
Abstract
The protozoan Plasmodium falciparum has a complex life cycle in which asexual multiplication in the vertebrate host alternates with an obligate sexual reproduction in the anopheline mosquito. Apart from the apparent recombination advantages conferred by sex, P. falciparum has evolved a remarkable biology and adaptive phenotypes to insure its transmission despite the dangers of sex. This review mainly focuses on the current knowledge on commitment to sexual development, gametocytogenesis and the evolutionary significance of various aspects of gametocyte biology. It goes further than pure biology to look at the strategies used to improve successful transmission. Although gametocytes are inevitable stages for transmission and provide a potential target to fight malaria, they have received less attention than the pathogenic asexual stages. There is a need for research on gametocytes, which are a fascinating stage, responsible to a large extent for the success of P. falciparum.
Similar content being viewed by others
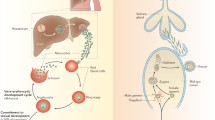

Introduction
Plasmodium falciparum has the morbid characteristic of being the deadliest protozoan parasite of humans. Like all malaria parasites, it is an organism with an obligatory sexual reproduction which takes place in the mosquito midgut. After several development stages, the parasite migrates to the salivary glands to be injected into the next human host. However, before it can succeed sexually in the mosquito host, P. falciparum undergoes a puberty-like process in the human blood: an asexual parasite goes through a series of changes, which will lead to the generation of a sexually competent parasite. This maturation has been termed gametocytogenesis, whereby male and female gametocytes (i.e. pre-gametes) are produced to later fertilize in the invertebrate host.
One of the intriguing facts about sex is that it has been established and maintained regardless of how expensive it might be to the organism bearing it [1]. Several costs are associated with sexual reproduction. There is a two-fold disadvantage as compared to asexual proliferation: the investment in securing a mate and the risk of mixing genes with another, possibly burdened, individual [2]. The fusion of genomes (syngamy), followed by meiosis, whereby chromosomal segregation and recombination occur, seems, however, to confer sufficiently powerful advantages to have driven and maintained sexual reproduction in a vast majority of eukaryotes.
Gametocytogenesis delivers a gametocyte, the only transmission stage from the human to the mosquito. Moreover, it allows sexual achievement and therefore recombination with other genotypes. The present review focuses on specific points of commitment to sexual development – gametocytogenesis and gametocyte biology–especially those relevant to transmission and evolution of P. falciparum transmission strategies.
The biology of gametocytogenesis
A gametocyte is a cell specializing in the transition between the human and the mosquito host. In order to adjust to life in such drastically different environments, many changes occur in its cell biology, metabolism, gene expression and protein synthesis.
Facts and figures on gametocytogenesis
Gametocytaemia, i.e. the presence of gametocytes in the peripheral blood, arises 7–15 days after the initial asexual wave [3, 4]; this maturation period has long been compared to that of the other human malaria species (1–3 days) [5]. It is well established that the ratio of gametocytes to asexual stages in P. falciparum is less than 1:10 [6–9]; a recent study calculates a much lower ratio (1:156) [3]. The half-life of the mature gametocyte in the blood is generally reported to be 2.4 days, based on the observations of Smalley and Sinden [10]. Eichner and colleagues [3] have, however, reported a mean circulation time of 6.4 days which is about twice the expected 3.4 days deduced from a 2.4 half-life. Some gametocytes have been found to have a longevity of up to four weeks in the bloodstream [10].
From merozoite to gametocyte
Gametocytes arise from erythrocytic asexual stages. The production of gametocytes directly from hepatic merozoites, which has been described in other species, does not occur in P. falciparum [11]. There has been much debate on the actual point of sexual differentiation and Bruce and colleagues [12] have shown that merozoites emerging from a single schizont developed either into further asexual stages or into gametocytes. It has been further shown that the gametocytes from one schizont are all male or all female [13, 14]. This suggests that the trophozoites of the preceding asexual generation were already committed to either sexual development or continuing asexual cycling.
Morphology of gametocytes
Field and Shute [15] first described five different maturation stages of P. faciparum gametocytes. These different steps were further characterized by light microscopy using blood from P. falciparum- infected Aotus monkeys [16] and later by electron microscopy [17, 18]. Stages I to V are described in Table 1. One of the most striking feature of gametocytes is the presence of a pellicular complex, which originates from a small membranous vesicle observed beneath the gametocyte plasmalemma in late stage I. This structure is absent from asexual stages. It consists of a subpellicular membrane vacuole subtended by an array of longitudinally-oriented microtubules [8], which strengthens the parasite, explaining the lack of amoeboid forms observed in asexual parasites [17]. The function of this structure is still unknown.
Gene expression in gametocytogenesis
Gametocytes of P. falciparum have been shown to exhibit a different pattern of gene expression than asexual stages, which is unsurprising if one considers the difference of fate between these two stages. Transcription and translation levels are not constant during gametocytogenesis: this was shown in drug sensitivity studies where RNA and protein synthesis levels were much more important in the early than the late gametocyte stages [8]. Furthermore, a sex-specific expression has also been discovered, with differences in RNA, mitochondria and ribosome content. The female is preparing for a continued development, whilst the male is terminally differentiated and only needs what is necessary for exflagellation (e.g. cell division cycle, dynein and α-tubulin II).

Mature female (F) and male (M) gametocytes and trophozoites (T) of Plasmodium falciparum in the blood of malaria-infected patient. This picture is a composite of several pictures originating from the same Giemsa-stained thin smear.
Many stage-specific RNAs and proteins have been described and have been reviewed elsewhere [19]. Most studies have concentrated on surface antigens, the majority of these characterized antigens are gamete antigens that are synthesized during gametocytogenesis. Some are released into the human host circulation in large numbers from dead gametocytes that were not ingested by mosquitoes, hence the immune response observed against some of these gamete surface antigens [20]. With the full genome sequence available, the functional characterization of individual open reading frames (ORFs) offers new insights into the biology of gametocytogenesis. Differential transcription and proteomic studies using different techniques have already been performed. The developmentally differential expression of distinct ribosomal RNA isoforms was highlighted in P. falciparum with at least two rDNA transcription units: one expressed predominantly in sporozoites (S-genes) and one in the asexual cycle (A-genes). Interestingly, the transition between the A and S expression starts during gametocytogenesis [21, 22].
The large scale proteomic study carried out by Lasonder et al. [23] has identified 1,289 malaria proteins of which 315 were solely expressed in the gametocyte, 103 were shared with trophozoite and schizont stages, 163 were shared with gametes and 350 were common to all stages (trophozoites, schizonts, gametes and gametocytes). Of these specific expressed proteins, many were found to have functions as diverse as mRNA processing, ribosomal proteins, cell cycle-DNA processing, energy metabolism [24] and cytoskeletal structure [25]. The number and variety of the gametocyte specific protein reflects the drastic change for which gametocytes are preparing. The way in which the transcription and translation patterns shift from asexual cycling to gametocytogenesis must be exceptionally well adapted to the shift from one host to another.
On the side of the crucial identification of transcripts and protein, it is also important to unravel the regulation processes in gene expression. Given the rich A+T content of the genome [26], it has been tricky to identify promoter sequences. Transcription regulation is thought to be monocistronic. Proteomic analyses have failed to detect a stage-specific chromosomal clustering of gene expression [24]. Accordingly, the differential transcriptome analysis performed for asexual blood stages has corroborated the hypothesis of polycistronic regulation in P. falciparum [27]. Although a common regulation for stage-specific groups of genes is attractive, a full transcriptome analysis of both asexual and sexual blood stages is needed to gain a clearer picture of the differential expression patterns.
Are gametocytes diploid?
DNA synthesis was shown to take place at the very beginning of gametocytogenesis, between the sexually committed ring form and the stage I gametocyte [28, 29]; from then on the DNA content of gametocytes is about twice the haploid amount. The gametocytes of Plasmodium berghei also have more DNA than the haploid content, but significantly less than the diploid amount [30]. This observation, together with the fact that genome segregation failed to be detected with Feulgen staining, indicates that the excess of DNA in P. falciparum may not be due to a duplication of the genome. The macrogametocyte has not been shown to be diploid, and the microgametocyte is certainly not octoploid, in contrast to that of P. berghei [30]. The triple duplication of the genome occurs only after activation of the microgametocyte [28]. Finally, the supplementary DNA may be accounted for by selective gene amplification. Sinden suggested that this DNA may represent amplified ribosomal RNA genes located in the nucleolus [8], as the gametocyte prepares for the expression of the S form in the mosquito stage [31]. Although rRNA could not be detected in the nucleolus of P. berghei by in situ hybridization [32], the concept of selective gene amplification pre-synthesis is an attractive hypothesis in view of the transient role of gametocytes and needs to be further explored.
