
Abstract
Background
High density lipoproteins (HDL) have many cardioprotective roles; however, in subjects with type 2 diabetes (T2D) these cardioprotective properties are diminished. Conversely, increased fruit and vegetable (F&V) intake may reduce cardiovascular disease risk, although direct trial evidence of a mechanism by which this occurs in subjects with T2D is lacking. Therefore, the aim of this study was to examine if increased F&V consumption influenced the carotenoid content and enzymes associated with the antioxidant properties of HDL in subjects with T2D.
Methods
Eighty obese subjects with T2D were randomised to a 1- or ≥6-portion/day F&V diet for 8-weeks. Fasting serum was collected pre- and post-intervention. HDL was subfractionated into HDL2 and HDL3 by rapid ultracentrifugation. Carotenoids were measured in serum, HDL2 and HDL3 by high performance liquid chromatography. The activity of paraoxonase-1 (PON-1) was measured in serum, HDL2 and HDL3 by a spectrophotometric assay, while the activity of lecithin cholesterol acyltransferase (LCAT) was measured in serum, HDL2 and HDL3 by a fluorometric assay.
Results
In the ≥6- vs. 1-portion post-intervention comparisons, carotenoids increased in serum, HDL2 and particularly HDL3, (α-carotene, p = 0.008; β-cryptoxanthin, p = 0.042; lutein, p = 0.012; lycopene, p = 0.016), as did the activities of PON-1 and LCAT in HDL3 (p = 0.006 and 0.044, respectively).
Conclusion
To our knowledge, this is the first study in subjects with T2D to demonstrate that increased F&V intake augmented the carotenoid content and influenced enzymes associated with the antioxidant properties of HDL. We suggest that these changes would enhance the cardioprotective properties of this lipoprotein.
Clinical trial registration
Similar content being viewed by others

Background
Cardiovascular disease (CVD) is the leading cause of morbidity and mortality in Western societies, whose incidence is augmented by T2D [1]. In 2008, diabetes was the direct cause of 17% of all CVD related deaths [2]. Conversely, high density lipoproteins (HDL) are normally associated with reduced CVD risk, as these lipoproteins have many antiatherogenic properties; including their antioxidant and reverse cholesterol transport (RCT) capabilities. Furthermore, HDL stimulates glucose uptake and fatty acid oxidation, opposing insulin resistance [3]. However, in subjects with T2D the function of HDL may be defective, as hyperglycaemia increases lipid peroxidation [4], while the release of the inflammatory marker serum amyloid A (SAA) from hypertrophic adipocytes also impacts on the anti-inflammatory and antioxidant properties of HDL [5]. Thus the main enzymes associated with HDL’s antioxidant function, paraoxonase-1 (PON-1) and lecithin cholesterol acyltransferase (LCAT), with this latter enzyme also being involved in the maturation of HDL, may be altered to a pro-atherogenic phenotype in dysfunctional HDL [6, 7].
Therefore, interventions that reduce the risk of diabetes and CVD, such as increased fruit and vegetable (F&V) intake, are highly desirable. Although the relationship between F&V intake and the incidence of T2D is not fully understood, there is substantial support for increased intake to reduce diabetes and CVD risk. In this regard, the study of Villegas et al. [8] reported that vegetables were protective against the development of T2D, while, in a population without diabetes, Panagiotakos et al. [9] identified that subjects consuming vegetables for more than 3 days per week had a 70% decreased risk of CVD and, furthermore, that an increase of one portion of fruit per day decreased CVD risk by 10%. In support of this, Cooper et al., [10] recently showed that increased vegetable intake and a variety of fruit reduced diabetes risk. Therefore, from the above evidence, it appears that increased F&V intake would be protective against the development of diabetes and the associated CVD complications.
The protective effect of F&V may be related to their rich antioxidant content, as these have been linked to a lower diabetes risk and the suppression of the inflammatory response related to diabetes and CVD development [11, 12]. Many F&V derived lipid phase antioxidants, such as the carotenoids, are transported in the circulation by lipoproteins, including HDL, and in the case of lycopene, has been shown to influence the cardioprotective properties of HDL. In fact, in a previous study by our group we found that among middle-aged, moderately overweight subjects, lycopene intake, via supplement or as a lycopene-rich diet, lead to antiatherogenic increases in the activities of PON-1 and LCAT within HDL [11]. Additionally, another study identified that serum carotenoids and the activity of serum-PON-1 increased following a Mediterranean-like diet, while the inflammatory marker C-reactive protein (CRP) decreased [13]. Overall, these studies suggest that the antioxidants within F&V may, in part, be responsible for their cardioprotective effects.
