenter phase II trial of YM155, a small-molecule suppressor of survivin, in patients with advanced, refractory, non-small-cell lung cancer. J Clin Oncol. 2009, 27: 4481-4486. 10.1200/JCO.2008.21.1862
Chang CC, Heller JD, Kuo J, Huang RC: Tetra-O-methyl nordihydroguaiaretic acid induces growth arrest and cellular apoptosis by inhibiting Cdc2 and survivin expression. Proc Natl Acad Sci USA. 2004, 101: 13239-13244. 10.1073/pnas.0405407101
Smolewski P: Terameprocol, a novel site-specific transcription inhibitor with anticancer activity. IDrugs. 2008, 11: 204-214.
Talbot DC, Davies J, Callies S, Andre V, Lahn M, Ang J, De Bono JS, Ranson M: First human dose study evaluating safety and pharmacokinetics of LY2181308, an antisense oligonucleotide designed to inhibit survivin. J Clin Oncol (Meeting Abstracts). 2008, 26: 3518-
Talbot DC, Davies J, Olsen A, Andre V, Lahn M, Powell E, Kadam S, de Bono J, McHugh P, Ranson M: Pharmacodynamic (PD) evaluation of LYPharmacodynamic (PD) evaluation of LY2181308 in patients with metastatic malignancies. J Clin Oncol (Meeting Abstracts). 2009, 27: 3507-
Saleem A, Ranson M, Callies S, Lahn M, Prenant C, Brown G, Matthews JC, Dence CS, McMahon A, Price P: Microdosing imaging pharmacokinetic (PK) study of the antisense oligonucleotide (ASO) to survivin (LY2181308) using positron emission tomography (PET): A novel paradigm in clinical drug development. J Clin Oncol (Meeting Abstracts). 2009, 27: 3578-
Pennati M, Binda M, Colella G, Zoppe M, Folini M, Vignati S, Valentini A, Citti L, De Cesare M, Pratesi G, Giacca M, Daidone MG, Zaffaroni N: Ribozyme-mediated inhibition of survivin expression increases spontaneous and drug-induced apoptosis and decreases the tumorigenic potential of human prostate cancer cells. Oncogene. 2004, 23: 386-394. 10.1038/sj.onc.1207071
Liu H, Guo S, Roll R, Li J, Diao Z, Shao N, Riley MR, Cole AM, Robinson JP, Snead NM, Shen G, Guo P: Phi29 pRNA vector for efficient escort of hammerhead ribozyme targeting survivin in multiple cancer cells. Cancer Biol Ther. 2007, 6: 697-704. 10.4161/cbt.6.5.3962
Ling X, Li F: Silencing of antiapoptotic survivin gene by multiple approaches of RNA interference technology. Biotechniques. 2004, 36: 450-454. 456-460
Paduano F, Villa R, Pennati M, Folini M, Binda M, Daidone MG, Zaffaroni N: Silencing of survivin gene by small interfering RNAs produces supra-additive growth suppression in combination with 17-allylamino-17-demethoxygeldanamycin in human prostate cancer cells. Mol Cancer Ther. 2006, 5: 179-186. 10.1158/1535-7163.MCT-05-0132
Plescia J, Salz W, Xia F, Pennati M, Zaffaroni N, Daidone MG, Meli M, Dohi T, Fortugno P, Nefedova Y, Gabrilovich DI, Colombo G, Altieri DC: Rational design of shepherdin, a novel anticancer agent. Cancer Cell. 2005, 7: 457-468. 10.1016/j.ccr.2005.03.035
Andersen MH, thor SP: Survivin--a universal tumor antigen. Histol Histopathol. 2002, 17: 669-675.
