
Abstract
Background
The effect of rainfall patterns on soil surface CO2 efflux, soil moisture, soil temperature and plant growth was investigated in a grassland ecosystem of northern Ontario, Canada, where climatic change is predicted to introduce new precipitation regimes. Rain shelters were established in a fallow field consisting mainly of Trifolium hybridum L., Trifolium pratense L., and Phleum pratense L. Daytime ambient air temperatures within the shelters increased by an average of 1.9°C similar to predicted future increases in air temperatures for this region. To simulate six precipitation regimes which cover the maximum range to be expected under climate change, a portable irrigation system was designed to modify the frequency of monthly rainfall events with a constant delivery rate of water, while maintaining contemporary average precipitation volumes. Controls consisted of blocks irrigated with frequencies and total monthly precipitation consistent with the 25 year average rainfall for this location.
Results
Seasonal soil moisture correlated with soil surface CO2 efflux (R = 0.756, P < 0.001) and above ground plant biomass (R = 0.447, P = 0.029). By reducing irrigation frequency, soil surface CO2 efflux decreased by 80%, P < 0.001, while soil moisture content decreased by 42%, P < 0.001.
Conclusions
Manipulating the number of precipitation events and inter-rainfall intervals, while maintaining monthly rainfall averages impacted CO2 efflux and plant growth. Even with monthly rainfall averages that are similar to contemporary monthly precipitation averages, decreasing the number of monthly rainfall events reduced soil surface CO2 efflux and plant growth through soil moisture deficits. Although many have speculated that climate change will increase ecosystem productivity, our results show that a reduction in the number of monthly rainfall events while maintaining monthly averages will limit carbon dynamics.
Similar content being viewed by others


Background
One of the greatest contemporary challenges in terrestrial ecology is to determine the future impact of climate change on the world's ecosystems. Because climate has a fundamental influence on the distribution and productivity of ecosystems, impacts from anthropogenic greenhouse gas emissions will have far-reaching social, economic and ecological consequences. Thus far, numerous general circulation models project a global mean temperature increase of 0.8–3.5°C by 2100 AD [1] and predicted changes in regional climate have been used as a basis to speculate about the effect of climate change on vegetation. In practice, the effect of climate change at the local level should vary due to the interaction among climate, topography and vegetation. Hence, ecologists must determine the influence of climate change on specific ecosystems in order to predict its impact at the local level.
However, the ecological dynamics that governed the establishment, succession, and maintenance of contemporary ecosystems may be altered dramatically under climate change: 1) possibly climate change will introduce new climatic extremes that may completely override contemporary ecological precepts regarding species distribution [2]; 2) current ecosystems are the result of thousands of years of interaction between climate and vegetation whereas new ecosystem types should rapidly develop under climate change [3]; and 3) human influences at the local and regional levels may restrict species migration, succession and dispersion across landscapes [4].
The objective of this investigation was to measure the impact of the range of precipitation regimes potentially associated with climate change on soil surface CO2 efflux (SSCE), plant growth, soil moisture and soil temperature in northern Ontario, Canada. We hypothesized that rainfall patterns will play a critical influence in future ecosystems. As a general rule heavy rainfalls may become more frequent, with light rainfalls occurring less frequently [5]. As well, incidence of droughts and growth-limiting water deficits may increase from the current five percent of the time to more than 50 percent by the 2050's [6]. The length of the growing season is predicted to increase by 41–61 days, and mean air temperature to rise by 1.5 – 1.9°C [7]. Smit [8] speculates that even if monthly rainfall volumes increase, precipitation effectiveness could be greatly reduced through runoff, evapotranspiration or a decrease in the number of rainfall events. Presumably, increased evapotranspiration should render the northern Ontario climate drier than present, to the extent that it should be similar to the contemporary climate of southwestern Ontario, located 500 km south [7]. We attempted to capture the range of extreme rainfall frequencies by manipulating inter-rainfall intervals and keeping monthly averages constant.
Results
As the irrigation treatments became more intense and less frequent, there was a decrease of 42% in soil moisture content at P < 0.001 (Table 1). The most frequent (6-litre treatment every 2 days) led to the greatest soil moisture content of 25.59%, where the least frequent (84-litre treatment every 28 days) produced a soil moisture content averaging 18.07%.