However, although the literature is suggestive that increased F&V intake is beneficial to subjects with T2D, no study has investigated the relationship between increased F&V intake and the enzymes associated with HDL in these subjects. Therefore, the aim of this study was to examine if increased F&V intake had the potential to enhance the antiatherogenic properties of this lipoprotein in subjects with T2D.
Methods
Study population
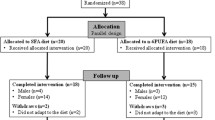
This study was a secondary analysis of blood samples collected from subjects with T2D following a randomised controlled F&V intervention. The study consisted of 80 subjects who were recruited from the diabetes outpatient clinics at the Regional Centre for Endocrinology and Diabetes at the Royal Victoria Hospital, Belfast, UK. All subjects were obese (BMI > 30 kg/m2), and aged between 40 and 70 years. In each case T2D was controlled by diet or oral hypoglycaemic therapy, which was maintained for the duration of the study. Prior to the 8-week intervention period all subjects underwent a 4-week run-in period, where they were asked to consume 1-portion of F&V/day. One portion of F&V is defined by the Food Standards Agency and the National Health Service as 80 g [14]. Following this, subjects were randomly assigned to 1 of 2 treatment groups (n = 40/group): a low F&V group, where they continued to consume 1-portion of F&V/day, or a high F&V group, were they consumed ≥6-portions of F&V/day. Fasting blood samples were obtained following the wash-out phase (week-0) and post-intervention (week-8), serum was isolated by centrifugation and stored in aliquots at -80°C until required for analysis. Figure 1 illustrates the study design for this intervention. F&V supplementation was similar to that of a normal diet, therefore, did not pose any health risk. Specific advice was also given to ensure a similar calorific and macronutrient intake to their normal diet, while weekly contact encouraged compliance. To assess compliance, two 7-day food diaries were completed during the run-in period and during the intervention (results not shown). Subjects selected the F&Vs of their choice, which were delivered weekly from a local supermarket during the 8-week intervention period. They were also offered advice on suitable storage and cooking methods for the F&Vs to minimise their degradation. This study was approved by the ethics committee of Queen’s University Belfast, and all subjects gave written informed consent.

Study Design.
Exclusion criteria
Exclusion criteria included any acute coronary/cerebrovascular event, surgery within the previous 3-months; pregnant or lactating females; oral anticoagulant therapy; excess alcohol consumption (>2 units per day for females; >3 units per day for males); food sensitivities that would interfere with the F&V consumption; medical conditions or dietary restrictions that would substantially limit the ability of the subject to complete the study requirements; and ingestion of oral vitamin/antioxidant supplements in the 4-weeks prior to the wash-out phase.
Isolation of HDL2 and HDL3 from serum
HDL2 and HDL3 were harvested from freshly thawed serum by rapid ultracentrifugation, according to the method of McPherson et al., (2007) [15]. This consisted of a 3-step, 6-hour long procedure. HDL samples were stored at -80°C and at this temperature HDL is stable for up to one year (results not shown), all samples were batch analysed.
Apolipoprotein (apo) AI concentration
Apo AI was determined by a single radial immunodiffusion method, as previously described [16].
Carotenoid concentration
Lipid soluble carotenoids, including α-carotene, β-carotene, β-cryptoxanthin, lutein, lycopene and zeaxanthin were measured in serum, HDL2 and HDL3 by reverse-phase-high performance lipid chromatography (HPLC) with diode array detection, as previously described [17].
PON-1 activity
The arylesterase activity of PON-1 was measured in serum, HDL2 and HDL3. This assay was based upon the ability of PON-1 to hydrolyse phenylacetate, and was an adaption of the method of Hasselwander et al., 1999 [18], utilising 5 μL of serum, 200 μL of HDL2 and 20 μL of HDL3. One unit of arylesterase activity (U) is defined as 1 μmol of phenol generated per minute [18].
