
Abstract
Background
The cattle ticks, Boophilus spp., affect cattle production in tropical and subtropical regions of the world. Tick vaccines constitute a cost-effective and environmentally friendly alternative to tick control. The recombinant B. microplus Bm86 protective antigen has been shown to protect cattle against tick infestations. Recently, the gene coding for B. annulatus Bm86 ortholog, Ba86, was cloned and the recombinant protein was secreted and purified from the yeast Pichia pastoris.
Results
Recombinant Ba86 (Israel strain) was used to immunize cattle to test its efficacy for the control of B. annulatus (Mercedes, Texas, USA strain) and B. microplus (Susceptible, Mexico strain) infestations. Bm86 (Gavac and Mozambique strain) and adjuvant/saline were used as positive and negative controls, respectively. Vaccination with Ba86 reduced tick infestations (71% and 40%), weight (8% and 15%), oviposition (22% and 5%) and egg fertility (25% and 50%) for B. annulatus and B. microplus, respectively. The efficacy of both Ba86 and Bm86 was higher for B. annulatus than for B. microplus. The efficacy of Ba86 was higher for B. annulatus (83.0%) than for B. microplus (71.5%). The efficacy of Bm86 (Gavac; 85.2%) but not Bm86 (Mozambique strain; 70.4%) was higher than that of Ba86 (71.5%) on B. microplus. However, the efficacy of Bm86 (both Gavac and Mozambique strain; 99.6%) was higher than that of Ba86 (83.0%) on B. annulatus.
Conclusion
These experiments showed the efficacy of recombinant Ba86 for the control of B. annulatus and B. microplus infestations in cattle and suggested that physiological differences between B. microplus and B. annulatus and those encoded in the sequence of Bm86 orthologs may be responsible for the differences in susceptibility of these tick species to Bm86 vaccines.
Similar content being viewed by others
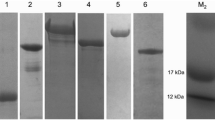

Background
Boophilus spp. (recently considered a synonym of Rhipicephalus (Boophilus) spp.) ticks are distributed in tropical and subtropical regions of the world with range expansion for some species due to changes in climatic conditions [1–3]. Infestations with the cattle tick, Boophilus microplus, economically impact cattle production by reducing weight gain and milk production, and by transmitting pathogens that cause babesiosis (Babesia bovis and B. bigemina) and anaplasmosis (Anaplasma marginale) [4]. B. annulatus is present in regions of Asia, Latin America and Africa [2] where it may also affect cattle production and vector pathogens.
Acaricide application constitutes a major component of integrated tick control strategies [5]. However, use of acaricides has had limited efficacy in reducing tick infestations and is often accompanied by serious drawbacks, including the selection of acaricide-resistant ticks, environmental contamination and contamination of milk and meat products with drug residues [5]. All of these issues reinforce the need for alternative approaches to control tick infestations such as the use of hosts with natural resistance to ticks, pheromone-impregnated decoys for attracting and killing ticks, biological control agents and vaccines [6–8].
In the early 1990s, vaccines were developed that induced immunological protection of vertebrate hosts against tick infestations. These vaccines contained the recombinant B. microplus Bm86 gut antigen [8–12]. Two vaccines using recombinant Bm86 were subsequently registered in Latin American countries (Gavac) and Australia (TickGARD) during 1993–1997 [13]. These vaccines reduce the number of engorging female ticks, their weight and reproductive capacity. Thus the greatest vaccine effect was the reduction of larval infestations in subsequent generations. Vaccine controlled field trials in combination with acaricide treatments demonstrated that an integrated approach resulted in control of tick infestations while reducing the use of acaricides [12–14]. These trials demonstrated that control of ticks by vaccination has the advantages of being cost-effective, reducing environmental contamination and preventing the selection of drug resistant ticks that result from repeated acaricide application. In addition, these vaccines may also prevent or reduce transmission of pathogens by reducing tick populations and/or affecting tick vectorial capacity [13–15].