Similarly, as the irrigation treatments became more intense and less frequent, SSCE decreased by 80%, P < 0.001 (Table 1). The greatest respiration rates (5.26 μmol m-2 s-1) were associated with the most frequent low volume irrigation treatment, whereas the smallest respiration rates (2.92 μmol m-2 s-1) were associated with the least frequent high volume irrigation treatment. SSCE correlated with both soil moisture at R = 0.756, P < 0.001, and above ground plant biomass at R = 0.379 and P = 0.067 (Table 2).
Soil moisture and plant biomass correlated at R = 0.447, P = 0.029 (Table 2). There was a decrease in above ground plant biomass of 45% as irrigation frequency decreased but the treatment effect was not significant at P = 0.419 (Table 1).
Discussion
Our results provide valuable insight into the effect of precipitation regimes on ecosystems. Current models for northern Ontario predict that increases in temperature, precipitation and longer growing seasons associated with climate change should lead to increased net productivity of agricultural crops where soils permit [8]. However, our findings show that increasing inter-rainfall intervals, while maintaining monthly precipitation volumes, impacts ecosystem productivity. Longer than average inter-rainfall intervals led to a negative correlation with SSCE, plant growth and soil moisture content in our experimental system (Table 1). Interestingly, because our investigation was conducted on a luvisol, which has greater water holding capacity than other soil types in northern Ontario [9], we speculate that the effect of the longer than average inter-rainfall intervals used in this investigation might be equivalent or greater on most other soil types.
In this investigation SSCE and soil moisture decreased significantly (P < 0.001, Table 1) as the inter-rainfall intervals was increased while maintaining monthly precipitation averages. SSCE is an important component of the carbon budget of agricultural or grassland ecosystems [10] and is made of varying contributions from root respiration [11] and microbial respiration. However, soil CO2 release can be limited either by extremes of saturation or water deficits [12, 13]. We were unable to incorporate into this investigation the effect of elevated atmospheric CO2 on SSCE. However, Luo [14] showed that SSCE doubles under high atmospheric CO2 levels such as those predicted under climate change. Therefore, we speculate that our observed results on the impact of increased inter-rainfall intervals on SSCE should be even more pronounced under elevated atmospheric CO2.
Contrary to the hypothesis that elevated temperature and atmospheric CO2 should increase plant productivity and water use efficiency under climate change [15, 16], our observations and those of other investigators [17, 18], suggest that inter-rainfall intervals are by themselves critical in controlling ecosystem productivity through soil moisture deficits [19]. Indeed, our results show that the average soil moisture is significantly lower (P < 0.001) in plots with greater inter-rainfall intervals compared with plots having shorter intervals, even though total monthly rainfall remained constant (Table 1). One implication of our findings to the forestry, cattle and dairy industries is that climate change will demand a greater land base to maintain current harvest levels. To maintain the current land base, irrigation measures will have to be implemented or cattle stocking rates and timing of grazing will have to be altered. O'Lear and Blair [20] support our findings by speculating that future weather patterns will impact grasslands since many of these ecosystems are influenced by water availability.
Although there were no direct, significant impacts on plant growth by the irrigation treatments in this experiment, there was a correlation between soil moisture and plant biomass at R = 0.447, P = 0.029, (Table 2). As irrigation frequency declined, we noted that soil moisture declined by 42%, and above ground plant biomass also declined by 45%, (Table 1). A reduction of 80% in SSCE indicates a possible link between these two variables and suggests that below ground biomass production was also affected since the direct contributions of roots to SSCE is of considerable significance in most agricultural ecosystems [21].
Our observations lend support in predicting the impacts of altered rainfall patterns on ecosystem processes, both for carbon models, and for plant growth models, as the total efflux of CO2 from the soil surface is derived from the respiration of plant roots and microbial respiration. According to Amthor [21] the fractions of total SSCE resulting from root respiration and microbial respiration can vary considerably from site to site. It was not possible for us to partition these components of SSCE, however, both are strongly influenced by available soil moisture [13]. We also observed a positive correlation of SSCE and above ground plant biomass (P = 0.067). Therefore, the decreased soil moisture in the least frequent irrigation regime has the potential to alter not only above ground plant biomass but SSCE through reduced root production [12, 22], and microbial activity [23].