LCAT activity
The activity of LCAT was measured in serum, HDL2 and HDL3 using a commercially available fluorometric assay (RB-LCAT, ROAR Biomedical, NY, USA), as per manufacturer’s instructions. The LCAT assay required 5 μL of serum, HDL2 or HDL3 per analysis.
hsCRP and SAA concentration
The concentration of serum high sensitive (hs)CRP was determined by immunoturbidimetry. While SAA was measured in serum, HDL2 and HDL3 by a commercially available ELISA (Invitrogen, Human SAA KHA0011C) and the analysis was performed on a Grifols TRITURUS automated ELISA system (Italy), as per the manufacturer’s instructions, with the following modifications prior to analysis: serum was diluted 1:150, HDL2 1:10 and HDL3 1:100.
Statistical analysis
A retrospective power calculation for HDL3-lycopene and HDL3-PON-1 activity indicated that a subject population of 20 per group was sufficient to give a 90% power at the 5% level of significance to detect a statistical difference between the groups. However, for completeness we have included all subjects in our analyses. Statistical analyses were performed using SPSS Statistics version 17.0. Variables were assessed for normality and logarithmically transformed where required. Between-group comparisons were analysed by an independent samples t-test. Within-group comparisons were analysed by a paired sample t-test. To adjust for potential confounders, an analysis of covariance was conducted, with final value of the variable of interest included as the dependent variable, the initial value of the variable of interest included as the covariate, and potential confounders, such as BMI, included in the model. All variables were summarised as mean (SEM) when normally distributed and as geometric mean (interquartile range) when normally distributed after logarithmic transformation. Correlations between variables were assessed by Pearson’s two-tailed bivariate analysis. Significance was set as p < 0.05 for all analyses.
Results
Subject characteristics
Initially 98 subjects were recruited into the study, 17 subjects withdrew during the run-in period prior to randomisation and 1 subject withdrew during the intervention period due to a family bereavement. A total of 80 subjects with T2D completed the 8-week intervention study. Six subjects had either pre- or post-intervention serum hsCRP levels >10 mg/l and thus were excluded from further analyses. This was in accordance with American Heart Association guidelines [19] as a hsCRP of >10 mg/l is taken as evidence of active infection or inflammation. Thus 74 subjects were included in the final analyses (Table 1). All baseline characteristics were comparable between the groups (p > 0.05 for all comparisons), and although there were more women randomised to the ≥6-portion intervention (n = 15/39) compared to the number of women randomised to the 1-portion intervention (n = 7/35), this was not statistically different (p = 0.295). Following intervention, BMI was different between the groups, which was driven by the significant increase in BMI (0.3 kg/m2) following the ≥6-portion intervention (p = 0.001). However, the effect of this change in BMI was tested independently using an analysis of covariance for all subsequent statistical analyses and was found not to influence the results (data not shown). Additionally, HbA1C decreased following the 1-portion intervention (p = 0.032), although this was not continued in the between group comparisons (p > 0.05).
Fruit and vegetable consumption
F&V intake was calculated from two 7-day food diaries, where it was identified that F&V intake was significantly different in the ≥6- vs. 1-portion comparisons (p ≤ 0.001) following intervention, which was driven by the significant increase in F&V intake following the ≥6-portion intervention [pre 1.4 (0.5) portions F&V/day vs. post 5.2 (1.4) portions F&V/day, p ≤ 0.001], which equated to a 3.8 (1.34) portion increase in F&V/day, while F&V intake did not change following the 1-portion intervention [pre 1.2 (0.4) portions F&V/day vs. post 1.5 (0.9) portions F&V/day, p = 0.335].
Dietary intake
Following intervention, total energy was not different in the ≥6- vs. 1-portion comparisons (p > 0.05; Table 2). However, carbohydrate and total sugar were significantly different between the groups (p < 0.05 and p ≤ 0.001, respectively), which were driven by the significant increase following the ≥6-portion intervention (p < 0.05 and p ≤ 0.001, respectively).
Apo AI concentration
Following intervention, apo AI was not different in the between or within group comparisons (p > 0.05 for all comparisons, results not shown).