Metabolic changes in sexual development
P. falciparum gametocytes switch from one environment to another and this must involve the parasite in considerable metabolic changes considering the major differences between the two microenvironments. The rich milieu of the blood is modified by mosquito factors (such as serine proteases and chymotrypsins) [33], there is a drop in temperature, a modified pH and, importantly, the parasite becomes extracellular from then on.
In the intra-erythrocytic asexual stage, energy is mainly produced by anaerobic glycolytic ATP production in the Embden-Meyerhoff-Parnas pathway [34]. It is presumed that glycolytic enzymes are also present and active in gametocyte and mosquito stages, but no evidence has supported this assumption so far, except that the mosquito stages of Plasmodium yoelii express lactate dehydrogenase in Anopheles Stephensi and Anopheles gambiae [35].
The mitochondria of asexual parasites and early-gametocytes have been shown to have few, if any, cristae. Stage III-IV-V macrogametocyte exhibit an increase in cristate mitochondria, whereas microgametes have very few mitochondria [17, 36]. Furthermore, susceptibility to drugs and metabolic inhibitors is also reduced during the end of gametocytogenesis, except for the drugs derived from artemisinin and the 8-aminoquinoline primaquine. Although the mode of action of these drugs is still unclear, it is probably related to mitochondria and oxygen consumption in the case of primaquine [37, 38]. Artemisinin is thought to inhibit a sarco/endoplasmic reticulum Ca2+-ATPase (SERCA)-type P. falciparum protein, called PfATP6 [39]. It has also been shown that the P. falciparum mitochondrial gene encoding cytochrome b had an expression several folds higher in sexual stages than in asexual stages [40, 41]. These differences in structure and chemical susceptibility have been hypothesized to be due to the development of an active tricarboxylic acid (TCA) cycle and respiration during gametocytogenesis. Although a differential oxygen consumption has not been clearly established, the possibility of a metabolically active, yet under-developed, TCA cycle has been suggested but remains to be proved [36, 42].
Another function of the electron transport system (ETS) is the pyrimidine biosynthesis. The latter evidence of mitochondrial activity could be accounted for by this mechanism. The ETS has been shown to be active in both the asexual and sexual parasite by the identification of the enzyme and by drug sensitivity assays [43]. Functional mitochondria of both stages contribute to the de novo synthesis of pyrimidine [36]; the enzyme by which this process is achieved is dihydroorotate dehydrogenase (DHODase). Drugs that inhibit specifically DHODase were found to affect P. falciparum : atovaquone and salicylhydrox-amic acid have been reported to inhibit the survival of asexual stages and early-stage gametocytes but not that of late stages. This tends to indicate that de novo pyrimidine synthesis in early stages of gametocytes is sufficient for the whole maturation process [43, 44].
Phosphoenolpyruvate carboxykinase (PfPEPCK) is another differentially-expressed enzyme known to catalyse CO2 fixation. The transcription and activity of this enzyme was found to be up-regulated in gametocytes and zygotes as compared to asexual stages. It was hypothesised that this enzyme was abundantly transcribed in gametocytes in order for the post-fertilization stages to adapt to the scarcer access to glucose in the mosquito haemolymph by means of an alternative ATP source through a gluconeogenesis pathway [45, 46].
Asexual blood parasites and young gametocytes rely extensively on haemoglobin digestion as a source of amino acid and iron [47]. As one might predict, this mechanism is abandoned during maturation of gametocytes, possibly between stage III and IV [46].
Whereas asexual metabolism has been quite well characterized, there is a general lack of research on the metabolism of gametocytes. Gametocytocidal drug development deserves more attention in view of their potential role in reducing transmission.
Sequestration of gametocytes
The mature asexual stages of P. falciparum are absent from the peripheral circulation, due to the adherence of infected erythrocytes to microvascular endothelia of many organs and tissues such as heart, lung, liver, skin and brain [48]. Sequestration allows avoidance of phagocytic clearance in the spleen during maturation [49]. A similar phenomenon is also observed in maturing gametocytes: stages I to IV are sequestered preferentially in the bone marrow and spleen [50, 51], whilst stage V are released in the peripheral circulation and only become infectious to mosquitoes after a further two or three days of circulation [10, 52].
The mechanism of sequestration of asexual stages is that of cytoadherence of infected red blood cells (IRBCs), mediated by a series of host receptors: intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1), P-selectin, thrombospondin (TSP) and most strongly with the glycoprotein CD36 [[53–57], reviewed in [58]]. The parasite ligand for CD36, ICAM-1 and TSP has been identified as the P. falciparum erythrocyte membrane protein 1 (PfEMP-1) [59]. This protein is encoded by the highly variant var gene family [60, 61].
Day and colleagues [62] have shown that the cytoadherence of stage I and IIA to C32 melanoma cells is indistinguishable from that of the asexual stages. CD36 binding was observed, as well as knobs on the surface of erythrocyte and HRP-1 expression. Furthermore, the PfEMP-1 pattern of expression is similar in these very young gametocytes compared to the one observed in asexual stages [63]. This is further corroborated by the work of Smith and colleagues [64] who observed CD-36 PfEMP-1 mediated non-opsonic phagocytosis of stage I and IIA infected erythrocytes, confirming the importance of the CD36-PfEMP-1 interaction in the sequestration of these stages. It is interesting to note that a deletion on chromosome 9 results in loss of both adhesion of asexual stages and gametocytogenesis [65].
Stage III and IV are also sequestered in the bone marrow, yet it has been found that the mechanism mediating this process is different from that of other stages: PfEMP-1 is not expressed on the surface after stage IIB [66], sequestration in bone marrow does not involve a PfEMP-1-CD36 interaction [67] and CD36 mediated phagocytosis is greatly reduced in late stages [64]. Candidate low avidity receptors for sequestration of stage IIB to IV have been identified, they include ICAM-1, CD49c, CD166 and CD164 [67]. The gametocyte ligand for this interaction remains unidentified.
Nacher [68] suggested that the specific banana shape of P. falciparum gametocytes could have been selected naturally to facilitate the detachment of mature transmission stages from the bone marrow, the sausage shape parasite being more susceptible to rheological forces.
On the available evidence to-date, one can hypothesize that the asexual blood stages and stage I and IIA gametocytes use the same mechanisms for sequestration. One might ask how do the asexual and early sexual stages sequester in different locations, with the same mechanisms, and what mechanism mediates this differential sequestration? And what is the advantage of changing of sequestration mechanism in the course of gametocytogenesis?
Commitment to gametocytogenesis
Genetic variation in commitment to sexual development
Differences in the rate of sexual conversion have been reported between isolates of P. falciparum from different patients [69, 70], and between cloned lines derived from the same isolate [71]. Individual clones in vitro also seem to lose spontaneously the ability to produce gametocytes after in vitro culture [72]; the period necessary to lose this ability is variable amongst clones, from a few weeks to more than a year [65, 70, 71, 73]. This suggests that a genetic factor influences the rate of conversion to sexual development in P. falciparum. There is a selection shift when the parasite is grown in vitro. Transmission stages are no longer selected for, which could explain the variation in isolates but not for individual clones. The probable mechanism accounting for this variation is chromosome deletion, especially chromosome 9 subtelomeric deletion, which is commonly observed in cultured parasites [65, 74]. One could also imagine single gene defects, for instance, the knock-out of the Pfg27 leads to loss of sexual phenotype [75]. The observation that in vitro lines that had not produced gametocytes for many years could still be environmentally stimulated [76, 77] could indicate that a mechanism for loss of sexual phenotype is the loss of capacity to respond to the environment, which would be bypassed by artificial induction.
Environmental factors influencing commitment to sexual development
The first demonstration that the rate of conversion was influenced by environmental factors was that of Carter and Miller [78]. The study demonstrated in three different strains that the addition of fresh blood cells to the culture and a lower parasitaemia significantly reduced commitment to gametocytogenesis. Since then many factors have been found to influence gametocytogenesis, even if there have been problems in reproducing some of these results [79].
Host factors
The rate of commitment to gametocytes production was shown to be influenced by host factors, of which the most important is host immunity. An induction of gametocytogenesis due to increased immune pressure was demonstrated in P. yoelii using immunization of a mouse by a protein expressed throughout the life cycle [80] and for Plasmodium chabaudi in partially immunized mouse [81]. This stimulation was also found in vitro for P. falciparum using lymphocytes and serum from P. falciparum- infected children [82] and the culture supernatant of hybridoma cells producing anti-P. falciparum antibody [83]. Although these results must be taken with caution as for most of them a decrease in gametocytaemia was associated with the increased proportion of gametocyte or rate of commitment to gametocytogenesis. However, as the immune system acts upon both the supply of gametocytes (asexual parasites), it is plausible that a higher commitment to gametocytogenesis is stimulated by immune stress. Further investigations are needed.