Otto K, Andersen MH, Eggert A, Keikavoussi P, Pedersen LO, Rath JC, Bock M, Brocker EB, Straten PT, Kampgen E, Becker JC: Lack of toxicity of therapy-induced T cell responses against the universal tumour antigen survivin. Vaccine. 2005, 23: 884-889. 10.1016/j.vaccine.2004.08.007
Tsuruma T, Hata F, Torigoe T, Furuhata T, Idenoue S, Kurotaki T, Yamamoto M, Yagihashi A, Ohmura T, Yamaguchi K, Katsuramaki T, Yasoshima T, Sasaki K, Mizushima Y, Minamida H, Kimura H, Akiyama M, Hirohashi Y, Asanuma H, Tamura Y, Shimozawa K, Sato N, Hirata K: Phase I clinical study of anti-apoptosis protein, survivin-derived peptide vaccine therapy for patients with advanced or recurrent colorectal cancer. J Transl Med. 2004, 2: 19- 10.1186/1479-5876-2-19
Wobser M, Keikavoussi P, Kunzmann V, Weininger M, Andersen MH, Becker JC: Complete remission of liver metastasis of pancreatic cancer under vaccination with a HLA-A2 restricted peptide derived from the universal tumor antigen survivin. Cancer Immunol Immunother. 2006, 55: 1294-1298. 10.1007/s00262-005-0102-x
Valdagni R, Marrari A, Squarcina P, Villa S, Filipazzi P, Salvioni R, Rancati T, Asioli M, Parmiani G, Rivoltini L: Vaccination with survivin and PSMA-derived peptides for controlling biochemical recurrence in prostate cancer: A pilot study. J Clin Oncol (Meeting Abstracts). 2009, 27: e16042-
Becker JC, Wobser M, Hofmeister V, Bauer B, Broecker EB, thor Straten P, Andersen MH: Safety, immunogenicity, and clinical response of a survivin-based peptide vaccine in therapy-resistant advanced cancer: Results from a phase I/II trial. J Clin Oncol (Meeting Abstracts). 2008, 26: 3046-
Asanuma K, Moriai R, Yajima T, Yagihashi A, Yamada M, Kobayashi D, Watanabe N: Survivin as a radioresistance factor in pancreatic cancer. Jpn J Cancer Res. 2000, 91: 1204-1209.
Rodel C, Haas J, Groth A, Grabenbauer GG, Sauer R, Rodel F: Spontaneous and radiation-induced apoptosis in colorectal carcinoma cells with different intrinsic radiosensitivities: survivin as a radioresistance factor. Int J Radiat Oncol Biol Phys. 2003, 55: 1341-1347. 10.1016/S0360-3016(02)04618-7
Lu B, Mu Y, Cao C, Zeng F, Schneider S, Tan J, Price J, Chen J, Freeman M, Hallahan DE: Survivin as a therapeutic target for radiation sensitization in lung cancer. Cancer Res. 2004, 64: 2840-2845. 10.1158/0008-5472.CAN-03-3547
Kappler M, Taubert H, Bartel F, Blumke K, Panian M, Schmidt H, Dunst J, Bache M: Radiosensitization, after a combined treatment of survivin siRNA and irradiation, is correlated with the activation of caspases 3 and 7 in a wt-p53 sarcoma cell line, but not in a mt-p53 sarcoma cell line. Oncol Rep. 2005, 13: 167-172.
Cao C, Mu Y, Hallahan DE, Lu B: XIAP and survivin as therapeutic targets for radiation sensitization in preclinical models of lung cancer. Oncogene. 2004, 23: 7047-7052. 10.1038/sj.onc.1207929
Pennati M, Binda M, Colella G, Folini M, Citti L, Villa R, Daidone MG, Zaffaroni N: Radiosensitization of human melanoma cells by ribozyme-mediated inhibition of survivin expression. J Invest Dermatol. 2003, 120: 648-654. 10.1046/j.1523-1747.2003.12082.x
Chakravarti A, Zhai GG, Zhang M, Malhotra R, Latham DE, Delaney MA, Robe P, Nestler U, Song Q, Loeffler J: Survivin enhances radiation resistance in primary human glioblastoma cells via caspase-independent mechanisms. Oncogene. 2004, 23: 7494-7506. 10.1038/sj.onc.1208049
Rodel F, Hoffmann J, Distel L, Herrmann M, Noisternig T, Papadopoulos T, Sauer R, Rodel C: Survivin as a radioresistance factor, and prognostic and therapeutic target for radiotherapy in rectal cancer. Cancer Res. 2005, 65: 4881-4887. 10.1158/0008-5472.CAN-04-3028
Iwasa T, Okamoto I, Suzuki M, Nakahara T, Yamanaka K, Hatashita E, Yamada Y, Fukuoka M, Ono K, Nakagawa K: Radiosensitizing effect of YM155, a novel small-molecule survivin suppressant, in non-small cell lung cancer cell lines. Clin Cancer Res. 2008, 14: 6496-6504. 10.1158/1078-0432.CCR-08-0468
Grabowski P, Kuhnel T, Muhr-Wilkenshoff F, Heine B, Stein H, Hopfner M, Germer CT, Scherubl H: Prognostic value of nuclear survivin expression in oesophageal squamous cell carcinoma. Br J Cancer. 2003, 88: 115-119. 10.1038/sj.bjc.6600696
Nasu S, Yagihashi A, Izawa A, Saito K, Asanuma K, Nakamura M, Kobayashi D, Okazaki M, Watanabe N: Survivin mRNA expression in patients with breast cancer. Anticancer Res. 2002, 22: 1839-1843.