Carotenoid concentration
Following intervention, serum α-carotene, β-cryptoxanthin, lutein and zeaxanthin increased in the ≥6- vs. 1-portion comparisons (p < 0.05 for all comparisons); Table 3) which were driven by increases of ~8% in α-carotene (p = 0.036), ~25% β-cryptoxanthin (p = 0.001) and ~25% lutein (p = 0.003), and the borderline non-significant increase in zeaxanthin (p = 0.067) following the ≥6-portion intervention. In HDL2, β-cryptoxanthin increased in the ≥6- vs. 1-portion post-intervention comparisons (p < 0.05), which was driven by the ~47% increase in β-cryptoxanthin (p = 0.015) following the ≥6-portion intervention, additionally, lutein also increased in HDL2 following the ≥6-portion intervention (p ≤ 0.001). For HDL3, α-carotene, β-cryptoxanthin, lutein and lycopene increased in the ≥6- vs. 1-portion post-intervention comparisons (p < 0.05 for all comparisons), which were driven by an increase of ~38% α-carotene (p = 0.001), ~32% β-cryptoxanthin (p = 0.041), ~9% lutein (p = 0.006) and ~27% lycopene (p = 0.015) following the ≥6-portion intervention. Additionally, following the ≥6-portion intervention there was a ~30% increase in β-carotene (p = 0.012) and a ~11% increase in zeaxanthin (p = 0.004).
PON-1 activity
Following intervention, serum-PON-1 activity was unchanged in the ≥6- vs. 1-portion comparisons (p > 0.05; Table 4), however, serum-PON-1 activity increased by ~7% (p ≥ 0.001) following the ≥6-portion intervention. In HDL2, PON-1 activity was unchanged in the ≥6- vs. 1-portion comparisons (p < 0.05) and following the 1 and ≥6-portion interventions (p > 0.05 for both comparisons). However, in HDL3, the activity of PON-1 significantly increased in the ≥6- vs. 1-portion comparison (p < 0.05), which was driven by the ~13% increase in its activity following the ≥6-portion intervention (p ≤ 0.001), despite the ~4% increase in its activity following the 1-portion intervention (p = 0.014).
LCAT activity
In serum, LCAT activity was unchanged in the ≥6- vs. 1-portion comparisons (p > 0.05; Table 4). However, similarly to PON-1, the activity of serum-LCAT increased by ~8% following the ≥6-portion intervention (p = 0.025). In HDL2, the activity of LCAT was unchanged in the ≥6- vs. 1-portion comparisons (p > 0.05) and following the 1 and ≥6-portion interventions (p > 0.05 for both comparisons). However, the activity of LCAT was increased in HDL3 in the ≥6- vs. 1-portion comparison (p < 0.05), which was driven by the ~8% increase in its activity following the ≥6-portion intervention (p < 0.001).
hsCRP and SAA concentration
Following intervention, serum hsCRP and serum-, HDL2- and HDL3-SAA were similar in the ≥6- vs. 1-portion comparisons (p < 0.05 for all comparisons; Table 4) and although SAA decreased by ~32% following the ≥6-portion intervention in HDL3, this was not significant (p = 0.133).
Correlations between change in F&V intake, change in carotenoid antioxidant levels and change in activity of HDL-associated enzymes
Change in F&V intake was positively correlated with change in HDL2-β-cryptoxanthin (r = 0.312, p = 0.047), HDL3 α-carotene (r = 0.416, p = 0.005), HDL3 β-cryptoxanthin (r = 0.325, p = 0.032) and HDL3 lutein (r = 0.317, p = 0.036). Additionally, change in HDL3 β-cryptoxanthin positively correlated with change in HDL3-PON-1 activity (r = 0.258, p = 0.043).