Controlled immunization trials have shown that B. microplus Bm86-containing vaccines also protect against related tick species, B. annulatus and B. decoloratus [16–18]. However, B. microplus strain-to-strain variations in the susceptibility to Bm86 vaccination have been reported and the efficacy of the Bm86 vaccine is higher against B. annulatus than against B. microplus strains [6, 16–18]. These results suggested that Bm86 sequence and/or tick physiological differences may influence the efficacy of the vaccine in Boophilus spp. [18–22].
Recently, the gene coding for B. annulatus Bm86 ortholog, Ba86, was cloned from an Israeli tick strain and the recombinant protein was secreted and purified from Pichia pastoris [23]. The Bm86 and Ba86 proteins showed over 90% similarity and immune cross-reactivity [23]. However, only cattle vaccination and tick infestation experiments could evaluate the efficacy of Ba86 against B. annulatus and B. microplus infestations and address the question of whether differences in the susceptibility to Bm86 vaccines between these tick species are due to sequence polymorphisms, physiological characteristics of the ticks or both.
In the experiments reported herein cattle were vaccinated with the recombinant Bm86 and Ba86 antigens and infested with B. annulatus and B. microplus to (i) evaluate the efficacy of recombinant Ba86 for the control of B. annulatus and B. microplus infestations and (ii) to provide evidence of whether sequence polymorphisms, tick physiological differences or both may account for differences in the efficacy of Bm86 vaccines against Boophilus spp. infestations.
Results and Discussion
The vaccination with recombinant Ba86 and Bm86 protected cattle against B. annulatus and B. microplusinfestations
This is the first report on the protective capacity of recombinant Ba86 for the control of cattle tick infestations. To evaluate the protective capacity of Ba86 against B. microplus and B. annulatus infestations, cattle were vaccinated with the recombinant protein and compared to cattle vaccinated with two Bm86 preparations and adjuvant/saline control. The vaccinated animals but not the controls developed antibodies against recombinant proteins (Fig. 1). The antibody titers were similar for all groups when measured against Ba86 and Bm86 antigens, thus reflecting the presence of common antigenic epitopes between both proteins (ref. [23] and Fig. 2). The antibody titers in cattle vaccinated with Ba86 and Bm86 (Mozambique strain) were similar and higher than those in animals vaccinated with Bm86 (Gavac) (Fig. 1). As in previous experiments [11, 20], except for animals vaccinated with Bm86 (Gavac), antibody titers increased after successive vaccinations and reached a peak two weeks after the third vaccination shot. The differences in the antibody response elicited by Bm86 (Gavac) and Bm86 (Mozambique strain) antigens could be attributed to differences in vaccine preparations (the Bm86 in Gavac was expressed in P. pastoris as membrane-bound while the Bm86 of the Mozambique strain was secreted in P. pastoris; ref. 23] and/or to other unknown factors such as cattle physiological status that have been demonstrated to affect cattle antibody response to Bm86 [14]. Additionally, as discussed bellow, Bm86 polymorphisms between Gavac and Mozambique strain antigens may affect antigen processing and immune response after vaccination [19].

Antibody response in vaccinated cattle. Bovine serum antibody titers to recombinant Ba86 (Israeli strain) and Bm86 (Mozambique strain) antigens were determined by ELISA in cattle vaccinated with Bm86 (Gavac, Cuban Camcord strain), Bm86 (Mozambique strain), Ba86 (Israeli strain) and adjuvant/saline control. Antibody titers in immunized cattle were expressed as the OD450 nm value for the highest serum dilution (1:1000) and compared between vaccinated and control cattle using an ANOVA test (*P < 0.05). The time of vaccination shots (arrows) and tick infestation are indicated.

Sequence comparison of recombinant Bm86 and Bm86 antigens. The protein sequences of the Bm86 (Cuban Camcord in Gavac), Bm86 (Mozambique), Bm86 (Susceptible, Mexico), Ba86 (Israeli strain) and Ba86 (Mission, TX) strains were aligned and the antigenic peptides (≥ 7 residues) predicted using the method of Kolaskar and Tongaonkar [29], with a reported accuracy of about 75% http://immunax.dfci.harvard.edu/Tools/antigenic.pl. The predicted antigenic peptides are underlined. The antigenic peptide present in Bm86 but absent in Ba86 (Israel strain) is shown in underlined red letters. The antigenic peptides absent in the Bm86 (Mozambique strain) sequence are shown in underlined blue letters.
