
Abstract
The selection of point mutation at codon 164 (from isoleucine to leucine) of the dihydrofolate reductase (DHFR) enzyme in Plasmodium falciparum is associated with high sulfadoxine /pyrimethamine (SP) resistance. Using the yeast expression system that allows the detection of dhfr allele present at low level, the presence of this mutation had previously been reported between 1998–1999 in Muheza, Tanzania, an area of high SP resistance. Eighty five P. falciparum isolates, obtained from the same area between 2002 and 2003, were analysed for the presence of Leu-164 mutation, using standard protocol based on PCR-RFLP. None of the isolates had the Leu-164 mutation.
Similar content being viewed by others

Background
The antifolate combination sulfadoxine /pyrimethamine (SP) is currently the first line treatment for uncomplicated falciparum malaria in several African countries. Pyrimethamine (PM) and sulfadoxine (SD) are inhibitors of dihydrofolate reductase (DHFR) and dihydropteroate synthase (DHPS) respectively. The selection and rapid spread of resistance to SP, which in some parts of East Africa has reached a prevalence of more than 30% [1], has already seriously compromised the therapeutic usefulness of SP. A new antifolate combination of chlorproguanil (CPG) and dapsone (DDS), known as Lapdap™ [2, 3], has now been developed. CPG is metabolised in vivo to its active triazine metabolite, chlorcycloguanil (CCG), which, like PM, inhibits the DHFR enzyme. DDS, like SD, inhibits the activity of dhps. LapDap™ is effective in vivo and retains efficacy against SP resistant infections [1], justifying the use of this drug as a replacement for SP.
Resistance to SP in Africa is attributable to parasites that carry point mutations at codons 108, 51 and 59 of dhfr, and resistance is augmented by point mutations at codon 437 and/or 540 of the dhps gene [4]. Experience in South East Asia and South America shows that the continued use of SP will eventually select for the mutation at codon 164 (from isoleucine to leucine). Once this occurs in Africa, parasites will become highly resistant to SP. This study shows that the selection of this Leu-164 mutation will also render the new antifolate combination CPG/DDS ineffective [5]. Therefore, the occurrence in Africa of this mutation would compromise the useful therapeutic life (UTL) of the new antifolate combination CPG/DDS.
Although SP has been widely used in Africa, the Leu-164 mutation has not yet been reported using standard protocols for the detection of this point mutation in dhfr (PCR-RFLP and sequencing). For instance, a study carried out on isolates collected during CPG/DDS trial in 1999, in Usambara Mountains, Tanzania, did not detect the 164-Leu using PCR-RFLP [6]. However, by using the yeast complementation approach, which is based on the expression of plasmodium dhfr genes in yeast cells followed by the selection of cells expressing highly resistant alleles, Hastings and co-workers [7] have reported the presence of the Leu-164 mutation in three SP resistant isolates collected between 1998–1999, from Muheza, Tanzania – an area with a high level of resistance resistance to SP [1]. This finding indicates that parasites carrying dhfr alleles with the Leu-164 mutation exist in Africa. As a consequence, the UTL of the new combination CPG/DDS could be very short.
Although the level of SP resistance is high in Muheza, this drug is still used as the first line treatment because of the lack of affordable alternative antimalarials. Under these conditions, one would expect the selection and spread of parasites carrying the Leu-164 mutation under SP pressure, if this mutation conferred a biological advantage. Treatment-mediated selection of the mutation would be expected to raise the gene frequency to the point where the dhfr alleles can be detected in Muheza using standard protocols.
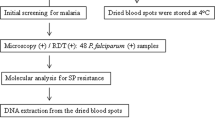
With this in mind, the presence of the Leu-164 mutation are being monitored in isolates collected at Muheza from 1999, the year the Leu-164 mutation was detected in this area, using the yeast cell complementation technique. In this paper, are presented the results of the analysis of 85 recent isolates collected randomly from children suffering from uncomplicated malaria at Muheza Designated District Hospital, Tanzania, between October 2002 to January 2003.
Materials and Methods
Patients were children under five years of age. Parent/caretakers had given written informed consent to participate in a separate on-going study assessing the effectiveness of combination therapies of SP+amodiaquine, artemether/lumefantrin and artesunate + amodiaquine. Approximately 50 μl of blood from the finger-prick, collected on day 0 before treatment, were spotted onto filter paper, air-dried and stored in plastic bags with silica gel at ambient temperature until PCR analysis. The preparation of parasite genomic material and the detection of point mutation at codon 164 of dhfr were carried out as described elsewhere [8]. Briefly, a small piece of blood-impregnated filter paper was snipped and put in a 0.5-ml microcentrifuge tube containing 150 μl of absolute methanol for 15 min at ambient temperature. Thereafter, the methanol was poured off and the paper was allowed to dry. Fifty microliters of sterilized water was then added to the tube, which was heated at 95°C for 15 min in a PCR machine. Following centrifugation (13,000 rpm [revolution per minute], 2 min), 2.5 to 5 μl of the solution was used as the template for PCR analysis.
Results and Discussion
The results show that none of these isolates had mutation at codon 164 of dhfr.
The number of isolates that were analysed is not representative of the whole population of the parasites in this area, but if this mutation is selected as the result of SP use, then 4 years after the detection of the Leu-164 mutation by yeast complementation technique, one might expect to detect it by PCR-RFLP in randomly selected isolates: this is not the case. In a similar study carried out recently on 277 isolates collected from several malaria endemic areas in Africa, the Leu-164 mutation was not detected using the standard PCR-RFLP protocol, in agreement with the findings reported in this paper [9].