Discussion
Carotenoids and the influence of increased F&V intake
This study has demonstrated that increased F&V intake augmented serum carotenoid levels in this subject population, illustrating subject compliance and confirming previous reports in healthy and T2D subjects [20]. In addition, we have shown for the first time in a population with T2D that this increase led to a concomitant increase in the carotenoid content of HDL2 and HDL3, which was particularly apparent for α-carotene, β-cryptoxanthin, lutein and lycopene in HDL3. Furthermore, this lycopene result confirms our previous findings, where we identified that lycopene, which is the most potent single oxygen quencher among the natural carotenoids [21], increased in HDL2 and HDL3 following a lycopene rich diet [11]. Thus we suggest that the increase in the carotenoids identified in this current study would enhance the antioxidant properties of HDL, similarly to that reported in subjects with T2D following a high polyphenol diet, where plasma malondialdehyde, a frequently used indicator of lipid peroxidation [22] decreased in subjects consuming this diet [23]. Furthermore, in support of our findings phylloquinone, a lipid-soluble molecule that is found in green leafy plants, improves markers related to insulin resistance and diabetes [24] and, due to its ability to accept electrons, contributes to the antioxidant capacity of HDL, which we suggest may also be the case for the increased carotenoids identified in this current study. In addition, a Mediterranean-style diet has been suggested to be advantageous from a metabolic perspective in subjects with T2D [25] and may even modulate genetic predisposition in such subjects [26]. Thus, overall we suggest that the increased antioxidants within the HDL subfractions, particularly HDL3, the subfraction more closely associated with HDL’s antioxidant properties [27, 28], would have enhanced antiatherogenic potential in this T2D cohort.
PON-1 and LCAT activities and the influence of increased F&V intake
This F&V intervention was also associated with significant increases in the activities of both PON-1 and LCAT, again confirming our previous findings following a lycopene rich diet [11]. Therefore, as one of HDL’s main antiatherogenic function is to inactivate peroxidised lipids removed from apolipoprotein B-containing lipoproteins, due to the proton donating properties of PON-1 [29] and to a lesser extent LCAT [30], an increase in the activities of these enzymes would be thought of as antiatherogenic. Therefore, as the activity of serum-PON-1 has been reported to be lower in subjects with T2D [31], we have now shown that increased F&V intake augmented its activity, in serum and particularly in HDL3, in this T2D population, which was further supported by the positive correlation between the concentration of β-cryptoxanthin and the activity of PON-1 in HDL3 (r = 0.258, p = 0.043). These findings are in support of previous studies, where quercetin and glabridin were shown to protect PON-1 against copper-induced oxidation [29], while pomegranate juice (a rich source of carotenoids) was shown to increase its activity in healthy male subjects [32]. Overall, we suggest that the increase in the described antioxidants may have enhanced PON-1’s proton donating properties and hence were responsible for the increase in its activity, especially in HDL3. We suggest that this increase in PON-1 activity would enhance the antiatherogenic potential of HDL in this T2D population.
With regards to LCAT, similarly to PON-1, this enzyme is mainly associated with HDL3, and its antioxidant property is related to it proton donating property [15, 33]. Therefore, as the activity of LCAT has previously been reported to be decreased in subjects with T2D [34], we have now shown that increased F&V intake augmented its activity within serum and particularly in HDL3 in this T2D population. This increase in LCAT’s activity may be related to the proton donating capacities of the additional F&V derived antioxidants, thus illustrating another antiatherogenic effect of increased F&V intake.
BMI, inflammation and the role of increased F&V intake
We must address the fact that BMI increased in the subjects randomised to the ≥6-portion intervention, in spite of the diets being designed and appearing isocalorific in both groups (p > 005). However, as both carbohydrate and total sugars significantly increased following the ≥6 portion intervention, and as these compounds are readily converted to fatty acids, this may have contributed to the increased BMI in this group, although a point to note is that these increases did not appear to influence the long term marker of glycaemic control, namely HbA1C, which was not different in the between group comparisons. Unfortunately, this increase in BMI may have negated a decrease in SAA, as there was only a trend for this inflammatory marker to decrease in HDL3 following the ≥6-portion intervention (p = 0.133). This was not consistent with previous non-reported findings by our group, where SAA did decrease following increased F&V intake, although in this study BMI remained stable. Therefore, the increase in BMI (0.3 Kg/m2) in the ≥6-portion group may have augmented SAA release, especially as SAA is released chronically from hypertrophic adipocytes [5]. Thus, the overall effect would be that any SAA-lowering effects afforded by the increased F&V intake would have been opposed by this additional adipose tissue. Overall, this finding raises a note of caution regarding the need to ensure that the composition of a F&V intervention should be closely monitored, to ensure that BMI remains stable. Nevertheless, this increase in BMI did not appear to impact on the enzymes examined in this current study.
Finally, we must consider the non-significant (p = 0.295) gender difference between the two groups at randomisation, where 20% of the subjects in the 1-portion intervention were female, while females made up 38.5% of the subjects in the ≥6-portion intervention. However, although females usually attach greater importance to healthy eating [35], all subjects appeared to adhere to the intervention group to which they were assigned, irrespective of gender. This was shown by the similar self-reported F&V intake and changes in carotenoids in both genders from each of the two groups (results not shown).
