
Abstract
Background
Applications of bacteriocins as food preservatives have been so far limited, principally because of their low antimicrobial activity in foods. Nisin is the only bacteriocin of significant use, but applications are restricted principally because of its very low activity at neutral or alkaline pH. Thus the isolation of new bacteriocins active in foods is desirable.
Results
We isolated a Bacillus licheniformis thermophilic strain producing a bacteriocin with some novel features, named here bacillocin 490. This bacteriocin was inactivated by pronase E and proteinase K and was active against closely related Bacillus spp. both in aerobic and in anaerobic conditions. Bactericidal activity was kept during storage at 4°C and was remarkably stable in a wide pH range. The bacteriocin was partially purified by elution after adhesion to cells of the food-isolated strain Bacillus smithii and had a rather low mass (2 KDa). Antimicrobial activity against B. smithii was observed also when this organism was grown in water buffalo milk.
Conclusions
Bacillocin 490 is a novel candidate as a food anti-microbial agent since it displays its activity in milk, is stable to heat treatment and during storage, is active in a wide pH range and has bactericidal activity also at high temperature. These features may allow the use of bacillocin 490 during processes performed at high temperature and as a complementary antimicrobial agent of nisin against some Bacillus spp. in non-acidic foods. The small size suggests its use on solid foods.
Similar content being viewed by others

Background
Bacteriocins are ribosomally synthesized antimicrobial peptides produced by a number of different bacteria [1], whose activity is generally directed towards species that are closely related or share the same ecological niche [2, 3]. Bacteriocins produced by Gram-positive bacteria have attracted much attention because of their potential use as food preservatives [4]. Many bacteriocins from Gram-positive bacteria, some of which produced by Bacillus spp., have been isolated and studied [5–13]. Nisin, a bacteriocin secreted by Lactococcus lactis, is commercially used as a food preservative in forty Countries since 1983 [14]. Like the lactic acid bacteria (LAB), that are the most studied bacteriocin-producing microorganisms, some representatives of Bacillus spp., such as B. subtilis and B. licheniformis, are "generally recognized as safe" (GRAS) bacteria [15].
A wider use of bacteriocins as antimicrobial agents in milk and dairy foods has been hindered so far by the low activity of many known bacteriocins in foods. In this study we report the isolation and partial characterization of a new bacteriocin with promising properties as an antimicrobial agent against Bacillus spp. frequently found in non-acidic foods.
Results
Approximately 23,000 bacterial isolates deriving from dairy foods were tested for their ability to produce antimicrobial activity against Gram-positive and Gram-negative species. We found that 0.48% of tested isolates produce antimicrobial activity against Salmonella, while a higher percentage (1.41%) produce anti-Listeria activity. The proteinaceous nature of the antimicrobial products was assessed by sensitivity to protease. During the screening, we isolated a strain showing high antimicrobial activity against some adjacent colonies. The producer strain was identified as Bacillus licheniformis based on standard phenotypic and metabolic characterization, and 16S ribosomal RNA gene sequence, and named Bacillus licheniformis 490/5. The bacteriocin produced, called here bacillocin 490, was completely inactivated by 1 mg of pronase E and proteinase K ml-1 but not by the same amount of trypsin or chemotrypsin.
B. licheniformis 490/5 was shown to be thermophilic with optimal growth rate at 65°C. Its growth was inhibited by a commercial bacitracin preparation at concentrations that kill the sensitive species Staphylococcus epidermidis but not the resistant one Escherichia coli.
The activity range of bacillocin 490 (Table 1) was rather narrow and limited to some Gram-positive bacteria. Highest antimicrobial activity was against B. stearothermophilus, B. smithii, B. subtilis and B. anthracis. We also observed some inhibition of B. cereus and B. licheniformis, very low inhibition of Listeria innocua and S. aureus and no inhibition of B. thuringensis and Streptococcus thermophilus. This activity spectrum clearly shows that bacillocin 490 is active principally against species phylogenetically related to the producer strain.
During growth the peak of bacteriocin production occurred in the late logaritmic phase and optimal production was obtained with resting cells at 55°C. Bacteriocin production was increased by 58% and 56% in the presence of 1% (w/v) skimmed milk and gelatin, respectively. This latter result is in agreement with previous observations that, in some cases, solid or semi-solid media (supplemented with gelling agents) increase significantly the amount of bacteriocin production [16–18].
Incubation of B. smithii (106 bacteria per ml) in the presence of bacillocin 490 (1 μg ml-1) at 60°C resulted in 96% killing in 30 minutes, indicating that the bacteriocin has a bactericidal effect.