Host steroids and corticosteroids have also been associated with a higher commitment to sexual development [84]. Furthermore, there are reports that, in vitro, a high proportion of reticulocytes in the blood stimulates gametocytogenesis [85, 86]. Anaemia of the host is also a risk factor associated with gametocyte carriage; as anaemia is usually due to long term poorly treated or untreated infections, increased gametocyte density might be the result of the length of the infection more than that of an adaptive mechanism for optimization of sexual stage development [87, 88].
Drug-treament
The induction of gametocytogenesis by treatment has been shown for both chloroquine [89–91] and for sulfadoxine+pyrimethamine [92, 93]. This effect has also been shown to be enhanced in resistant strains [92, 94, 95]. These effects are to be taken with caution as data for untreated patients are impossible to collect, for obvious ethical reasons. The mechanisms by which this commitment would be enhanced are quite obscure; however, the "stress" of treatment on the asexual stages may act as a positive feedback on gametocytogenesis (see below). It is worth noting that artemisinin derivatives combination therapy has been shown to be gametocytocidal [96] and to reduce but not suppress transmission [95, 97].
Signal transduction pathway
The fact that a population of parasites is capable of having a marked response to the environment indicates that there are means of receiving and responding to environmental signals. There is consistent evidence of the involvement of cAMP-dependent and Protein Kinase C-dependent pathways involved in the induction of gametocytogenesis [70, 98–104]. Yet the exact pathway for induction of gametocytogenesis is still to be determined.
Parasite factors
Mixed-genotype infections have been associated with a higher gametocytaemia in P. chabaudi, indicating that the parasite responds in terms of transmission to the presence of a competitor [105]. This has, however, not been investigated in P. falciparum. There also is a widely recognized correlation between asexual parasitaemia and commitment to sexual development. It is commonly accepted that, when the levels of asexual parasitaemia decrease, the rate of conversion to gametocytogenesis will increase [79, 106]. An autocrine factor may be responsible for the link between asexual parasitaemia and stimulation of sexual conversion. This was first demonstrated by Williams [107], who observed higher conversion rates in cultures supplemented with conditioned media and grown in co-culture. This was also observed by Dyer & Day [108], who hypothesized that the sexual pathway is the default pathway and that its inhibition by an autocrine factor would favour asexual growth in the first instance, the sexual route being then favoured in a density-dependent manner. The fact that lysed parasitized erythrocytes stimulate gametocytogenesis [109] could indicate that this factor is liberated at schizogony with the rupture of the erythrocyte. However, the actual factors responsible and the way in which they can modify the level of sexual conversion are still to be determined.
The parasite appears to have the ability to modulate the proportion of the parasite population within an infection that undergoes sexual development. In 1908, Stephens and Christophers concluded that gametocytogenesis increases when conditions are unfavourable to the parasite [110]. Indeed "stress" seems to be the general enhancer of sexual conversion whether from the host immunity, drug-pressure and competing parasites in the host. This fast adaptive system seems to be regulated by a signal transduction pathway and allows P. falciparum to maximise its transmission success.
The sex of gametocytes
Plasmodium species are hermaphrodite. An asexual parasite can develop after several rounds of multiplication into either a male or a female gametocyte [111]. In P. falciparum, gametocytes are produced from asexual stages. All the gametocytes produced from one sexually committed schizont are of the same sex, suggesting that sex is determined at the very beginning of sexual development [14]. However, gametocyte sex can only be microscopically differentiated from stage III onwards.
Sex-ratio in malaria parasite
The sex ratio has been shown to be generally female-biased in P. falciparum and related protozoan species [94, 112–117]. In P. falciparum, ordinarily one male gametocyte is observed for three or four females. This is mainly explained by the fact that one male gametocyte can give rise to up to eight male gametes, thus establishing an approximate 1:1 ratio in the mosquito midgut. However, large fluctuations in gametocyte sex ratio are observed [94, 113, 118].
The work of Burkot and colleagues [111] suggests that the sex ratio is clone specific and would, therefore, have a genetic component. This is widely accepted in the literature given the role of natural selection in determining sex ratios [112, 114].
Sex allocation theory
The way by which Plasmodium modulates its gametocyte production and gametocyte sex ratio has recently been reappraised within an evolutionary framework wherein gametocyte allocation is considered as an adaptive phenotype. The framework proposes two complementary cues to explain the observed gametocyte sex ratio variation: (a) the average complexity of infection and thus the level of in-breeding and (b) that the sex ratio is optimized to ensure successful fertilization in the short term.
Complexity of infection
In-breeding is thought to play a role in the observed variation in sex ratio [112]. If having an equal number of male and female (in the mosquito) is the optimal sex allocation strategy in a random mating population [119], a bias in the sex ratio is expected where this condition is not fulfilled. In a monoclonal population (100% inbred), the sex ratio that maximizes the success of transmission, i.e. the one producing the greatest number of zygotes, will be favoured, hence creating a very female-biased sex ratio. Whereas in the case of outcrossing, natural selection will favour a less female biased sex ratio, because a clone producing more males will have a higher genetic representation in the progeny than one producing a more female biased sex ratio. This corresponds in fact to a shift in the level of selection from the clone level to the individual parasite level.
Malaria parasites do not mate randomly and can exhibit elevated levels of in-breeding [120], which can contribute to the observation of a female biased sex ratio. A few studies bring support to these predictions in P. falciparum [112, 113, 121]. But this alone is insufficient to explain all the fluctuations observed [122].
Fertility insurance
The parasites also have fast adaptive mechanisms to respond to the environment in order to maximize fertility. It has been shown that they respond to at least two factors. Anaemia appears to favour a less female-biased sex ratio [118, 123]. An increase in the human hormone erythropoietin, induced by anaemia, directly or indirectly triggers this higher commitment to the production of male parasites [123]. As infection progresses, malaria-induced anaemia coincides with an increase in immune pressure against gametocytes; such a pressure would disadvantage the male gamete in the mosquito gut, given their motile status and the urgency to find a female to mate with (within 30 minutes for a successful fertilization). A more male-biased sex ratio would, therefore, be more favourable for a successful transmission [122]. Another possible explanation is that anaemia reduces the chance of encounter between male and female gametes, because of the lower gametocyte content in the bloodmeal [124]: a higher proportion of male would, thus, maximize the chances of fertilization [5]. A recent study by Reece et al. [125] demonstrates a higher longevity of male gametocytes in P. chabaudi. This could account for some of the variation in sex ratio observed in the course of an infection. However, this question needs to be further investigated in P. falciparum.
The second related factor is the temporal fluctuations of gametocytaemia. A gametocytaemia peak was associated with a higher proportion of females from this peak during the two following weeks. This would be favourable in that the encounter of a male and female is guaranteed by the large number of gametocytes, a balanced gamete sex ratio, therefore, giving the greatest reproductive success [118]. Other factors which have been shown to induce gametocytogenesis do not appear to influence sex ratios, such as the vertebrate host testosterone [126] and drug-treatment [91]. This also suggests that sexual development and sexual differentiation operate through two distinct induction pathways. It is vital to understand the mechanism of sex determination and of sex ratio control given the importance that sex ratios have on the infectivity to mosquitoes [94, 122].
Evolutionary considerations
Sex in the Apicomplexa
Sexual reproduction is a common phenomenon in natural populations; all apicomplexan have a sexual stage in the course of their life cycle [127]. The advantages conferred by sex have intrigued evolutionary biologists for many years. The various theories [128–132] fall into one of two types of models: the environmental model, which suggests that sex has evolved to allow adaptation to a changing environment by favouring recombination, and the mutation-based models, which assume that sex is advantageous to suppress deleterious mutations more rapidly. However, the explanation might be in a pluralistic approach [133].
Aside from the nucleus, P. falciparum has two organelles containing DNA: the mitochondria (linear, 8 kb) and the apicoplast (circular, 35 kb). The latter seems to be evolutionarily related to a structure present in red algae and chloroplast of plants. This plastid is essential to the parasite, but its comprehensive role is still uncertain [134]. The exclusive maternal inheritance of this structure suggests the existence of a cytoplasmic incompatibility between male and female gametes, as previously demonstrated in plants [135]. Such incompatibility had been strongly suspected in outcrossing experiments and could be linked to reproductive isolation of the parasite from different geographical areas [136]. In the same way, research into evolutionary maintenance of sex can benefit from the large amount of work already done in plants and algae, of which the sex life seems quite related or may even be homologous to that of P. falciparum.
Apicomplexan parasites supposedly benefit from genetic recombination following sexual achievement during meiosis. Recombination has been shown to occur in P. falciparum [137–140], Plasmodium vivax [141], P. chabaudi [105], and Toxoplasma gondii [142].