Conclusion
In subjects with T2D, increased F&V consumption resulted in an increase in carotenoid levels and the activities of PON-1 and LCAT, which was particularly apparent in HDL3, which we suggest would augment the antioxidant capabilities of this lipoprotein and would enhance its antiatherogenic potential. This study provides mechanistic support for the effectiveness of increased F&V intake, which may be a realistic lifestyle intervention to help lower the risk of T2D and its associated CVD.
Abbreviations
- AI:
-
(apo AI) Apolipoprotein
- BMI:
-
body mass index
- CVD:
-
Cardiovascular disease
- F&V:
-
Fruit and vegetable
- FSA:
-
Food Standards Agency
- HDL:
-
High-density lipoprotein
- HPLC:
-
High performance liquid chromatography
- hsCRP:
-
High-sensitivity C-reactive protein
- LCAT:
-
Lecithin cholesterol acyltransferase
- PON-1:
-
Paraoxonase-1
- RCT:
-
Reverse cholesterol transport
- T2D:
-
Type 2 diabetes.
References
Hamano M, Saito M, Eto M, Nishimatsu S, Suda H, Matsuda M, Matsuki M, Yamamoto S, Kaku K: Serum amyloid A, C-reactive protein and remnant-like lipoprotein particle cholesterol in type 2 diabetic patients with coronary heart disease. Ann Clin Biochem. 2004, 41 (2): 125-129. 10.1258/000456304322880005.
World Health Organisation: Cardiovascular Disease. 2012, http://www.who.int/mediacentre/factsheets/fs317/en/index.html.
Zhang Q, Zhang Y, Feng H, Guo R, Jin L, Wan R, Wang L, Chen C, Li S: High density lipoprotein (HDL) promotes glucose uptake in adipocytes and glycogen synthesis in muscle cells. PLoS One. 2011, 6 (8): e23556-10.1371/journal.pone.0023556.
Mastorikou M, Mackness B, Liu Y, Mackness M: Glycation of paraoxonase-1 inhibits its activity and impairs the ability of high-density lipoprotein to metabolize membrane lipid hydroperoxides. Diabet Med. 2008, 25 (9): 1049-1055. 10.1111/j.1464-5491.2008.02546.x.
Hatanaka E, Monteagudo PT, Marrocos MS, Campa A: Interaction between serum amyloid A and leukocytes - a possible role in the progression of vascular complications in diabetes. Immunol Lett. 2007, 108 (2): 160-166. 10.1016/j.imlet.2006.12.005.
Khovidhunkit W, Shigenaga JK, Moser AH, Feingold KR, Grunfeld C: Cholesterol efflux by acute-phase high density lipoprotein: role of lecithin: cholesterol acyltransferase. J Lipid Res. 2001, 42 (6): 967-975.
Mackness B, Mackness M: Anti-inflammatory properties of paraoxonase-1 in atherosclerosis. Adv Exp Med Biol. 2010, 660: 143-151. 10.1007/978-1-60761-350-3_13.
Villegas R, Shu XO, Gao YT, Yang G, Elasy T, Li H, Zheng W: Vegetable but not fruit consumption reduces the risk of type 2 diabetes in chinese women. J Nutr. 2008, 138 (3): 574-580.
Panagiotakos DB, Pitsavos C, Kokkinos P, Chrysohoou C, Vavuranakis M, Stefanadis C, Toutouzas P: Consumption of fruits and vegetables in relation to the risk of developing acute coronary syndromes; the CARDIO2000 case–control study. Nutr J. 2003, 8: 2-2.
Cooper AJ, Sharp SJ, Lentjes MA, Luben RN, Khaw KT, Wareham NJ, Forouhi NG: A prospective study of the association between quantity and variety of fruit and vegetable intake and incident type 2 diabetes. Diabetes Care. 2012, 35: 1293-1300. 10.2337/dc11-2388.
McEneny J, Wade L, Young IS, Masson L, Duthie G, McGinty A, McMaster C, Thies F: Lycopene intervention reduces inflammation and improves HDL functionality in moderately overweight middle-aged individuals. J Nutr Biochem. 2013, 24 (1): 163-168. 10.1016/j.jnutbio.2012.03.015.