Bacillocin 490 was purified taking advantage of its ability to bind to the sensitive cells of B. smithii. After elution of bound proteins with phosphate buffer, pH 4.5 (see Materials and Methods), a single 2 kDa band showing bactericidal activity was obtained by 10–25% acrylamide SDS-PAGE electrophoresis (data not shown) with a very high yield (60%) and increased specific activity (from 33.7 to 237.8 AU per mg of total proteins) with a purification factor of approximately 7. The latter low value is probably explained by small amount of protein secreted and is in agreement with other bacteriocin purifications [19]. The apparent weight of the bacteriocin was determined by gel-filtration and resulted approximately 2 kDa in agreement with electrophoretic data.
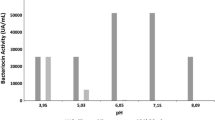
Purified bacillocin 490 was partially characterized. It was equally active both in the presence and in the absence of oxygen, showed great stability in a wide pH range, since it had optimum activity at neutral and moderately basic pH and maintained some 60% of activity after incubation at pH 3.5 and 11.0, with 95–100% of activity detected rather uniformly from pH 4.5 to 9.0. This is of great advantage from the industrial point of view, because the pH of many foods varies from very acidic to basic. In this connection it is interesting to notice that nisin, the only bacteriocin so far employed commercially as food additive against Gram-positive bacteria, acts in acidic conditions, as its maximum stability is at pH 2.0, and it is pratically not soluble at pH 8.0 [20].
Bacillocin 490 showed high stability at 100°C with 46.4% residual activity after 1 hour of exposure at this temperature (Fig. 1). It also showed remarkable stability at 4°C (no loss of activity after 12-month storage).

Thermal resistance of bacillocin 490 at 100°C. Other experimental details are in the Materials and Methods.
Bacillocin 490 was also used in preliminary experiments as an antimicrobial agent against the most sensitive species, B. smithii, inoculated in sterile water buffalo milk. Bacillocin 490 was not affected by protein aggregation since no difference of stability was observed when the bacteriocin was dissolved in milk and in buffer. As shown in Fig. 2, we observed 50% growth inhibition of B. smithii, after 5-h incubation in milk in the presence of 65 ng of bacillocin 490 ml-1. This result indicates that this bacteriocin is suitable for milk-based foods.

Survival of Bacillus smithii in milk at 25°C in the absence (open circles) and in the presence (bold circles) of bacillocin 490. See Materials and Methods for experimental details.
Two other bacteriocins found in B. licheniformis isolates have been described recently. One, named by the authors Lichenin, displays its activity only in strictly anaerobic conditions and against anaerobic organisms [21]. The other, named BSCY2, has a higher molecular mass with respect to both bacillocin 490 and Lichenin, and is moderately heat stable [22].
Conclusions
We report here the identification, purification and partial characterization of bacillocin 490, a B. licheniformis bacteriocin with very promising features for industrial applications. It displayed bactericidal activity in milk as well as in synthetic media, showed remarkable stability to heat and during storage and was active up to 60°C in a wide pH range. The latter feature may allow the use of bacillocin 490 at high temperature and as a complementary antimicrobial agent of nisin against some Bacillus spp. in non-acidic foods. The small size of bacillocin 490 suggests its employment on solid foods.
Materials and Methods
Bacterial strains and growth conditions
Bacterial strains used are those listed in Table 1, with the addition of B. licheniformis 490/5, the bacteriocin producer. All strains, except S. thermophilus, were cultured aerobically in TY medium (16 g tryptone l-1, 10 g yeast extract l-1, 5 g sodium chloride l-1). S. thermophilus was growth anaerobically using GasPack apparatus (OXOID, Basingstoke, UK) in HJL broth (3% tryptone, 1% yeast extract, 1% lactose, 0.2% beef extract, 0.5% KH2PO4, pH 6.5) and plated on solid M17 agar containing 1% lactose.
Solid and soft media were prepared by adding 20 g/liter and 7 g/liter of agar to the broth, respectively.
Bacillus licheniformis 490/5 and Bacillus smithii PRO/S were isolated from dairy products at 65°C as adjacent colonies on the same plate. Taxonomic classification of both strains was performed by use of API 50 CHB fermentation strips (bioMérieux, Marcy l'Etoile, France) and by sequence analysis of one of the 16S rRNA genes amplified with two oligonucleotides: P1 (5'-GCGGCGTGCCT AATACATGC) and P2 (5'-CACCTTCCGATACGGCTACC), annealing to nucleotides 40 to 59 and 1532 to 1513, respectively, of B. subtilis rrnE.
Screening of bacteria producing antimicrobial activities
Milk and dairy products were homogenized, diluted and plated onto various selective and non selective media (PCA, M17, MRS) in different growth conditions (varying temperature and oxygen supply). After incubation, individual clones were replicated in duplicate plates, incubated and, after appearance of colonies, one set of plates was u.v.254nm irradiated for 15 min to kill the cells and overlaid with 5 ml of 50°C – heated soft agar suspension containing 100 μl of an exponentially growing bacterial culture (106 cells per ml) of an indicator strain. Plates were incubated at 37°C for 18 h and inhibition zones were measured. Corresponding producer strains were isolated from the non-irradiated set of plates.