Conclusions
Reductions in inter-rainfall frequency, while maintaining current monthly precipitation averages, decrease SSCE, plant biomass production and soil moisture. Our findings are supported by Fay et al. [24] who suggests that the intervals between rainfall events may be the main variable affecting SSCE and aboveground net primary productivity under climate change. The relationships between SSCE and these variables are significant (Table 2), indicating that SSCE can serve as a reliable indicator of ecosystem disturbance [25] through the measurement of soil biological activity. Indeed, Dulohery et al. [26] speculates that CO2 evolution could be the most vigorous and stable biological indicator of soil condition, encompassing most metabolic activities in the soil. However, our work is limited to one ecosystem type and further investigation is needed worldwide to ascertain the validity of SSCE as an indicator of productivity in other ecosystem types.
Methods
Study site
The study site is located near Echo Bay, Ontario, Canada, (46°25 'N, 84°02 ' W) approximately 28 kilometres southeast of Sault Ste. Marie, Ontario (map 41 K/8 & 41 K/7 278444). The site is in the Algoma Section (L.10) of the Great Lakes-St. Lawrence Forest Region [27]. The topography of the area is undulating, with an average altitude of 205 m above sea level and a gentle slope (<1%) facing towards NNE. The soil is a gleyed gray luvisol over a thick (>10 m) clay-loam deposit [28].
The surface vegetation consists predominately of Trifolium hybridum L.,Trifolium pratense L., and Phleum pratense L., with a relative abundance of 1/1/1, w/w/w. This site was selected for its uniformity in vegetation distribution with a total area of 10 ha allowing for adequate spacing of plots from potential impacts of shade and competition by surrounding forests. Primeval forests were cleared for agriculture approximately 100 years ago and used for forage growth until the 1980's when a change in land ownership occurred. Since then the land has been sitting fallow. The surrounding, remaining forest over storey consists of approximately 40-year-old Populus tremuloides Michx. with Betula papyrifera Marsh., Pinus strobus L., Picea glauca (Moench) Voss, Acer rubrum L and Thuja occidentalis L.
The climate is temperate continental, with an average growing season length of 200 days from mid-April through late-October, and mean growing season precipitation of 500 mm [29]. Total annual precipitation at the study site during the period between 1967 and 1997 was 906 mm, of which approximately 30% was snow. Mean annual temperature is 4.4°C, with a monthly mean of -10.4°C for January and 14.3°C for July [30].
Rain shelters
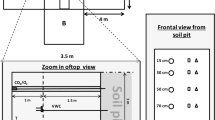
To test the effect of altered precipitation we established the experiment within five rain shelters (7.3 m × 7.3 m) in the spring of 1998. The shelters consisted of wooden frames with a slanted roof (4:12 pitch; 1 m above ground at the lowest point of the roof), mounted on nine support posts with an average roof height above vegetation of 1.6 m. The roof was covered by transparent vapour barrier (6 mil: Duchesne Enterprises, Joliette, Que.), and held in place by wooden slats. Air was allowed to circulate freely within the structure as the side walls were covered only by 3 cm2 wire mesh to exclude wildlife. This height of the rain shelter was selected through trials and errors to increase inside noon air temperatures 10 cm above the soil surface by 1.9°C ± 0.9°C, which coincided with the predicted increase of the air temperature in this region under climate change [7].
Irrigation regime
The experiment was established as a randomized complete block design and within each of the five rain shelters (blocks), sixteen 1 m × 1 m plots were staked out with 20 cm buffer zones between each plot, and 1.25 m buffers around the outside perimeter to avoid effects from natural rainfall. Three plots within each shelter were randomly selected for each of the five irrigation treatments (leaving the sixteenth plot unused), with one plot dedicated to SSCE measurements and soil moisture core extractions, one plot for above ground biomass harvests, and one for a seed germination study not included in this paper. The shelters were covered on June 8 – 9, 1998, and irrigation commenced on June 11, 1998 and continued through Sept. 8, 1998. To simulate different precipitation scenarios, we modified the number of monthly rainfall events, while maintaining contemporary average monthly precipitation in a replicated experimental design. For this, the 30-year average June precipitation for the Algoma District (84 mm: Environment Canada, Luc Pouliot, Pers. Com.) was delivered artificially in a 28 day cycle as 1, 2, 4, 8, and 14 rainfall events in the form of 6, 12, 24, 42 and 84 L m-2, respectively. Historical weather data indicates few variations in growing season precipitation patterns therefore the June precipitation regime was applied for the duration of the experiment. Irrigation water was obtained from a domestic well and delivered by hand with a slosh nozzle at a constant flow rate of 2 L min-1 that minimized lateral movement of water across the soil surface. Controls consisted of the 14 rainfall events every 28 days based on the 25 year measurable precipitation frequency of one precipitation event every 2.4 days for this area [30]. Hence treatments are designated: Control (6 litres m-2 of water applied every 2 days), 12 litres m-2 every 4 days, 24 litres m-2 every 8 days, 42 litres m-2 every 14 days, and 84 litres m-2 every 28 days. Volumes of irrigation water were calibrated at the beginning of the experiment and checked weekly throughout its duration.