Asgard R, Rytter E, Basu S, Abramsson-Zetterberg L, Moller L, Vessby B: High intake of fruit and vegetables is related to low oxidative stress and inflammation in a group of patients with type 2 diabetes. Scand J Food Nutr. 2007, 51 (4): 149-158. 10.1080/17482970701737285.
Blum S, Aviram M, Ben-Amotz A, Levy Y: Effect of a Mediterranean meal on postprandial carotenoids, paraoxonase activity and C-reactive protein levels. Ann Nutr Metab. 2006, 50 (1): 20-24. 10.1159/000089560.
National Health Service: 5 A Day Portion Sizes. 2011, http://www.nhs.uk/livewell/5aday/documents/downloads/5aday_portion_guide.pdf.
McPherson PA, Young IS, McKibben B, McEneny J: High density lipoprotein subfractions: isolation, composition, and their duplicitous role in oxidation. J Lipid Res. 2007, 48 (1): 86-95.
McEneny J, Loughrey CM, McNamee PT, Trimble ER, Young IS: Susceptibility of VLDL to oxidation in patients on regular haemodialysis. Atherosclerosis. 1997, 129 (2): 215-220. 10.1016/S0021-9150(96)06019-4.
Craft NE: Carotenoid reversed-phase high-performance liquid chromatography methods: reference compendium. Methods Enzymol. 1992, 213: 185-205.
Hasselwander O, McEneny J, McMaster D, Fogarty DG, Nicholls DP, Maxwell AP, Young IS: HDL composition and HDL antioxidant capacity in patients on regular haemodialysis. Atherosclerosis. 1999, 143 (1): 125-133. 10.1016/S0021-9150(98)00286-X.
Pearson TA, Mensah GA, Alexander RW, Anderson JL, 3rd Cannon RO, Criqui M, Fadl YY, Fortmann SP, Hong Y, Myers GL, Rifai N, Smith SC, Taubert K, Tracy RP, Vinicor F, Centres for Disease Control and Prevention; American Heart Association: Markers of inflammation and cardiovascular disease: application to clinical and public health practice: A statement for healthcare professionals from the Centres for Disease Control and Prevention and the American Heart Association. Circulation. 2003, 107: 499-511. 10.1161/01.CIR.0000052939.59093.45.
McEvoy CT, Temple N, Woodside JV: Vegetarian diets, low-meat diets and health: a review. Public Health Nutr. 2012, 15 (12): 2287-2294. 10.1017/S1368980012000936.
Di Mascio P, Kaiser S, Sies H: Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch Biochem Biophys. 1989, 274 (2): 532-538. 10.1016/0003-9861(89)90467-0.
Nielsen F, Mikkelsen BB, Nielsen JB, Anderson HR, Grandjean P: Plasma malondialdehyde as biomarker for oxidative stress: reference interval and effects of life-style factors. Clin Chem. 1997, 43 (7): 1209-1214.
Fenercioglu AK, Saler T, Genc E, Sabuncu H, Altuntas Y: The effects of polyphenol-containing antioxidants on oxidative stress and lipid peroxidation in Type 2 diabetes mellitus without complications. J Endocrinol Invest. 2010, 33 (2): 118-124.
Juanola-Falgarona M, Salas-Salvadó J, Estruch R, Portillo MP, Casas R, Miranda J, Martínez-González MA, Bulló M: Association between dietary phylloquinone intake and peripheral metabolic risk markers related to insulin resistance and diabetes in elderly subjects at high cardiovascular risk. Cardiovasc Diabetol. 2013, 12: 7-10.1186/1475-2840-12-7.
Fernemark H, Fernemark H, Jaredsson C, Bunjaku B, Rosenqvist U, Nystrom FH, Guldbrand H: A randomized cross-over trial of the postprandial effects of three different diets in patients with type 2 diabetes. PLoS One. 2013, 8 (11): e79324-10.1371/journal.pone.0079324.