Mixed infections
Where there is recombination, there has to be at least two genetically distinct parasites; the interaction of several populations of different genotypic types remains elusive. The adaptive sex ratio function of complexity of the parasite population in the long term has already been discussed (see Sex allocation theory). It was postulated that the parasite could also have a fast adaptive system responding to the complexity of the infection [143]. This was borne out by the fact that multiple-genotype infections are often more successful in terms of transmission than single-genotype infections [105]. This might not reflect co-operation between conspecific clones, but it could be that competition between parasites increases the chances of transmission by creating an advantageous effect on the competiting clones. This may be confirmed by the work of Arez and colleagues [144], who observed a lower proportion of mixed-genotype infections in the mosquito than in the human host. This may, however, only be due to the fact that mixed-infections are not necessarily synchronous, which lowers the chances of recombination. The dynamics of genotype interaction needs to be further characterized, to reveal how it might affect both the disease and the transmission.
Commitment to gametocytogenesis has also been shown to be altered when another species of human malaria parasite is present. For instance, the presence of Plasmodium malariae may boost the gametocyte production of P. falciparum [145].
Why so few gametocytes?
Not only are gametocytes the first step to sexual genetic recombination, but they also are the transmission stage. A great selection pressure is, therefore, upon them. The transmission phenotypes observed in natural infections must be ones that optimize the greater chances of a successful shift of host. A positive correlation has been found between gametocyte density in the blood and infectiousness to mosquitoes [94, 146–148]. However, this correlation must be qualified as loose and may be hampered by the low sensitivity of microscopy [149] and by transmission-blocking immunity [19].
Plasmodium and closely related apicomplexan have evolved an asexual erythrocytic cycle. This character is thought to have evolved several times in the Apicomplexa [127]; an acceptable explanation is that erythrocytic proliferation allows them to generate more transmission stages than with tissue merogony alone. Furthermore, Dyer and Day [79] proposed that indefinitive rounds of asexual proliferation would augment the length of time of circulation of gametocytes and therefore increase the chances for successful transfer to the mosquito. One cannot forget, however, that various related parasites, such as Leucocytozoa and Haemoproteus, release only sexual stages in the blood and are very successfully transmitted [150].
More intriguing is the fact that very few of the erythrocytic stages actually commit to a differentiation to transmission stages in the blood [reviewed in [151]]. It is paradoxical that, having the possibility to generate more gametocytes through asexual proliferation, the parasite actually makes less, especially since a higher density would increase transmission [152]. Several hypotheses have been proposed to explain this restraint.
Taylor and Read [151] have postulated that immune pressure through transmission-blocking activity could account for the low density of gametocytes. Transmission-blocking immunity has been demonstrated for several surface antigens of P. falciparum. If this anti-gamete immunity is dependent on the density of gametocytes, it is obviously advantageous to reduce the number of gametocytes. Alternatively, it could be that the level of this immunity was dependent on the ratio of asexual: sexual parasites, the asexual parasites acting as a decoy for the immune system. Piper and colleagues [63] proposed a role for a cross-stage immunity against PfEMP-1. An increase of anti-PfEMP-1 immunity with age was observed, which was accompanied by a decline in prevalence and density of asexual and sexual blood stages of P. falciparum. PfEMP-1 being expressed on the surface of both asexual stages and young gametocytes (see Sequestration of gametocytes), regulation of the level of gametocytes could have evolved either by controlling asexual proliferation (the source of gametocytes) or directly by affecting maturing gametocytes.
It has also been proposed that frequencies of super-infection, modulated by cross-immunity between the phenotypes involved, may serve to constrain gametocyte production to levels less than would be optimal for transmission in a single-phenotype context [153].
Another possible explanation is that the gametocyte number is kept low in order to reduce the damage mosquito stages are likely to inflict on their host [151]. Evidence that natural selection drives the parasite towards lessening the number of mosquito stages in a mosquito to favour survival of the host and, therefore, successful completion of the life cycle, has only recently come to light. The ookinete, which is responsible for the penetration of the stomach wall, is the most harmful stage to the mosquito; this stage has been shown to be capable of apoptosis (programmed cell death) [154], which results in a reduction in the number of ookinetes and, therefore, lessens the damage due to penetration and eventual opportunistic bacterial infection.
It has been demonstrated that the parasite has evolved several strategies to maximize transmission in spite of the low number of gametocytes: (a) aggregation of gametocytes to favour encounter of males and females in the bloodmeal [155] (b) preferential localization of infectious rodent-malaria gametocytes in sub-dermal capillaries [156] and possible sequestration of mature gametocytes in the derma [157] and (c) active suppression of insect melanization by ookinetes [158].
Malaria parasites may have evolved restraint in the production of transmission stages in order to avoid harmful consequences either directly for the parasite through the immune system of the definitive host or to spare the vector in order to maximize its success in transmitting the parasite. This restraint provides an evolutionary justification for the development of autocrine/exocrine mechanisms for means of communication. One could imagine that asexual parasites express a highly labile diffusible molecule [108] that prevents other asexual parasites from undergoing gametocytogenesis or encourages them to pursue asexual proliferation. When a high level of "stress" (immunity) arises, the parasite population would respond by restricting the expression of the diffusible molecule and, therefore, would stimulate a higher level of asexual parasites to develop sexually.
Conclusions
P. falciparum gametocytes exhibit adaptability and biological achievement. The gametocytogenesis of this parasite, comparable to a puberty process, obviously contributes to its present tropical-wide success. The present review addressed several aspects of gametocytes, focusing on sex differentiation and determination, transmission strategies and evolution; such a review endeavours to highlight the areas where the gametocyte might be targetted for control interventions.
To date, reported attempts to fight the parasite burden by targetting the gametocyte are non-existent. However, when one considers malaria control strategies as a whole, gametocytes are often secondary targets. Any anti-anopheline intervention, as it reduces vector density, also decreases the passage of gametocytes from man to mosquito. Newly developed drugs, such as combination therapy with artemisinin derivatives, have as their main purpose the efficient removal of asexual stages, but the gametocytocidal activity of these drugs is considered an important issue. There also is a recent "courant de pensée"among the scientific community that the management of anti-malarial resistance will necessarily involve gametocyte control.
Author contributions
AMT scanned the literature and wrote the draft. All authors contributed to the writing of the manuscript and approved the submitted version.
References
Bell G: The masterpiece of nature: The evolution and genetics of sexuality. 1982, Berkeley: University of California Press
Otto SP, Lenormand T: Resolving the paradox of sex and recombination. Nat Rev Genet. 2002, 3: 252-261. 10.1038/nrg761.
Eichner M, Diebner HH, Molineaux L, Collins WE, Jeffery GM, Dietz K: Genesis, sequestration and survival of Plasmodium falciparum gametocytes: parameter estimates from fitting a model to malariatherapy data. Trans R Soc Trop Med Hyg. 2001, 95: 497-501. 10.1016/S0035-9203(01)90016-1.
Day KP, Hayward RE, Dyer M: The biology of Plasmodium falciparum transmission stages. Parasitology. 1998, 116: S95-S109. 10.1017/S0031182097002370.
Robert V, Boudin C: Biologie de la transmission homme-moustique du Plasmodium. Bull Soc Pathol Exot. 2003, 96: 6-20.
Kitchen SF, Putnam P: Observations on the mechanism of the parasite cycle in falciparum malaria. Am J Trop Med. 1942, 22: 381-386.
Kitchen SF: Falciparum malaria. In Malariology. Edited by: Boyd MF. 1949, Philadelphia: WB Saunders, 995-1016.
Sinden RE: Sexual development of malaria parasite. Adv Parasitol. 1983, 22: 153-216.
Carter R, Graves PM: Gametocytes. In Malaria. Principles and practise of malariology. Edited by: Wernsdorfer WH & Mc Gregor I. 1988, Edinburgh, Scotland, Churchill Livingstone, 1-59.
Smalley ME, Sinden RE: Plasmodium falciparum gametocytes their longevity and infectivity. Parasitology. 1977, 74: 1-8.
Garnham PCC: Malaria parasites and other haemosporisia. 1966, Oxford (UK): Blackwell Scientific
Bruce MC, Alano P, Duthie S, Carter R: Commitment of the malaria parasite Plasmodium falciparum to sexual and asexual development. Parasitology. 1990, 100: 191-200.
Smith TG, Lourenço P, Carter R, Walliker D, Ranford-Cartwright LC: Commitment to sexual differentiation in the human malaria parasite, Plasmodium falciparum. Parasitology. 2000, 121: 127-133. 10.1017/S0031182099006265.
Silvestrini F, Alano P, Williams JL: Commitment to the production of male and female gametocytes in the human malaria parasite Plasmodium falciparum. Parasitology. 2000, 121: 465-471. 10.1017/S0031182099006691.