A possible explanation of these observations is that the Leu-164 mutation detected by the yeast complementation techniques may only represent normal and baseline polymorphism of the dhfr gene, and that this mutation does not confer any selective advantage to the African parasite. Thus, the low fitness of DHFR enzyme carrying the Leu-164 mutation could explain the failure to select this mutation. Therefore, the selection of the Leu-164 mutation in South East Asia and South America may indicate that parasites from these areas developed compensatory mechanisms to make up for the low efficiency of DHFR enzyme. One of the most interesting explanations for the selection of the Leu-164 mutation has come from a microsatellite study carried out on isolates from South East Asia. Contrary to the case with isolates carrying Asn 108, Ile 51 and Arg 59 mutations, those harboring the Leu-164 mutation have derived from a single origin. This "single parasite" has spread in South East Asia as a result of drug pressure [10]. If the African parasites that carry the Leu-164 mutation currently lack this putative compensatory mechanism, this would explain why the Leu-164 mutation has not yet been detected by standard methods. However, if such parasites were to be imported into Africa while SP or CPG/DDS is still in use, then the parasites would spread and render the antifolate antimalarial drugs ineffective. This is a dangerous scenario for Africa. The question then is: when, and how will such parasites be imported into Africa? And, perhaps, whether it is possible to prevent this importation? Whatever the mechanisms associated with the selection of Leu-164 mutation may be, continuous monitoring of this mutation should be carried out, especially in areas of high SP resistance, so as determine whether the prevalence of resistant dhfr alleles harboring this mutation is increasing.
References
Mutabingwa T, Nzila A, Mberu E, Nduati E, Winstanley P, Hills E, Watkins W: Chlorproguanil-dapsone for treatment of drug-resistant falciparum malaria in Tanzania. Lancet. 2001, 358: 1218-23. 10.1016/S0140-6736(01)06344-9.
Winstanley P: Chlorproguanil-dapsone (LAPDAP) for uncomplicated falciparum malaria. Trop Med Int Health. 2001, 6: 952-954. 10.1046/j.1365-3156.2001.00751.x.
Lang T, Greenwood B: The development of Lapdap, an affordable new treatment for malaria. Lancet Infect Dis. 2003, 3: 162-168. 10.1016/S1473-3099(03)00547-4.
Sibley CH, Hyde JE, Sims PF, Plowe CV, Kublin JG, Mberu EK, Cowman AF, Winstanley PA, Watkins WM, Nzila AM: Pyrimethamine-sulfadoxine resistance in Plasmodium falciparum: what next?. Trends Parasitol. 2001, 17: 582-528. 10.1016/S1471-4922(01)02085-2.
Watkins WM, Mberu EK, Winstanley PA, Plowe CV: The Efficacy of antifolate antimalarial combination in africa: a predictive model based on pharmacodynamic and pharmacokinetic analyses. Parasitol Today. 1997, 13: 459-464. 10.1016/S0169-4758(97)01124-1.
Curtis J, Maxwell CA, Msuya FH, Mkongewa S, Alloueche A, Warhurst DC: Mutations in dhfr in Plasmodium falciparum infections selected by chlorproguanil-dapsone treatment. J Infect Dis. 2002, 186: 1861-4. 10.1086/345765.
Hastings MD, Bates SJ, Blackstone EA, Monks SM, Mutabingwa TK, Sibley CH: Highly pyrimethamine-resistant alleles of dihydrofolate reductase in isolates of Plasmodium falciparum from Tanzania. Trans R Soc Trop Med Hyg. 2002, 96: 674-6.
Nzila AM, Mberu EK, Sulo J, Dayo H, Winstanley PA, Sibley CH, Watkins WM: Towards an understanding of the mechanism of pyrimethamine-sulfadoxine resistance in Plasmodium falciparum: genotyping of dihydrofolate reductase and dihydropteroate synthase of Kenyan parasites. Antimicrob Agents Chemother. 2000, 44: 991-996. 10.1128/AAC.44.4.991-996.2000.
Wichmann O, Jelinek T, Peyerl-Hoffmann G, Muhlberger N, Grobusch MP, Gascon J, Matteelli A, Hatz C, Laferl H, Schulze M, Burchard G, Cunha Sd S, Beran J, McWhinney P, Kollaritsch H, Kern P, Cuadros J, Alifrangis M, Gjorup I: Molecular surveillance of the antifolate-resistant mutation I164L in imported african isolates of Plasmodium falciparum in Europe: sentinel data from TropNetEurop. Malar J. 2003, 2: 17-10.1186/1475-2875-2-17.
Nair S, Williams JT, Brockman A, Paiphun L, Mayxay M, Newton PN, Guthmann JP, Smithuis FM, Hien TT, White NJ, Nosten F, Anderson TJ: A selective sweep driven by pyrimethamine treatment in South East Asian malaria parasites. Mol Biol Evol. 2003, 20: 1526-36. 10.1093/molbev/msg162.
Acknowledgments
This work was supported by the Wellcome Trust of Great Britain (grant ref. 056769 and 062372) and the National Institute of Health of the U.S.A. (NIH Fogarty International grant TW 01186). AN, WW and KM are grateful to the Wellcome Trust for personal support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contribution
KM, GK and WW designed the study and TM carried out the collection of samples. EO, SK carried out the molecular analyses under the supervision of AN.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Ochong, E., Nzila, A., Kimani, S. et al. Molecular monitoring of the Leu-164 mutation of dihydrofolate reductase in a highly sulfadoxine/pyrimethamine-resistant area in Africa. Malar J 2, 46 (2003). https://doi.org/10.1186/1475-2875-2-46
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2875-2-46