The vaccination with recombinant Ba86 protected cattle against B. microplus and B. annulatus infestations (Tables 1 and 2). As in previous experiments with recombinant Bm86 preparations, tick vaccines decreased the number and weight of adult female ticks, oviposition and egg fertility [9, 11, 12, 14, 16, 18, 20–24]. The efficacy of both Ba86 and Bm86 vaccines was higher for B. annulatus than for B. microplus (Tables 1 and 2). The efficacy of Ba86 was higher for B. annulatus (83.0%) than for B. microplus (71.5%) while the efficacy of Bm86 (Gavac; 85.2%) but not Bm86 (Mozambique strain; 70.4%) was higher than that of Ba86 (71.5%) on B. microplus. However, the efficacy of Bm86 vaccines was higher than that of Ba86 for B. annulatus and similar to previous reports showing close to 100% efficacy of Bm86 for the control of B. annulatus infestations [16, 18]. The efficacy of Bm86 vaccine preparations against B. microplus was within the range reported in previous experiments with other tick strains [12, 17–19, 24].
Polymorphisms in Bm86 orhtologs and physiological differences between B. annulatus and B. microplusmay account for differences in the efficacy of Bm86 vaccines
The fact that both Ba86 and Bm86 vaccines had a higher efficacy for B. annulatus than for B. microplus suggested a tick species-specific effect that resulted in higher susceptibility of B. annulatus to vaccination. This effect may be related to tick physiological processes such as feeding and digestion. For example, a higher amount of blood ingestion or a lesser protease activity in the gut of B. annulatus would result in an increase in the number of antibody-antigen interactions and vaccine efficacy. A direct correlation between antibody titers and vaccine efficacy has been demonstrated for Bm86-based vaccines [14, 24]. Further experiments would have to be conducted to address this important issue by comparing the amount of ingested blood using artificial feeding systems and protease and antibody gut concentration in feeding ticks between different Boophilus species and strains.
However, two results suggested that polymorphisms in Bm86 orthologs may also contribute to differences in vaccine efficacy between B. annulatus and B. microplus: (i) Despite lower antibody titers in vaccinated cattle, vaccine efficacy on B. microplus was higher for Bm86 (Gavac; Cuban Camcord strain) than for Bm86 (Mozambique strain) and (ii) the efficacy of Bm86 vaccines was higher than that of Ba86 on B. annulatus.
The analysis of Bm86 protein sequences showed that the antigen in the Mexican (Susceptible) strain used for infestation was 97% homologous to the sequence in Gavac Cuban Camcord strain but 93% homologous to the sequence of the Mozambique strain (Fig. 2). These differences in the sequence of Bm86 may affect the efficacy of Bm86 vaccines in different strains [19]. For example, two of the predicted antigenic peptides in Bm86 (Gavac) were located on a deletion in the Bm86 (Mozambique strain) sequence but conserved in the Bm86 (Susceptible, Mexico) strain sequence (Fig. 2). The efficacy of the Ba86 vaccine on B. microplus, which was slightly higher (71.5%; Table 1) than that of the Bm86 (Mozambique strain) vaccine (70.4%; Table 1) also supports this hypothesis because the homology to the Bm86 sequence in the Mexican strain used for infestation was also higher for Ba86 (94%) than for Bm86 (Mozambique strain; 93%). However, as discussed above, differences in the production of Gavac and the Mozambique strain vaccines [23] together with cattle physiological factors [14] and antibody isotype composition [25] may also account for differences in vaccine efficacy.