Soil surface CO2 efflux
SSCE was used as an indicator of ecological processes as it integrates microbial decomposition and root respiration [31]. SSCE was measured using a LI-6200, dynamic, flow through, portable, infrared gas analyzer (LI-COR, Lincoln, NE, USA) from each treatment at 4-day intervals from July 22 through Sept. 8, 1998. Measurements were taken at approximately the same time each day (0900 through 1400 hrs) to minimize changes in flux rates caused by the diurnal cycle [32].
To avoid soil disturbance and allow repeated measurements at the same locations throughout the experiment, 25 permanent soil collars 10 cm in diameter and 4.4 cm high, constructed out of poly vinyl chloride pipe were installed 1 cm into the soil surface 48 hours prior to the first measurement. The collars were randomly located in each of the five irrigation treatments, and replicated through each rain shelter. To standardize the LI-6200 internal system volume, the collars were kept free of herbaceous vegetation by clipping aboveground biomass bi-weekly throughout the experiment. Collars were not inserted deeper than 1 cm to facilitate root growth into the soil directly under the collar to minimize bias caused by lowering the contribution of roots to total SSCE and minimize any impacts caused by clipping.
Prior to each measurement, the ambient CO2 concentration was measured by diverting an air sample through a tube located adjacent to each soil collar. A portable umbrella was positioned to shade the collar and soil chamber assembly to prevent overheating and gas expansion within the chamber. The chamber was then placed on the soil collar and the CO2 concentration in the chamber was lowered below ambient CO2 levels by scrubbing with soda lime for a few seconds. The scrubber was then turned off and the atmosphere within the chamber allowed to equilibrate prior to logging. When SSCE causes the CO2 within the chamber to rise, the logging interval begins, with the CO2 logging range spanning concentrations below, near, and above the ambient levels. The spanning of ambient concentrations external to the chamber during logging was performed to reduce the incidence of leaks between the chamber and the outside atmosphere and represents the best estimate of flux with the minimum leakage error [33]. Each measurement took approximately 2 to 3 minutes to complete depending on the SSCE rate. Between measurements, the closed loop within the infrared gas analyzer (IRGA) was opened via a toggle switch, with the chamber fan running at maximum speed to restore the entire IRGA and internal instrument plumbing to ambient CO2 concentrations. In order to avoid passive soil degassing caused by irrigation, CO2 measurements commenced at least two hours after watering as instructed by LI-COR [34].
SSCE was expressed in μmol m-2 s-1 and based on the difference of CO2 concentrations entering and leaving the chamber. The respiration rate was computed using the equations supplied for the LI-6200 portable photosynthesis system [34].
Soil temperature, moisture, and plant biomass
At each SSCE measurement, soil temperature was measured using a soil temperature thermocouple probe (Omega Engineering Inc., Stamford, CT) adjacent to the PVC collar to a depth of 10 cm below the soil surface. This temperature data was interfaced with the LI-6200 and the data recorded using the LI-6200 software. Percent soil water content was measured during each CO2 reading for a 0- to 10-cm depth by extracting a 6 cm diameter soil core within one-half meter of the permanent sample collar. To minimize the impact on soil water balance, soil core holes were backfilled with similar materials and marked with colored toothpicks to avoid other samples being taken from the same location. The cores were placed in plastic bags and frozen until processed. The cores were weighed before drying then dried in an oven for three days at 70°C, and then re-weighed again.
Above ground plant biomass was harvested from one, 1 × 1 m plot per treatment per shelter for each of the five rain shelters, in June and again in August to simulate total biomass harvest under an average agricultural season. For this, all above ground vegetation within the plot was cut, placed in plastic bags, dried at 70°C for three days and weighed. Weights from both harvests were combined for a seasonal total and expressed in terms of g m-2.