Field JW, Shute PG: The microscopic diagnostic of human malaria. In A morphological study of the erythrocytic parasites. 1956, Kuala Lumpur: Government Press, 142-
Hawking F, Wilson ME, Gammage K: Evidence for cyclic short-lived maturity in the gametocytes of Plasmodium falciparum. Trans R Soc Trop Med Hyg. 1971, 65: 549-559. 10.1016/0035-9203(71)90036-8.
Langreth SG, Jensen JB, Reese RT, Trager W: Fine structure of human malaria in vitro. J Protozool. 1978, 25: 443-452.
Sinden RE: Gametocytogenesis of Plasmodium falciparum in vitro: an electron microscopic study. Parasitology. 1982, 84: 1-11.
Sauerwein RW, Eling WMC: Sexual and sporogenic stage antigens. Chem Immunol. 2002, 80: 188-203.
Graves PM, Wirtz RA, Carter R, Burkot TR, Looker M, Targett GA: Naturally occurring antibodies to an epitope on Plasmodium falciparum gametes detected by monoclonal antibody-based competitive enzyme-linked immunosorbent assay. Infect Immun. 1988, 56: 2818-2821.
Waters AP, Syin C, McCutchan TF: Developmental regulation of stage-specific ribosome population in Plasmodium. Nature. 1989, 342: 238-250. 10.1038/342438a0.
McCutchan TF, Li J, McConkey GA, Rogers MJ, Waters AP: The cytoplasmic ribosomal RNAs of Plasmodium spp. Parasitol Today. 1995, 11: 134-138. 10.1016/0169-4758(95)80132-4.
Lasonder E, Ishiham Y, Andersen JS, Vermunt AMW, Pain A, Sauerwein RW, Eling WMC, Hall N, Waters AP, Stunnenberg HG, Mann M: Analysis of the Plasmodium falciparum proteome by high-accuracy mass spectrometry. Nature. 2002, 419: 537-542. 10.1038/nature01111.
Florens L, Washburn MP, Raine JD, Anthony RM, Grainger M, Haynes JD, Moch JK, Muster N, Sacci JB, Tabb DL, Witney AA, Wolters D, Wu Y, Gardner MJ, Holder AA, Sinden RE, Yates JR, Carucci DJ: A proteomic view of the Plasmodium falciparum life cycle. Nature. 2002, 419: 520-526. 10.1038/nature01107.
Alano P, Carter R: Sexual differentiation in malaria parasites. Ann Rev Microbiol. 1990, 44: 429-449. 10.1146/annurev.mi.44.100190.002241.
Gardner MJ, Shallom SJ, Carlton JM, Salzberg SL, Nene V, Shoaibi A, Ciecko A, Lynn J, Rizzo M, Weaver B, Jarrahi B, Brenner M, Parvizi B, Tallon L, Moazzez A, Granger D, Fujii C, Hansen C, Pederson J, Feldblyum T, Peterson J, Suh B, Angiuoli S, Pertea M, Allen J, Selengut J, White O, Cummings LM, Smith HO, Adams MD, Venter JC, Carucci DJ, Hoffman SL, Fraser CM: Sequence of Plasmodium falciparum chromosomes 2,10,11 and 14. Nature. 2002, 419: 531-534. 10.1038/nature01094.
Bozdech Z, Llinás M, Pulliam BL, Wong ED, Zhu J, DeRisi JL: The transcriptome of the intraerythrocytic developmental cycle of Plasmodium falciparum. PLOS Biology. 2003, 1: 85-100. 10.1371/journal.pbio.0000005.
Janse CJ, Ponnudurai T, Lensen AHW, Meuwissen JHE, Ramesar J, Van Der Ploeg M, Overdulve JP: DNA synthesis in gametocyte of Plasmodium falciparum. Parasitology. 1988, 96: 1-7.
Janse CJ, Waters AP: Sexual development of malaria parasites. In Malaria parasites: genomes and molecular biology. Edited by: Waters AP, Janse CJ. 2004, Leiden: Caister Academic Press, chap 14-
Janse CJ, Van Der Klooster PFJ, Van Der Kaay HJ, Van Der Ploeg M, Overdulve JP: DNA synthesis in Plasmodiun berghei during asexual and sexual development. Mol Biochem Parasitol. 1986, 20: 173-183. 10.1016/0166-6851(86)90029-0.
Sinden RE: Mitosis and Meiosis in malarial parasites. Acta Leidensa. 1991, 60: 19-27.
Shaw MK, Thompson JT, Sinden RE: Localization of ribosomal RNA and Pbs21-mRNA in the sexual stages of Plasmodium berghei using electron microscope in situ hybridization. EurJ Cell Biol. 1996, 71: 270-276.
Vizioli J, Catteruccia F, della Torre A, Reckmann I, Mailer HM: Blood digestion in the malaria mosquito Anopheles gambiae. Eur J Biochem. 2001, 268: 4027-4035. 10.1046/j.1432-1327.2001.02315.x.
Geary TG, Divo AA, Bonanni LC, Jensen JB: Nutritional requirements of Plasmodium falciparum in culture. III. Further observations on essential nutrients and antimetabolites. J Protozool. 1985, 32: 608-613.
Riandey MF, Sannier C, Peltre G, Monteny N, Cavaleyra M: Lactate dehydrogenase as a marker of Plasmodium infection in malaria vector Anopheles. J Am Mosq Con Assoc. 1996, 12: 194-198.
Krungkrai J, Burat D, Kudan S, Krungkrai SR, Prapunwattana P: Mitochondrial oxygen consumption in asexual and sexual blood stages of the human malaria parasite Plasmodium falciparum. S Asian J Trop Med Pub Health. 1999, 30: 636-642.
Lanners HN: Effect of the 8-aminoquinoline primaquine on culture-derived gametocytes of the malaria parasite Plasmodium falciparum. Parasitol Res. 1991, 77: 478-481.
Kawai S, Kano S, Suxuki M: Morphologic effects of arthemeter on Plasmodium falciparum in Aotus trivirgatus. Am J Trop Med Hyg. 1993, 49: 812-818.
Eckstein-Ludwig U, Webb RJ, van Goethem IDA, East JM, Lee AG, Kimura M, O'Neil PM, Bray PG, Ward SA, Krishna S: Artemisinins target the SERCA of Plasmodium falciparum. Nature. 2003, 424: 957-961. 10.1038/nature01813.
Petmir S, Krungkrai J: Mitochondrial cytochrome b gene in two developmental stages of human malaria parasite Plasmodium falciparum. S Asian J Trop Med Pub Health. 1995, 26: 600-605.
Learngaramkul P, Petmir S, Krungkrai SR, Prapunwattana P, Krungkrai J: Molecular characterisation of mitochondria in asexual and sexual blood stages of Plasmodium falciparum. Mol Cell Biol Res Commun. 1999, 2: 15-20. 10.1006/mcbr.1999.0145.
Krungkrai J, Prapunwattana P, Krungkrai SR: Ultrastructure and function of mitochondria in gametocytic stage of Plasmodium falciparum. Parasite. 2000, 7: 19-26.
Unnasch NL, Murphy AD: Metabolic changes of the malaria parasite during the transition from the human to the mosquito host. Ann Rev Microbiol. 1998, 52: 561-590. 10.1146/annurev.micro.52.1.561.
Fleck SL, Pudney M, Sinden RE: The effect of atovaquone (566C80) on the maturation and viability of Plasmodium falciparum gametocytes in vitro. Trans R Soc Trop Med Hyg. 1996, 90: 309-312. 10.1016/S0035-9203(96)90266-7.
Clements AN: The circulatory system. In The biology of mosquitoes. Edited by: Clements AN. 1984, London: Chapman and Hall, 1: 195-205.
Hayward RE: Plasmodium falciparum phosphoenolpyruvate carboxykinase is developmentally regulated in gametocytes. Mol Biochem Parasitol. 2000, 107: 227-240. 10.1016/S0166-6851(00)00191-2.
Rosenthal PJ, Meshnick SR: Hemoglobin processing and the metabolism of amino acids, heme, and iron. In Malaria: parasite biology, pathogenesis, and protection. Edited by: Sherman IW. 1998, Washington DC: American Society for Microbiology Press, 145-158.
MacPherson GG, Warrell MJ, White NJ, Looareesuwan S, Warrell DA: Human cerebral malaria. A quantitative ultrastructural analysis of parasitised erythrocyte sequestration. Am J Pathol. 1985, 119: 385-401.
Howard RJ: Malarial proteins at the membrane of Plasmodium falciparum infected erythrocytes and their involvement in adhesion to endothelial cells. Prog Allergy. 1988, 41: 98-147.
Thomson JG, Robertson A: The structure and development of Plasmodium falciparum gametocytes in the internal organs and peripheral circulation. Trans R Soc Trop Med Hyg. 1935, 29: 31-40. 10.1016/S0035-9203(35)90015-3.