Despite the possibility of higher susceptibility of B. annulatus to vaccination discussed above, the analysis of predicted antigenic regions in Ba86 and Bm86 also suggested an effect of protein sequence on vaccine efficacy. Several predicted antigenic regions were polymorphic between Bm86 and Ba86 (Fig. 2). Furthermore, Bm86 contained one predicted antigenic region not present in Ba86 (Israel strain) but present in Ba86 (Mercedes, Texas strain) (Fig. 2). The predicted antigenic regions may contain protective epitopes and thus could be involved in eliciting a protective response after vaccination. Therefore, polymorphisms in these regions could explain, at least for some Ba86 antigens, the higher efficacy of Bm86 vaccines for the control of B. annulatus.
Conclusion
The results reported herein demonstrated the efficacy of recombinant Ba86 for the control of B. annulatus and B. microplus infestations in cattle. These experiments also expanded the results of the efficacy of Bm86 vaccines by including protection against cattle infestation by new strains of B. anulatus (Mercedes, Texas, USA) and B. microplus (Susceptible, Mexico). Finally, these results suggested that physiological differences between B. microplus and B. annulatus and those encoded in the sequence of Bm86 orthologs may be responsible for the differences in susceptibility of these tick species to Bm86 vaccines.
Methods
Tick strains
The B. microplus (Susceptible, CENAPA, Mexico strain) and B. annulatus (Mercedes, Texas, USA strain) ticks were obtained from laboratory colonies maintained at the University of Tamaulipas, Mexico. Originally, these tick strains were collected from infested cattle in Tapalpa, Jalisco, Mexico and Mercedes County, Texas, USA for B. microplus and B. annulatus, respectively. Ticks were maintained during two years at the facilities of the Faculty of Veterinary Medicine, University of Tamaulipas, where several generations of tick larvae were fed on cows and collected until repletion to allow for oviposition and hatching in humidity chambers at 12 hr light: 12 hr dark photoperiod, 22–25°C and 95% relative humidity. Larvae were 15 days of age at the time of infestations.
Vaccine formulations
The recombinant Ba86 (Israeli strain) and Bm86 (Mozambique strain) were secreted in P. pastoris and purified as reported previously [23]. Protein adjuvation was made by mixing a solution of anhydromannitoletheroctodecenoate (Montanide ISA 50 V; Seppic, Paris, France) with the recombinant protein solution in batch-by-batch processes using a high-speed mixer Heidolph DIAX 900 (Heidolph Elektro, Kelheim, Germany) at 8,000 rpm and the vaccine was filled manually under sterile conditions in glass bottles of 20 ml (Wheaton, Millville, NJ, USA) at a concentration of 100 μg/2 ml dose. Quality controls were made by testing mechanical and thermal stability of vaccine emulsions as described by Canales et al. [26]. The commercial Bm86 (Cuban Camcord strain) vaccine (Gavac, Revetmex, Mexico City, Mexico) also contains 100 μg/2 ml dose of P. pastoris-derived purified recombinant protein formulated as described above.
Cattle immunization with recombinant proteins and tick infestations
Five crossbred calves per group were each immunized with 3 doses (weeks 1, 3 and 7) containing 100 μg/dose of purified recombinant proteins formulated as described above. Negative controls were injected with adjuvant/saline alone. Cattle were injected intramuscularly with 2 ml/dose using a 5 ml syringe and an 18G needle. Twelve days after the last immunization, cattle in vaccinated and control groups were infested with 10,000 B. annulatus (Mercedes, Texas, USA strain) and B. microplus (Susceptible, Mexico strain) larvae/animal applied individually to each animal in separate cotton cells attached to the back of the animals. Cattle were cared for in accordance with standards specified in the Guide for Care and Use of Laboratory Animals.
Data collection and evaluation
Adult female ticks dropping from cattle were daily collected, counted and weighted. All the collected adult female ticks were assessed for oviposition and egg fertility [27]. The personnel collecting the ticks were 'blinded' as to which group animals belonged. The efficacy of vaccine formulations was evaluated employing the following formulae [27].
Effect on the number of adult female ticks (DT) = 100 [l-(NTV/NTC)], where NTV is the number of adult female ticks in the vaccinated group and NTC is the number of adult female ticks in the control group.
Effect on tick weight (DW) = 100 [1-(WTV/WTC)], where WTV is the average adult female tick weight in the vaccinated group and WTC is the average adult female tick weight in the control group.