Statistical analyses
SSCE, soil moisture and soil temperature data showed non significant differences within treatments over the course of the growing season at p < 0.05. Therefore the repeated measures for each treatment were pooled. The effect of treatments on SSCE, soil moisture, soil temperature and above ground plant biomass were analyzed using analysis of variance (ANOVA) to test for significant differences using Sigma Stat [35]. When ANOVA results indicated a significant treatment effect, Tukey's HSD test was performed on each reduced data set to determine which levels of the treatments differed significantly (α = 0.05). Strength of associations were tested using Pearson Product moment correlation and regression analyses were conducted using SYSTAT 8.0 [36].
References
Stocks BJ, Fosberg MA, Wotton BM, Lynham TJ, Ryan KC: Climate change and forest fire activity in North American boreal forests. Fire, climate change, and carbon cycling in the boreal forest, Ecological Studies, 138, Springer-Verlag, New York. 2000
Environment Canada: The Canada Country Study: climate impacts and adaptation. Responding to Global Climate Change in the Arctic. Ottawa. 1997, [http://www.ec.gc.ca/climate/ccs/arc_summ.htm]
Natural Resources Canada: Climate change and forests: context for the Canadian Forest Service's science program. Cat. No. Fo42-289/1999, Natural Resources Canada, Canadian Forest Service, Ottawa. 1999, 7-
Parker WC, Colombo SJ, Cherry ML, Greifenhagen S, Papadopol CS, Scarr T: Forest management responses to climate change. In: The impacts of climate change on Ontario's forests. Ministry of Natural Resources, Ontario Forest Research Institute, Sault Ste Marie, Ontario. Forest Research Information Paper, ISSN 1319-9118. Edited by: Colombo SJ, Buse LJ. 1998, 143: 34-43.
Francis D, Hengeveld H: Extreme weather and climate change. Climate Change Digest, Atmospheric Environment Service, Minister of Supply and Services Canada, Climate and Water Products Division, Downsview, On. 1998, , Catalogue No. En57-27/1998-01E
MacCracken MC, Budyko MI, Hecht AD, Izrael YA, eds: Prospects for future climate, A special US/USSR report on climate and climate change. 1990, Lewis Publishers, Inc., Chelsea, Michigan, 217:
Stewart R: Possible effects of climatic change on estimated crop yields in Canada: A review. Proceedings of an international symposium/workshop on climate change: Implications for water and ecological resources, Department of Geography Publication Series No. 11, University of Waterloo, Ontario. 1990, 275-283.
Smit B: Implications of climatic change for agriculture in Ontario. Climate Change Digest, Atmospheric Environment Service, Canadian Program Office, Canadian Climate Centre, Downsview, On. 1987, , Catalogue No. EN57-27/1987-01
Armson KA: Forest soils. University of Toronto Press, Toronto. 1977
Rochette P, Ellert B, Bregorick EG, Desjardins RL, Patty E, Lessard R, Johnson BG: Description of a dynamic closed chamber for measuring soil respiration and its comparison with other techniques. Can J Soil Sci. 1997, 77: 195-203.
Norman JM, Garcia R, Verma SB: Soil surface CO2 fluxes and the carbon budget of a grassland. J Geophys Res. 1992, 97 (D17): 18845-18853.
Kucera CL, Kirkham DR: Soil respiration studies in tallgrass prairie in Missouri. Ecology. 1971, 52 (5): 912-915.
Howard DM, Howard PJA: Relationships between CO2 evolution, moisture content and temperature for a range of soil types. Soil Biol Biochem. 1993, 25 (11): 1537-1546. 10.1016/0038-0717(93)90008-Y.
Luo Y, Jackson RB, Field CB, Mooney HA: Elevated CO2 increases belowground respiration in California grasslands. Oecologia. 1996, 108: 130-137.
Jones MB, Jongen M, Doyle T: Effects of elevated carbon dioxide concentrations on agricultural grassland production. Agri and For Meteor. 1996, 79 (4): 243-252. 10.1016/0168-1923(95)02289-9.
Hunt HW, Trlica MJ, Redente EF, Moore JC, Detling JK, Kittel TGF, Walter DE, Fowler MC, Klein DA, Elliott ET: Simulation model for the effects of climate change on temperate grassland ecosystems. Ecol Model. 1991, 53 (3–4): 205-246.