Smalley ME, Abdalla S, Brown J: The distribution of Plasmodium falciparum in the peripheral blood and bone marrow of Gambian children. Trans R Soc Trop Med Hyg. 1980, 75: 103-105. 10.1016/0035-9203(81)90019-5.
Lensen A, Bril A, van de Vegte M, van Gemert GJ, Eling W, Sauerwein R: Plasmodium falciparum: infectivity of cultured, synchronized gametocytes to mosquitoes. Exp Parasitol. 1999, 91: 101-103. 10.1006/expr.1998.4354.
Barnwell JW, Asch AS, Nachman RL, Yamaya M, Aikawa M, Ingravallo P: A human 88-kD membrane glycoprotein (CD36) functions in vitro as a receptor for a cytoadherence ligand on Plasmodium falciparum-infected erythrocytes. J Clin Invest. 1989, 84: 765-772.
Ockenhouse CF, Tandon NN, Magowan C, Jamieson GA, Chulay JD: Identification of a platelet membrane glycoprotein as a falciparum malaria sequestration receptor. Science. 1989, 243: 1469-1471.
Roberts DD, Sherwood JA, Spitalnik SL, Panton LJ, Howard RJ, Dixit VM, Frazier WA, Miller LH, Ginsburg V: Thrombospondin binds falciparum malaria parasitised erythrocytes and may mediate adhesion. Nature. 1985, 318: 64-66.
Berendt AR, Simmons DL, Tansey J, Newbold Cl, Marsh K: Intercellular adhesion molecule-1 is an endothelial cell adhesion receptor for Plasmodium falciparum. Nature. 1989, 341: 57-59. 10.1038/341057a0.
Ockenhouse CF, Tegoshi T, Maeno Y, Benjamin C, Ho M, Kan KE, Thway Y, Win K, Aikawa M, Lobb RR: Human vascular endothelial cell adhesion receptors for Plasmodium falciparum – infected erythrocytes: roles for ELAM-1 and VCAM-1. J Exp Med. 1992, 176: 1183-1189. 10.1084/jem.176.4.1183.
Sherman IW, Eda S, Winograd E: Cytoadherence and sequestration in Plasmodium falciparum : defining the ties that bind. Microb Infect. 2003, 5: 897-909. 10.1016/S1286-4579(03)00162-X.
Baruch Dl, Gormely JA, Ma C, Howard RJ, Pasloske BL: Plasmodium falciparum erythrocyte membrane protein 1 is a parasitized erythrocyte receptor for adherence to CD36, thrombospondin, and intercellular adhesion molecule 1. Proc Natl Acad Sci U S A. 1996, 93: 3497-3502. 10.1073/pnas.93.8.3497.
Baruch Dl, Pasloke BL, Singh HB, Bi X, Ma XC, Feld-man M, Taraschi TF, Howard RJ: Cloning the Plasmodium falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995, 82: 77-87. 10.1016/0092-8674(95)90054-3.
Su XZ, Heatwole VM, Wertheimer SP, Guinet F, Herrfeldt JA, Peterson DS, Ravetch JA, Wellems TE: The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum infected erythrocytes. Cell. 1995, 82: 89-100. 10.1016/0092-8674(95)90055-1.
Day KP, Hayward RE, Smith D, Culvenor JG: CD36-dependent adhesion and knob expression of the transmission stages of Plasmodium falciparum is stage-specific. Mol Biochem Parasitol. 1998, 93: 167-177. 10.1016/S0166-6851(98)00040-1.
Piper KP, Hayward RE, Cox MJ, Day KP: Malaria transmission and naturally acquired immunity to PfEMP-1. Infect Immun. 1999, 67: 6369-6374.
Smith TG, Serghides L, Patel SN, Febbraio M, Silverstein RL, Kain KC: CD36-mediated nonopsonic phagocytosis of erythrocytes infected with stage I and IIA gametocytes of Plasmodium falciparum. Infect Immun. 2003, 71: 393-400. 10.1128/IAI.71.1.393-400.2003.
Day KP, Karamalis F, Thompson J, Barnes DA, Peterson C, Brown H, Brown GV, Kemp DJ: Genes necessary for expression of a virulence determinant and for transmission of Plasmodium falciparum are located on a 0.3 megabase region of chromosome 9. Proc Natl Acad Sci USA. 1993, 90: 8292-8296.
Hayward RE, Tiwari B, Piper KP, Baruch Dl, Day KP: Virulence and transmission success of the malarial parasite Plasmodium falciparum. Proc Natl Acad Sci USA. 1999, 96: 4563-4568. 10.1073/pnas.96.8.4563.
Rogers NJ, Hall BS, Obiero J, Targett GAT, Sutherland CJ: A Model for sequestration of the transmission stages of Plasmodium falciparum : Adhesion of gametocytes-infected erythrocytes to human bone marrow cells. Infect Immun. 2000, 68: 3455-3462. 10.1128/IAI.68.6.3455-3462.2000.
Nacher M: Does the shape of Plasmodium falciparum gametocytes have a function?. Med Hypotheses. 2004, 62: 618-619. 10.1016/j.mehy.2003.11.011.
Chin W, Collins WE: Comparative studies of three strains of Plasmodium falciparum isolated by the culture method of Trager and Jensen. Am J Trop Med Hyg. 1980, 29: 1143-1146.
Brockelman CR: Conditions favouring gametocytogenesis in the continuous culture of Plasmodium falciparum. J Protozool. 1982, 29: 454-458.
Graves PM, Carter R, McNeil KM: Gametocyte production in cloned lines of Plasmodium falciparum. Am J Trop Med Hyg. 1984, 33: 1045-1050.
Bhasin VK, Trager W: Gametocyte-forming and non-gametocyte-forming clones of Plasmodium falciparum. Am J Trop Med Hyg. 1984, 33: 534-537.
Jeffery GM: Observations on a gametocyteless strain of Plasmodium falciparum. J Nat Malaria Soc. 1951, 10: 337-344.
Kemp DJ, Thompson J, Barnes DA, Triglia T, Karamalis F, Petersen C, Brown GV, Day KP: A chromosome 9 deletion in Plasmodium falciparum results in loss of cytoadherence. Mem Inst Oswaldo Cruz. 1992, 87 (Suppl 3): 85-89.
Lobo CA, Fujioka H, Aikawa M, Kumar N: Disruption of the Pfg27 locus by homologous recombination leads to loss of sexual of the sexual phenotype in Plasmodium falciparum. Mol Cell. 1999, 3: 793-798. 10.1016/S1097-2765(01)80011-3.
Ono T, Nakabayashi T: Gametocytogenesis induction by ammonium compounds in cultured Plasmodium falciparum. Int J Parasitol. 1990, 2: 615-618. 10.1016/0020-7519(90)90118-7.
Ono T, Ohnishi Y, Nagamune K, Kano M: Gametocytogenesis induction by Berenil in cultured Plasmodium falciparum. Exp Parasitol. 1993, 77: 74-78. 10.1006/expr.1993.1062.
Carter R, Miller LH: Evidence for environmental modulation of gametocytogenesis in Plasmodium falciparum in continuous culture. Bull World Health Organ. 1979, 57 (Suppl 1): 37-52.
Dyer M, Day KP: Commitment to gametocytogenesis in Plasmodium falciparum. Parasitol Today. 2000, 16: 102-107. 10.1016/S0169-4758(99)01608-7.
Motard A, Marussig M, Renia L, Baccam D, Landau I, Mattei D, Targett G, Mazier D: Immunization with the malaria heat shock like protein hsp70-1 enhances transmission to the mosquito. Int Immunol. 1995, 7: 147-150.
Buckling A, Read AF: The effect of partial host immunity on the transmission of malaria parasites. Proc R Soc Lond B Biol Sci. 2001, 68: 2325-2330. 10.1098/rspb.2001.1808.
Smalley ME, Brown J: Plasmodium falciparum gametocytogenesis stimulated by lymphocytes and serum from infected Gambian children. Trans R Soc Trop Med Hyg. 1981, 75: 316-317. 10.1016/0035-9203(81)90348-5.
Ono T, Nakai T, Nakabayashi T: Induction of gametocytogenesis in Plasmodium falciparum by the culture supernatant of hybridoma cells producing anti-P. falciparum antibody. Biken J. 1986, 29: 7781-7786.
Lingnau A, Margos G, Maier WA, Seitz HM: The effects of hormones on gametocytogenesis of Plasmodium falciparum in vitro. Appl Parasitol. 1993, 4: 153-160.
Trager W, Gill GS: Enhanced gametocyte formation in young erythrocytes by Plasmodium falciparum in vitro. J Protozool. 1992, 39: 429-432.
Trager W, Gill GS, Lawrence C, Nagel RL: Plasmodium falciparum: enhanced gametocyte formation in vitro in reticulocyte-rich blood. Exp Parasitol. 1999, 91: 115-118. 10.1006/expr.1998.4347.