Effect on oviposition (DO) = 100 [1-(PATV/PATC)], where PATV is the average weight of the eggs per survived tick in the vaccinated group and PATC is the average weight of the eggs per survived tick in the control group.
Effect on egg fertility (DF) = 100 [1-(PPLOV/PPLOC)], where PPLOV is the average weight of the larvae per gram of eggs in the vaccinated group and PPLOC is the average weight of the larvae per gram of eggs in the control group.
Vaccine efficacy (E) was calculated as 100 [l-(CRT × CR0 × CRF)], where CRT = NTV/NTC, CR0 = PATV/PATC and CRF = PPLOV/PPLOC that represent the reduction in the number of adult female ticks, oviposition and egg fertility as compared to the control group, respectively.
A Student's t-test with unequal variance (P = 0.05) was used to compare the results of adult female tick number, tick weight, oviposition and egg fertility between vaccinated and control groups.
Determination of serum antibody levels by ELISA
Before each immunization and 12 (before tick infestation) and 37 days after the last immunization, blood samples were collected from each calf into sterile tubes and maintained at 4°C until arrival at the laboratory. Serum was then separated after centrifugation and stored at -20°C. Serum antibody titers were determined using an antigen-specific indirect ELISA. Purified recombinant Bm86 (Mozambique strain) and Ba86 (Israeli strain) antigens (0.1 μg/well) were used to coat ELISA plates overnight at 4°C. Sera were serially diluted to 1:10, 1:100 and 1:1000 in PBST (PBS/0.5% Tween 20, pH 7.2) and 10% fetal bovine serum (Sigma). The plates were incubated with the diluted sera for 1 hr at 37°C and then incubated with 1:10,000 rabbit anti-bovine IgG-HRP conjugates (Sigma) for 1 hr at 37°C. The color reaction was developed with 3,3',5,5'-tetramethylbenzidine (Sigma) and the OD450 nm was determined. After incubation the plates were washed with PBST. Antibody titers were considered positive when they yielded an OD450 nm value at least twice as high as the preimmune serum. Antibody titers in immunized cattle were expressed as the OD450 nm value for the highest serum dilution (1:1000) and compared between vaccinated and control cattle using an ANOVA test (P < 0.05).
Sequence analysis
The sequences of the Ba86 (Mercedes, Texas, USA; Genbank accession number FJ456927) and Bm86 (Susceptible, Mexico; FJ456928) strains were determined as described previously [23]. The protein sequences were aligned with Ba86 (Israeli strain; ABY58969), Bm86 (Mozambique strain; ABY58968) and Bm86 (Cuban Camcord strain in Gavac; [11]) using the program AlignX (Vector NTI Suite V 8.0, InforMax, Invitrogen, Carlsbad, CA, USA) with an engine based on the Clustal W algorithm [28]. Antigenic peptides (=7 residues) were predicted using the method of Kolaskar and Tongaonkar [29], with a reported accuracy of about 75% http://immunax.dfci.harvard.edu/Tools/antigenic.pl.
References
Barker SC, Murrell A: Systematics and evolution of ticks with a list of valid genus and species names. Parasitol. 2004, 129: S15-S36. 10.1017/S0031182004005207.
Estrada-Peña A, Bouattour A, Camicas JL, Guglielmone A, Horak I, Jongejan F, Latif A, Pegram R, Walker AR: The known distribution and ecological preferences of the tick subgenus Boophilus (Acari: Ixodidae) in Africa and Latin America. Exp Appl Acarol. 2006, 38: 219-235. 10.1007/s10493-006-0003-5.
Olwoch JM, Van Jaarsveld AS, Scholtz CH, Horak IG: Climate change and the genus Rhipicephalus (Acari: Ixodidae) in Africa. Onderstepoort J Vet Res. 2007, 74: 45-72.
Peter RJ, Bossche Van den P, Penzhorn BL, Sharp B: Tick, fly, and mosquito control-Lessons from the past, solutions for the future. Vet Parasitol. 2005, 132: 205-215. 10.1016/j.vetpar.2005.07.004.