Sala PE, Parton WJ, Joyce LA, Lauenroth WK: Primary production of the central grassland region of the United States. Ecology. 1988, 69 (1): 40-45.
Redmann RE: Soil respiration in a mixed grassland ecosystem. Can J Soil Sci. 1978, 58: 119-124.
Newton PCD, Clark H, Bell CC, Glasgow EM: Interaction of soil moisture and elevated CO2 on the above-ground growth rate, root length density and gas exchange of turves from temperate pasture. J Exp Bot. 1996, 47 (299): 771-779.
O'Lear HA, Blair JM: Responses of soil microarthropods to changes in soil water availability in tallgrass prairie. Biol Fert of Soils. 1999, 29 (2): 207-217. 10.1007/s003740050546.
Amthor JS: Respiration and crop productivity. Springer-Verlag, New York. 1989
Mogensen VO: Field measurements of dark respiration rates of roots and aerial parts in Italian ryegrass and barley. J Appl Ecol. 1977, 14: 243-252.
Pietikäinen J, Vaijärvi E, Ilvesniemi H, Fritze H, Westman CJ: Carbon storage of microbes and roots and the flux of CO2 across a moisture gradient. Can J For Res. 1999, 29: 1197-1203. 10.1139/cjfr-29-8-1197.
Fay PA, Carlisle JD, Knapp AK, Blair JM, Scott SL: Altering rainfall timing and quantity in a mesic grassland ecosystem: Design and performance of rainfall manipulation shelters. Ecosystems. 2000, 3 (3): 308-319. 10.1007/s100210000028.
Buyanovsky GA, Wagner GH, Gantzer CJ: Soil respiration in a winter wheat ecosystem. Soil Sci Soc Am J. 1986, 50: 338-344.
Dulohery CJ, Morris LA, Lowrance R: Assessing forest soil disturbance through biogenic gas fluxes. Soil Sci Soc Am J. 1996, 60: 291-298.
Rowe JS: Forest regions of Canada. Can For Serv, Ottawa. Publication Number 1300. 1972
Agriculture Canada: Soil survey report 50. Bruce Mines 41 J/5, Cartography Section, Land Resource Research Institute, Research Branch Agriculture Canada, Ottawa. 1983
Mackey BG, McKenney DW, Yang Y-Q, McMahon JP, Hutchinson MF: Site regions revisited: a climatic analysis of Hill's site regions for the province of Ontario using a parametric method. Can J For Res. 1996, 26: 333-354.
Environment Canada: Annual Meteorological Summary at Sault Ste. Marie "A", Ontario. 1997
Fernandez IJ, Son Y, Kraske CR, Rustad LE, David MB: Soil carbon dioxide characteristics under different forest types and after harvest. Soil Sci Soc Am J. 1993, 57: 1115-1121.
Davidson EA, Belk E, Boone RD: Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology. 1998, 4: 217-227. 10.1046/j.1365-2486.1998.00128.x.
LI-COR: 6000-09 Soil respiration chamber instruction manual, 1993. LI-COR, inc. 4421, Superior St., Lincoln, NE. 68504, USA. Publication No. 9311-69
LI-COR: LI-6200 technical reference, 1990. LI-COR, Box. 4425, /4421, Superior St., Lincoln, NE. 68504, USA
Jandel Corporation: Sigma stat users manual. Version 2.0. 1995
SPSS Inc.: SYSTAT 8.0 statistics users manual. Version 8.0, SPSS Inc., 233 South Wacher Dr., Chicago, IL. 60606-6307. 1998
Acknowledgments
Thanks are addressed to Y. Hurtubise and D. Jacobs for their excellent technical support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
Laporte MF carried out data collection, synthesis and statistical analysis, participated in the design and coordination of the study, and drafted the manuscript. Duchesne LC conceived the study, participated in its design and coordination, and participated in the drafting of the manuscript as senior author. Wetzel S participated in the design and implementation of the study, reviewed and modified the manuscript. All authors, Laporte, Duchesne and Wetzel have read and approve of the final manuscript.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Laporte, M.F., Duchesne, L. & Wetzel, S. Effect of rainfall patterns on soil surface CO2 efflux, soil moisture, soil temperature and plant growth in a grassland ecosystem of northern Ontario, Canada: implications for climate change. BMC Ecol 2, 10 (2002). https://doi.org/10.1186/1472-6785-2-10
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1472-6785-2-10