Ortega-Azorín C, Sorlí JV, Asensio EM, Coltell O, Martínez-González MÁ, Salas-Salvadó J, Covas MI, Arós F, Lapetra J, Serra-Majem L, Gómez-Gracia E, Fiol M, Sáez-Tormo G, Pintó X, Muñoz MA, Ros E, Ordovás JM, Estruch R, Corella D: Associations of the FTO rs9939609 and the MC4R rs17782313 polymorphisms with type 2 diabetes are modulated by diet, being higher when adherence to the Mediterranean diet pattern is low. Cardiovasc Diabetol. 2012, 11: 137-10.1186/1475-2840-11-137.
Hockerstedt A, Tikkanen MJ, Jauhiainen M: LCAT facilitates transacylation of 17 beta-estradiol in the presence of HDL3 subfraction. J Lipid Res. 2002, 43 (3): 392-397.
Bergmeier C, Siekmeier R, Gross W: Distribution spectrum of paraoxonase activity in HDL fractions. Clin Chem. 2004, 50 (12): 2309-2315. 10.1373/clinchem.2004.034439.
Aviram M, Rosenblat M, Billecke S, Erogul J, Sorenson R, Bisgaier CL, Newton RS, La Du B: Human serum paraoxonase (PON 1) is inactivated by oxidized low density lipoprotein and preserved by antioxidants. Free Radic Biol Med. 1999, 26 (7–8): 892-904.
Calabresi L, Franceschini G: Lecithin:cholesterol acyltransferase, high-density lipoproteins, and atheroprotection in humans. Trends Cardiovasc Med. 2010, 20 (2): 50-53. 10.1016/j.tcm.2010.03.007.
Abbott CA, Mackness MI, Kumar S, Boulton AJ, Durrington PN: Serum paraoxonase activity, concentration, and phenotype distribution in diabetes mellitus and its relationship to serum lipids and lipoproteins. Arterioscler Thromb Vasc Biol. 1995, 15 (11): 1812-1818. 10.1161/01.ATV.15.11.1812.
Aviram M, Dornfeld L, Rosenblat M, Volkova N, Kaplan M, Coleman R, Hayek T, Presser D, Fuhrman B: Pomegranate juice consumption reduces oxidative stress, atherogenic modifications to LDL, and platelet aggregation: studies in humans and in atherosclerotic apolipoprotein E-deficient mice. Am J Clin Nutr. 2000, 71 (5): 1062-1076.
Vohl MC, Neville TA, Kumarathasan R, Braschi S, Sparks DL: A novel lecithin-cholesterol acyltransferase antioxidant activity prevents the formation of oxidized lipids during lipoprotein oxidation. Biochemistry. 1999, 38 (19): 5976-5981. 10.1021/bi982258w.
Nakhjavani M, Asgharani F, Khalilzadeh O, Esteghamati A, Ghaneei A, Morteza A, Anvari M: Oxidized low-density lipoprotein is negatively correlated with lecithin-cholesterol acyltransferase activity in type 2 diabetes mellitus. Am J Med Sci. 2011, 341 (2): 92-95. 10.1097/MAJ.0b013e3181f3ce10.
Westenhoefer J: Age and gender dependent profile of food choice. Forum Nutr. 2005, 57: 44-51.
Acknowledgments
Foremost, we would like to thank Dr Ciara Mulligan (formally McLaughlin) who was responsible for the recruitment and running of the primary study, which provided the samples for these secondary analyses. We would also like to thank the staff at the Regional Centre for Endocrinology and Diabetes at the Royal Victoria Hospital, Belfast, UK, and the participants who took part in this study. We would also like to thank Mrs Kathy Pogue for assisting with the HPLC analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interest.
Authors’ contributions
JAD undertook the laboratory analyses for this study and drafted the manuscript. CM (formally McLaughlin) was responsible for the recruitment and running of the primary study and commented on the current manuscript. DMcC supervised CM in the primary analyses and also commented on the current manuscript. JW was involved in the primary analyses and also commented on the current manuscript. IY supervised CM in the primary analyses and also commented on the current manuscript. JMcE was responsible for the study concept, supervised JAD and also had the final say in manuscript preparation and submission. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Daniels, JA., Mulligan, C., McCance, D. et al. A randomised controlled trial of increasing fruit and vegetable intake and how this influences the carotenoid concentration and activities of PON-1 and LCAT in HDL from subjects with type 2 diabetes. Cardiovasc Diabetol 13, 16 (2014). https://doi.org/10.1186/1475-2840-13-16
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2840-13-16