Graf JF, Gogolewski R, Leach-Bing N, Sabatini GA, Molento MB, Bordin EL, Arantes GJ: Tick control: an industry point of view. Parasitol. 2004, 129: S427-S442. 10.1017/S0031182004006079.
de la Fuente J, Kocan KM: Strategies for development of vaccines for control of ixodid tick species. Parasite Immunol. 2006, 28: 275-283. 10.1111/j.1365-3024.2006.00828.x.
Sonenshine DE, Kocan KM, de la Fuente J: Tick control: further thoughts on a research agenda. Trends Parasitol. 2006, 22: 550-551. 10.1016/j.pt.2006.09.003.
Willadsen P: Tick control: thoughts on a research agenda. Vet Parasitol. 2006, 138: 161-168. 10.1016/j.vetpar.2006.01.050.
Willadsen P, Riding GA, McKenna RV, Kemp DH, Tellam RL, Nielsen JN, Lahstein J, Cobon GS, Gough JM: Immunological control of a parasitic arthropod: identification of a protective antigen from Boophilus microplus. J Immunol. 1989, 143: 1346-1351.
Rand KN, Moore T, Sriskantha A, Spring K, Tellam R, Willadsen P, Cobon G: Cloning and expression of a protective antigen from the cattle tick Boophilus microplus. Proc Natl Acad Sci USA. 1989, 86: 9657-9661. 10.1073/pnas.86.24.9657.
Rodríguez M, Rubiera R, Montesinos R, Cremata JA, Falcón V, Sanchez G, Bringas R, Cordovés C, Valdés M, Lleonart R, Herrera L, de la Fuente J: High level expression of the B. microplus Bm86 antigen in the yeast P. pastoris forming highly inmunogenic particles for cattle. J Biotechnol. 1994, 33: 135-146. 10.1016/0168-1656(94)90106-6.
de la Fuente J, Kocan KM: Advances in the identification and characterization of protective antigens for development of recombinant vaccines against tick infestations. Expert Rev Vaccines. 2003, 2: 583-593. 10.1586/14760584.2.4.583.
de la Fuente J, Almazán C, Canales M, Pérez de la Lastra JM, Kocan KM, Willadsen P: A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim Health Res Rev. 2007, 8: 23-28. 10.1017/S1466252307001193.
de la Fuente J, Rodríguez M, Redondo M, Montero C, García-García JC, Méndez L, Serrano E, Valdés M, Enríquez A, Canales M, Ramos E, de Armas CA, Rey S, Rodríguez JL, Artiles M, García L: Field studies and cost-effectiveness analysis of vaccination with Gavac™ against the cattle tick Boophilus microplus. Vaccine. 1998, 16: 366-373. 10.1016/S0264-410X(97)00208-9.
Rodríguez Valle M, Méndez L, Valdez M, Redondo M, Espinosa CM, Vargas M, Cruz RL, Barrios HP, Seoane G, Ramírez ES, Boué O, Vigil JL, Machado H, Nordelo CB, Piñeiro MJ: Integrated control of Boophilus microplus ticks in Cuba based on vaccination with the anti-tick vaccine Gavac. Exp Appl Acarol. 2004, 34: 375-382.
Fragoso H, Hoshmand Rad P, Ortiz M, Rodríguez M, Redondo M, Herrera L, de la Fuente J: Protection against Boophilus annulatus infestations in cattle vaccinated with the B. microplus Bm86-containing vaccine Gavac. Vaccine. 1998, 16: 1990-1992. 10.1016/S0264-410X(98)00116-9.
de la Fuente J, Rodríguez M, Garcia-Garcia JC: Immunological control of ticks through vaccination with Boophilus microplus gut antigens. Ann N Y Acad Sci. 2000, 916: 617-621.
de Vos S, Zeinstra L, Taoufik O, Willadsen P, Jongejan F: Evidence for the utility of the Bm86 antigen from Boophilus microplus in vaccination against other tick species. Exp Appl Acarol. 2001, 25: 245-261. 10.1023/A:1010609007009.