Bacteriocin production and detection
Bacillus licheniformis 490/5 was grown at 55°C in TY broth to stationary phase, and the culture was centrifuged at 2,500×g for 10 min. 20 μl of cell-free filter-sterilized supernatant were spotted on a sterile susceptibility blank discs (OXOID Basingstoke, UK) placed onto a plate filled with 5 ml of solidified soft agar containing 106 cells of Bacillus smithii (in 100 μl) used as indicator strain for detection of bacteriocin activity. The plate was incubated at 55°C for 18 h in the presence and in the absence of oxygen (the latter condition was accomplished by incubating the plates in GasPack apparatus) and inhibition areas were measured using a GelDoc 2000 photographic apparatus (BioRad) equipped with MultyAnalyst software (BioRad). One Arbitrary Unit (AU) was defined as the amount of bacteriocin producing 1 mm2 of inhibition area around the sterile disk.
Bacteriocin characterization
To evaluate thermal stability, partially purified bacteriocin was heated at 100°C or exposed at 4°C. Samples were collected and the activity measured at various times and reported as % of values of untreated samples.
Bacteriocin was also treated with pronase E, trypsin, α-chemotrypsin, proteinase K (all purchased from Sigma Chemical Co.). Enzymes were filter-sterilized and added to crude bacteriocin preparations at final concentration of 1 mg/ml. Following incubation at 37°C for 2 h, enzymes were denatured by heating at 100°C for 5 min. Untreated samples incubated in the same conditions were used as controls. Residual activity of samples was determined as mentioned above.
Purification
The bacteriocin was partially purified from 50 ml cultures of Bacillus licheniformis 490/5. Cells were grown to stationary phase in TY broth at 55°C and then collected by centrifugation (2,500 × g at 4°C, 10 min). The supernatant was filter-sterilized, ultrafiltered on 1,000 cut-off membrane to concentrate 10-fold and finally mixed with a pellet of pre-washed B. smithii cells from 1 liter of an 18 h culture at 55°C. The suspension was gently up-down mixed for 1 h to favour adhesion of the bacteriocin to the cell walls, and then centrifuged 5 min at 700 × g. The supernatant containing unbound proteins was discarded and cells were washed 3 times with 5 ml of 50 mM sodium phosphate buffer, pH 7.0. Finally, bound bacteriocin was eluted with the same buffer at pH 4.5.
SDS-polyacrylamide gel electrophoresis
Electrophoresis was performed by standard protocols [23]. For detection of bacteriocin activity after gel separation a previously reported method was used [9].
Gel-filtration
To determine its apparent molecular weight, the bacteriocin was loaded on a 1.3 ml G-75 Superdex (Pharmacia) gel filtration column, pre-equilibrated with the elution buffer (50 mM sodium phosphate, pH 7.0). Antimicrobial activity was assayed on each fraction.
Stability and antimicrobial activity in milk
For stability studies, equal amounts of bacillocin 490 were added to 10 ml of either 50 mM sodium phosphate buffer pH 7.0 or sterile fresh water buffalo milk and incubated at 25°C. 20 μl of samples were tested for antimicrobial activity each day.
To test the efficacy of bacteriocin against microorganisms present in milk, 10 ml of heat-sterilized fresh water buffalo milk containing 106 cells per ml of Bacillus smithii strain PRO/S were incubated 5 h at 25°C in the presence of 65 ng of bacteriocin ml-1. 100 μl samples were diluted in buffer and plated on TY medium each time to detect cell survival. A sample treated in the same conditions, without bacteriocin, was used as a control. Plates were incubated at 55°C.
References
Tagg JR, Dayani AS, Wannamaker LW: Bacteriocins of Gram-positive bacteria. Bacteriol Rev. 1976, 40: 722-756.
Klaenhammer TR: Bacteriocins of Lactic Acid Bacteria. Biochimie. 1988, 70: 337-349. 10.1016/0300-9084(88)90206-4
Klaenhammer TR: Genetics of bacteriocins produced by Lactic Acid Bacteria. FEMS Microbiol Rev. 1993, 12: 39-86. 10.1016/0168-6445(93)90057-G
Gould G: Industry perspectives on the use of natural antimicrobial and inhibitors for food applications. J Food Prot. 1996, 82-86. Suppl
Favret ME, Yousten AA: Thuricin: the bacteriocin produced by Bacillus thuringensis. J Invertebr Pathol. 1989, 53: 206-216.