Drakeley C, Secka I, Correa S, Greenwood BM, Targett GA: Host haematological factors influencing the transmission of Plasmodium falciparum gametocytes to Anopheles gambiae s.s. mosquitoes. Trop Med Int Health. 1999, 4: 131-138. 10.1046/j.1365-3156.1999.00361.x.
Price R, Nosten F, Simpson JA, Luxemburger C, Phaipun L, ter Kuile F, van Vugt M, Chongsuphajaisiddhi T, White NJ: Risk factors for gametocyte carriage in uncomplicated falciparum malaria. Am J Trop Med Hyg. 1999, 60: 1019-1023.
Hogh B, Gamage-Mendis A, Butcher GA, Thompson R, Begtrup K, Mendis C, Enosse SM, Dgedge M, Barreto J, Eling W, Sinden RE: The differing impact of chloroquine and pyrimethamine/ sulfadoxine upon the infectivity of malaria species to the mosquito vector. Am J Trop Med Hyg. 1998, 58: 176-82.
Buckling A, Ranford-Cartwright LC, Miles A, Read AF: Chloroquine increases Plasmodium falciparum gametocytogenesis in vitro. Parasitology. 1999, 118: 339-346. 10.1017/S0031182099003960.
Talman AM, Paul REL, Sokhna CS, Domarle O, Ariey F, Trape J-F, Robert V: Influence of chemotherapy on Plasmodium gametocytes sex ratio Of Mice and Men. Am J Trop Med Hyg.
Sokhna CS, Trape J-F, Robert V: Gametocytaemia in Senegalese children with uncomplicated falciparum malaria treated with chloroquine, amodiaquine or sulfadoxine + pyrimethamine. Parasite. 2003, 8: 243-250.
Puta C, Manyando C: Enhanced gametocyte production in Fansidar-treated Plasmodium falciparum malaria patients: implications for malaria transmission control programmes. Trop Med Int Health. 1997, 2: 227-229. 10.1046/j.1365-3156.1997.d01-267.x.
Robert V, Read AF, Essong J, Tchuinkam T, Mulder B, Verhave JP, Carnevale P: Effect of gametocyte sex ratio on infectivity of Plasmodium falciparum to Anopheles gambiae. Trans R Soc Trop Med Hyg. 1996, 90: 621-624. 10.1016/S0035-9203(96)90408-3.
Drakeley CJ, Jawara M, Targett GA, Walraven G, Obisike U, Coleman R, Pinder M, Sutherland CJ: Addition of artesunate to chloroquine for treatment of Plasmodium falciparum malaria in Gambian children causes a significant but short-lived reduction in infectiousness for mosquitoes. Trop Med Int Health. 2004, 9: 53-61. 10.1046/j.1365-3156.2003.01169.x.
Dutta GP, Bajpai R, Vishwakarma RA: Artemisinin (qinghaosu) – a new gametocytocidal drug for malaria. Chemotherapy. 1989, 35: 200-207.
Targett G, Drakeley C, Jawara M, von Seidlein L, Coleman R, Deen J, Pinder M, Doherty T, Sutherland C, Walraven G, Milligan P: Artesunate reduces but does not prevent posttreatment transmission of Plasmodium falciparum to Anopheles gambiae. J Infect Dis. 2001, 183: 1254-1259. 10.1086/319689.
Kaushal DC, Carter R, Miller LH, Krishna G: Gametocytogenesis by malaria parasites in continuous culture. Nature. 1980, 286: 490-492.
Inselburg J: Stage-specific inhibitory effect of cyclic AMP on asexual maturation and gametocyte formation of Plasmodium falciparum. J Parasitol. 1983, 69: 592-597.
Trager W, Gill GS: Plasmodium falciparum gametocyte formation in vitro: its stimulation by phorbol diesters and by 8-bromo cyclic adenosine monophosphate. J Protozool. 1989, 36: 451-454.
Read LK, Mikkelsen RB: Comparison of adenylate cyclase and cAMP-dependent protein kinase in gametocytogenic and nongametocytogenic clones of Plasmodium falciparum. J Parasitol. 1991, 77: 346-352.
Dyer M, Day KP: Expression of Plasmodium falciparum trimeric G proteins and their involvement in switching to sexual development. Mol Biochem Parasitol. 2000, 110: 437-448. 10.1016/S0166-6851(00)00288-7.
Muhia DK, Swales CA, Eckstein-Ludwig U, Saran S, Polley SD, Kelly JM, Schaap P, Krishna S, Baker DA: Multiple splice variants encode a novel adenylyl cyclase of possible plastid origin expressed in the sexual stage of the malaria parasite Plasmodium falciparum. J Biol Chem. 2003, 278: 22014-22022. 10.1074/jbc.M301639200.
Li JL, Targett GA, Baker DA: Primary structure and sexual stage-specific expression of a LAMMER protein kinase of Plasmodium falciparum. Int J Parasitol. 2001, 31: 387-392. 10.1016/S0020-7519(01)00126-6.
Taylor LH, Walliker D, Read AF: Mixed-genotype infections of the rodent malaria Plasmodium chabaudi are more infectious to mosquitoes than single-genotype infections. Parasitology. 1997, 115: 121-132. 10.1017/S0031182097001145.
Thomson D: A research into the production, life and death of crescents in malignant tertian malaria in treated and untreated cases, by an enumerative method. Ann Trop Med Parasitol. 1911, 5: 57-82.
Williams JL: Stimulation of Plasmodium falciparum gametocytogenesis by conditioned medium from parasite cultures. Am J Trop Med Hyg. 1999, 60: 7-13.
Dyer M, Day KP: Regulation of the rate of asexual growth and commitment to sexual development by diffusible factors from in vitro cultures of Plasmodium falciparum. Am J Trop Med Hyg. 2003, 68: 403-409.
Schneweis S, Maier WA, Seitz HM: Haemolysis of infected erythrocytes – a trigger for formation of Plasmodium falciparum gametocytes?. Parasitol Res. 1991, 77: 458-460.
Stephens JWW, Christophers SR: The practical study of malaria. 1908, London: University Press of Liverpool
Burkot T, Williams JL, Schneider I: Infectivity to mosquitoes of Plasmodium falciparum clones grown in vitro from the same isolate. Trans R Soc Trop Med Hyg. 1984, 78: 339-341. 10.1016/0035-9203(84)90114-7.
Read AF, Narara A, Nee S, Keymer AE, Day KP: Gametocyte sex ratios as indirect measures of outcrossing rates in malaria. Parasitology. 1992, 104: 387-395.
Read AF, Anwar M, Shutler D, Nee S: Sex allocation and population-structure in malaria and parasitic protozoa. Proc R Soc Lond B Biol. 1995, 260: 359-363.
West SA, Reece SE, Read AF: Evolution of gametocyte sex ratios in malaria and related apicomplexan (protozoan) parasites. Trends Parasitol. 17: 525-531. 10.1016/S1471-4922(01)02058-X.
Pickering J, Read AF, Guerrero S, West SA: Sex ratio and virulence in two species of lizard malaria parasites. Evol Ecol Res. 2000, 2: 171-184.
Shutler D, Bennett GF, Mullie A: Sex proportions of Haemoproteus blood parasites and local mate competition. Proc Natl Acad Sci USA. 1995, 92: 6748-6752.
Schall JJ: The sex ratio of Plasmodium gametocytes. Parasitology. 1989, 98: 343-350.
Robert V, Sokhna CS, Rogier C, Ariey F, Trape J-F: Sex ratio of Plasmodium falciparum in inhabitants of Dielmo, Senegal. Parasitology. 2003, 127: 1-8. 10.1017/S0031182003003299.
Charnov EL: The theory of sex allocation. 1982, Princeton: University Press, Princeton
Paul REL, Packer MJ, Walmsley M, Lagog M, Ranford-Cartwright LC, Paru R, Day KP: Mating patterns in Papua New guinea. Science. 1995, 269: 1709-1711.
Ariey F, Chalvet W, Hommel D, Peneau C, Hulin A, Mercereau-Puijalon O, Duchemin JB, Sarthou JL, Reynes JM, Fandeur T: Plasmodium falciparum parasites in French Guiana: limited genetic diversity and high selfing rate. Am J Trop Med Hyg. 1999, 61: 978-985.
Paul REL, Brey PT, Robert V: Plasmodium sex determination and transmission to mosquitoes. Trends Parasitol. 2002, 18: 32-38. 10.1016/S1471-4922(01)02122-5.
Paul REL, Coulson TN, Raibaud A, Brey PT: Sex determination in malaria parasites. Science. 2000, 287: 128-131. 10.1126/science.287.5450.128.
Taylor PJ, Hurd H: The influence of host hematocrit on the blood feeding success of Anopheles Stephensi : implication for enhanced malaria transmission. Parasitology. 2001, 122: 491-496. 10.1017/S0031182001007776.