Satoh K, Kaneko K, Hirota M, Masamune A, Satoh A, Shimosegawa T: Expression of survivin is correlated with cancer cell apoptosis and is involved in the development of human pancreatic duct cell tumors. Cancer. 2001, 92: 271-278. 10.1002/1097-0142(20010715)92:2<271::AID-CNCR1319>3.0.CO;2-0
Sarela AI, Verbeke CS, Ramsdale J, Davies CL, Markham AF, Guillou PJ: Expression of survivin, a novel inhibitor of apoptosis and cell cycle regulatory protein, in pancreatic adenocarcinoma. Br J Cancer. 2002, 86: 886-892. 10.1038/sj.bjc.6600133
Cohen C, Lohmann CM, Cotsonis G, Lawson D, Santoianni R: Survivin expression in ovarian carcinoma: correlation with apoptotic markers and prognosis. Mod Pathol. 2003, 16: 574-583. 10.1097/01.MP.0000073868.31297.B0
Ikeguchi M, Ueda T, Sakatani T, Hirooka Y, Kaibara N: Expression of survivin messenger RNA correlates with poor prognosis in patients with hepatocellular carcinoma. Diagn Mol Pathol. 2002, 11: 33-40. 10.1097/00019606-200203000-00007
Ito T, Shiraki K, Sugimoto K, Yamanaka T, Fujikawa K, Ito M, Takase K, Moriyama M, Kawano H, Hayashida M, Nakano T, Suzuki A: Survivin promotes cell proliferation in human hepatocellular carcinoma. Hepatology. 2000, 31: 1080-1085. 10.1053/he.2000.6496
Yu J, Leung WK, Ebert MP, Ng EK, Go MY, Wang HB, Chung SC, Malfertheiner P, Sung JJ: Increased expression of survivin in gastric cancer patients and in first degree relatives. Br J Cancer. 2002, 87: 91-97. 10.1038/sj.bjc.6600421
Lehner R, Lucia MS, Jarboe EA, Orlicky D, Shroyer AL, McGregor JA, Shroyer KR: Immunohistochemical localization of the IAP protein survivin in bladder mucosa and transitional cell carcinoma. Appl Immunohistochem Mol Morphol. 2002, 10: 134-138. 10.1097/00022744-200206000-00007
Mori A, Wada H, Nishimura Y, Okamoto T, Takemoto Y, Kakishita E: Expression of the antiapoptosis gene survivin in human leukemia. Int J Hematol. 2002, 75: 161-165. 10.1007/BF02982021
Acknowledgements
Funding
This research was supported in part by the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
RK: Conception and design, manuscript writing, final approval of manuscript
AL: Manuscript writing, final approval of manuscript
DC: Manuscript writing, final approval of manuscript
JJ: Manuscript writing, final approval of manuscript
JM: Conception and design, manuscript writing, final approval of manuscript
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Kelly, R.J., Lopez-Chavez, A., Citrin, D. et al. Impacting tumor cell-fate by targeting the inhibitor of apoptosis protein survivin. Mol Cancer 10, 35 (2011). https://doi.org/10.1186/1476-4598-10-35
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1476-4598-10-35