Hyronimus B, Le Marrec C, Urdaci MC: Coagulin, a bacteriocin-like inhibitory substance produced by Bacillus coagulans I4. J Appl Microbiol. 1998, 85: 42-50. 10.1046/j.1365-2672.1998.00466.x
Klein C, Entian KD: Genes involved in self-protection against the lantibiotic subtilin produced by Bacillus subtilis ATCC 6633. Appl Environ Microbiol. 1994, 60: 2793-2801.
Le Marrec C, Hyronimus B, Bressollier P, Verneuil B, Urdaci MC: Biochemical and genetic characterization of Coagulin, a new antilisterial bacteriocin in the pediocin family of bacteriocins, produced by Bacillus coagulans I4. Appl Environ Microbiol. 2000, 66: 5213-5220. 10.1128/AEM.66.12.5213-5220.2000
Naclerio G, Ricca E, Sacco M, De Felice M: Antimicrobial activity of a newly identified bacteriocin of Bacillus cereus. Appl Environ Microbiol. 1993, 59: 4313-4316.
Oscariz JC, Lasa I, Pisabarro AG: Detection and characterization of cerein 7, a new bacteriocin produced by Bacillus cereus with a broad spectrum of activity. FEMS Microbiol Letters. 1999, 178: 337-341. 10.1016/S0378-1097(99)00370-5
Piuri M, Sanchez-Rivas C, Ruzal SM: A novel antimicrobial activity of a Paenibacillus polymixa strain isolated from regional fermented sausages. Lett Appl Microbiol. 1998, 27: 9-13. 10.1046/j.1472-765X.1998.00374.x
Sthal S: A new bacteriocin activity: megacin BII encoded by plasmid PSE 203 in strains of Bacillus megaterium. Arch Microbiol. 1989, 151: 159-165.
Zheng G, Slavik MF: Isolation, partial purification and characterization of a bacteriocin produced by a newly isolated Bacillus subtilis strain. Lett Appl Microbiol. 1999, 28: 363-367. 10.1046/j.1365-2672.1999.00545.x
Hoover DG, Steenson LR: Bacteriocins of Lactic Acid Bacteria. Academic Press. 1993
Sharp RJ, Scawen MD, Atkinson T: Fermentation and downstream processing of Bacillus. In Biotechnology Handbook: Bacillus. Edited by: CR Harwood. 1989, 255-292. New York, Plenum Press,
Kelstrup J, Gibbons RS: Bacteriocins from human and rodent Streptococci. Arch Oral Biol. 1969, 14: 251-258.
Setten AM, Vogels GD, De Windt F: Production and purification of a Staphylococcus epidermidis bacteriocin. J Bacteriol. 1972, 112: 235-242.
Piard J-C, Muriana PH, Desmazeaud MJ, Klaenhammer TR: Purification and partial characterization of Lactacin 481, a lanthionine-containing bacteriocin produced by Lactococcus lactis subs. lactis CNRZ 481. Appl Environ Microbiol. 1992, 58: 279-284.
Zheng G, Yan LZ, Vederasand JC, Zuber P: Genes of the SBO-ALB locus of Bacillus subtilis are required for production of the antilisterial bacteriocin subtilosin. J Bacteriol. 1999, 181: 7346-7355.
Liu W, Hansen JN: Some chemical and physical properties of nisin, a small-protein antibiotic produced by Lactococcus lactis. Appl Environ Microbiol. 1990, 56: 2551-2558.
Pattnaik P, Kaushik JK, Grover S, Batish VK: Purification and characterization of a bacteriocin-like compound (Lichenin) produced anaerobically by Bacillus licheniformis isolated from water buffalo. J Appl Microbiol. 2001, 91: 636-645. 10.1046/j.1365-2672.2001.01429.x
Chang J-Y, Lee H-H, Kim I-C, Chang H-C: Characterization of a bacteriocin produced by Bacillus licheniformis cy2. J. Korean Soc. Food Sci. Nutr. 2001, 30: 410-414.
Sambrook J, Fitsch EF, Maniatis T: Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, Cold Spring Harbor Press. 1989
Acknowledgements
This work belongs to the project "Biotecnologie Mediche ed Agro-alimentari" supported by "MIUR-Ministero dell'Istruzione, dell'Università e della Ricerca" and the "Consorzio Technapoli – Parco Scientifico e Tecnologico dell'Area Metropolitana di Napoli" under responsibility of "CEINGE", Naples, Italy.
The authors wish to thank "Bacillus Genetic Stock Center" that provided some Bacillus strains.
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Martirani, L., Varcamonti, M., Naclerio, G. et al. Purification and partial characterization of bacillocin 490, a novel bacteriocin produced by a thermophilic strain of Bacillus licheniformi s. Microb Cell Fact 1, 1 (2002). https://doi.org/10.1186/1475-2859-1-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2859-1-1