Reece SE, Duncan AB, West SA, Read AF: Sex ratios in the rodent malaria parasite, Plasmodium chabaudi. Parasitology. 2003, 127: 419-425. 10.1017/S0031182003004013.
Osgood SM, Eisen RJ, Wargo AR, Schall JJ: Manipulation of the vertebrate host's testosterone does not affect gametocyte sex ratio of a malaria parasite. J Parasitol. 2003, 89: 190-192.
Smith TG, Walliker D, Ranford-Cartwright LC: Sexual differentiation and sex determination in the Apicomplexa. Trends Parasitol. 2002, 18: 315-323. 10.1016/S1471-4922(02)02292-4.
Kondrashov AS: Classification of hypotheses on the advantage of amphimixis. J Hered. 1993, 84: 372-387.
Agrawal AF: Sexual selection and the maintenance of sexual reproduction. Nature. 2001, 411: 692-695. 10.1038/35079590.
Butlin R: Opinion-evolution of sex: The costs and benefits of sex: new insights from old asexual lineages. Nat Rev Genet. 2002, 3: 311-317. 10.1038/nrg749.
CoIegrave N: Sex releases the speed limit on evolution. Nature. 2002, 420: 664-666. 10.1038/nature01191.
Otto SP: The advantages of segregation and the evolution of sex. Genetics. 2003, 164: 1099-1118.
West SA, Lively CM, Read AF: A pluralist approach to sex and recombination. J Evol Biol. 1999, 12: 1003-1012. 10.1046/j.1420-9101.1999.00119.x.
Foth BJ, McFadden Gl: The apicoplast: a plastid in Plasmodium falciparum and other Apicomplexan parasites. Int Rev Cytol. 2003, 224: 57-110.
Creasey AM, Ranford-Cartwright LC, Moore DJ, Williamson DH, Wilson RJ, Walliker D, Carter R: Uniparental inheritance of the mitochondrial gene cytochrome b in Plasmodium falciparum. Curr Genet. 1993, 23: 360-364.
Vaidya AB, Morrisey J, Plowe CV, Kaslow DC, Wellems TE: Unidirectional dominance of cytoplasmic inheritance in two genetic crosses of Plasmodium falciparum. Mol Cell Biol. 1993, 13: 7349-7357.
Walliker D, Quakyi IA, Wellems TE, McCutchan TF, Szarfman A, London WT, Corcoran LM, Burkot TR, Carter R: Genetic analysis of the human malaria parasite Plasmodium falciparum. Science. 1987, 236: 1661-1666.
Conway DJ, Roper C, Oduola AM, Arnot DE, Kremsner PG, Grobusch MP, Curtis CF, Greenwood BM: High recombination rate in natural populations of Plasmodium falciparum. Proc Natl Acad Sci U S A. 1999, 96: 4506-4511. 10.1073/pnas.96.8.4506.
Babiker HA, Ranford-Cartwright LC, Walliker D: Genetic structure and dynamics of Plasmodium falciparum infections in the Kilombero region of Tanzania. Trans R Soc Trop Med Hyg. 1999, 93 (Suppl 1): 11-14.
Kadekoppala M, Cheresh P, Catron D, Ji DD, Deitsch K, Wellems TE, Seifert HS, Haldar K: Rapid recombination among transfected plasmids, chimeric episome formation and trans gene expression in Plasmodium falciparum. Mol Biochem Parasitol. 2001, 112: 211-218. 10.1016/S0166-6851(00)00368-6.
Rosenberg R, Rungsiwongse J, Kangsadalampai S, Sattabongkot J, Suwanabun N, Chaiyaroj SC, Mongkolsuk S: Random mating of natural Plasmodium populations demonstrated from individual oocysts. Mol Biochem Parasitol. 1992, 53: 129-133. 10.1016/0166-6851(92)90015-C.
Pfefferkorn LC, Pfefferkorn ER: Toxoplasma gondii : genetic recombination between drug resistant mutants. Exp Parasitol. 1980, 50: 305-316. 10.1016/0014-4894(80)90034-X.
Taylor LH, Read AF: Determinants of transmission success of individual clones from mixed-clone infections of the rodent malaria parasite, Plasmodium chabaudi. Int J Parasitol. 1998, 28: 719-725. 10.1016/S0020-7519(98)00032-0.
Arez AP, Pinto J, Palsson K, Snounou G, Jaenson TG, do Rosario VE: Transmission of mixed Plasmodium species and Plasmodium falciparum genotypes. Am J Trop Med Hyg. 2003, 68: 161-168.
McKenzie FE, Jeffery GM, Collins WE: Plasmodium malariae infection boosts Plasmodium falciparum gametocyte production. Am J Trop Med Hyg. 2002, 67: 411-414.
Graves PM, Burkot TR, Carter R, Cattani JA, Lagog M, Parker J, Brabin BJ, Gibson FD, Bradley DJ, Alpers MP: Measurement of malarial infectivity of human populations to mosquitoes in the Madang area, Papua, New Guinea. Parasitology. 1988, 96: 251-263.
Tchuinkam T, Mulder B, Dechering K, Stoffels H, Verhave JP, Cot M, Carnevale P, Meuwissen JH, Robert V: Experimental infections of Anopheles gambiae with Plasmodium falciparum of naturally infected gametocyte carriers in Cameroon: factors influencing the infectivity to mosquitoes. Trop Med Parasitol. 1993, 44: 271-276.
Sattabongkot J, Maneechai N, Rosenberg R: Plasmodium vivax : gametocyte infectivity of naturally infected Thai adults. Parasitology. 1991, 102: 27-31.
Dowling MAC, Shute GT: A comparative study of thick and thin films in the diagnosis of scanty malaria parasitemia. Bull World Health Organ. 1966, 34: 249-267.
Paul REL, Ariey F, Robert V: The evolutionary ecology of Plasmodium. Ecol Let. 2003, 6: 866-880. 10.1046/j.1461-0248.2003.00509.x.
Taylor LH, Read AF: Why so few transmission stages? Reproductive restraint by malaria parasites. Parasitol Today. 1997, 13: 135-140. 10.1016/S0169-4758(97)89810-9.
Bishop A: Problems concerned with gametogenesis in Haemosporidiidea, with particular reference to the genus Plasmodium. Parasitology. 1955, 45: 163-185.
McKenzie FE, Bossert WH: The optimal production of gametocytes by Plasmodium falciparum. J TheorBiol. 1998, 193: 419-428. 10.1006/jtbi.1998.0710.
Al-Olayan EM, Williams GT, Hurd H: Apoptosis in the malaria protozoan, Plasmodium berghei: a possible mechanism for limiting intensity of infection in the mosquito. Int J Parasitol. 2002, 32: 1133-1143. 10.1016/S0020-7519(02)00087-5.
Pichon G, Awono-Ambene HP, Robert V: High heterogeneity in the number of Plasmodium falciparum gametocytes in the bloodmeal of mosquitoes fed on the same host. Parasitology. 2000, 121: 115-120. 10.1017/S0031182099006277.
Gautret P, Miltgen F, Gantier JC, Chabaud AG, Landau I: Enhanced gametocyte formation by Plasmodium chabaudi in immature erythrocytes: pattern of production, sequestration, and infectivity to mosquitoes. J Parasitol. 1996, 82: 900-906.
Chardome M, Janssen PJ: Enquête sur l'incidence malarienne par la méthode dermique dans la région de Lubilash (Congo Beige). Ann Soc Belge Med Trop. 1952, 32: 209-211.
Boëte C, Paul REL, Koella JC: Reduced efficacy of the immune melanization response in mosquitoes infected by malaria parasites. Parasitology. 2002, 125: 93-98. 10.1017/S0031182002001944.
Sinden RE: Gametocytes and sexual development. In Malaria: parasite biology, pathogenesis, and protection. Edited by: Sherman IW. 1998, Washington DC: American Society for Microbiology Press, 25-48.
Sinden RE, Smalley ME: Gametocytogenesis of Plasmodium falciparum in vitro : the cell-cycle. Parasitology. 1979, 79: 277-296.
Acknowledgements
This work was partly financed by the IRD and Programs VIH/PAL and PAL+ (Transmission blocking immunity) from the Ministére de la Recherche, France. AMT received support from Fondation Jeunesse Internationale. The authors AMT, OD, FA and VR, belong to the Grand Programme Horizontal Anopheles of Institut Pasteur. We thank Rosemary Barraclough for insightful comments. We are grateful for the bloody contribution of our colleague Nicole Andriaholinirina to the realisation of Figure 1.
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Talman, A.M., Domarle, O., McKenzie, F.E. et al. Gametocytogenesis : the puberty of Plasmodium falciparum. Malar J 3, 24 (2004). https://doi.org/10.1186/1475-2875-3-24
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2875-3-24