García-García JC, González IL, González DM, Valdés M, Méndez L, Lamberti J, D'Agostino B, Citroni D, Fragoso H, Ortiz M, Rodríguez M, de la Fuente J: Sequence variations in the Boophilus microplus Bm86 locus and implications for immunoprotection in cattle vaccinated with this antigen. Exp Appl Acarol. 1999, 23: 883-895. 10.1023/A:1006270615158.
García-García JC, Montero C, Redondo M, Vargas M, Canales M, Boué O, Rodríguez M, Joglar M, Machado H, González IL, Valdés M, Méndez L, de la Fuente J: Control of ticks resistant to immunization with Bm86 in cattle vaccinated with the recombinant antigen Bm95 isolated from the cattle tick, Boophilus microplus. Vaccine. 2000, 18: 2275-2287. 10.1016/S0264-410X(99)00548-4.
de la Fuente J, García-García JC, González DM, Izquierdo G, Ochagavia ME: Molecular analysis of Boophilus spp. (Acari: Ixodidae) tick strains. Vet Parasitol. 2000, 92: 209-222. 10.1016/S0304-4017(00)00308-3.
Sossai S, Peconick AP, Sales-Junior PA, Marcelino FC, Vargas MI, Neves ES, Patarroyo JH: Polymorphism of the bm86 gene in South American strains of the cattle tick Boophilus microplus. Exp Appl Acarol. 2005, 37: 199-214. 10.1007/s10493-005-3262-7.
Canales M, Pérez de la Lastra JM, Naranjo V, Nijhof AM, Hope M, Jongejan F, de la Fuente J: Expression of recombinant Rhipicephalus (Boophilus) microplus, R. annulatus and R. decoloratus Bm86 orthologs as secreted proteins in Pichia pastoris. BMC Biotechnol. 2008, 8: 14-10.1186/1472-6750-8-14.
Cobon G, Hungerford J, Woodrow M, Smith D, Willadsen P: Vaccination against Boophilus microplus. Tha Australian field experience. Recombinant Vaccines for the Control of Cattle Tick. Edited by: de la Fuente J. 1995, Elfos Scientiae, La Habana, Cuba, 163-176.
Rodríguez M, Montero C, Machado H, Joglar M, de la Fuente J, Garcia-Garcia JC: The evaluation of yeast derivatives as adjuvants for the immune response to the Bm86 antigen in cattle. BMC Biotechnol. 2001, 1: 2-10.1186/1472-6750-1-2. Erratum in BMC Biotechnol 2001, 1: 3
Canales M, Enriquez A, Ramos E, Cabrera D, Dandie H, Soto A, Falcon V, Rodríguez M, de la Fuente J: Large-scale production in Pichia pastoris of the recombinant vaccine Gavac™ against cattle ticks. Vaccine. 1997, 15: 414-422. 10.1016/S0264-410X(96)00192-2.
de la Fuente J, Rodríguez M, Montero C, Redondo M, García-García JC, Méndez L, Serrano E, Valdés M, Enríquez A, Canales M, Ramos E, Boué O, Machado H, Lleonart R: Vaccination against ticks (Boophilus spp.): the experience with the Bm86-based vaccine Gavac™. Genetic Analysis: Biomolecular Engineering. 1999, 15: 143-148. 10.1016/S1050-3862(99)00018-2.
Thompson JD, Higgins DG, Gibson TJ, CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighing, positions-specific gap penalties and weight matrix choice. Nucl Acid Res. 1994, 22: 4673-4680. 10.1093/nar/22.22.4673.
Kolaskar AS, Tongaonkar PC: A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276: 172-174. 10.1016/0014-5793(90)80535-Q.
Acknowledgements
We thank Peter Willadsen (CSIRO Livestock Industries, Queensland, Australia) for suggestions to the experiments presented in this paper. Vladimir Guerrero, Baldomero Ibarra and Urbano Martinez (Universidad Autónoma de Tamaulipas, Mexico) are acknowledged for technical assistance. This work was supported by the Wellcome Trust under the Animal Health in the Developing World initiative through project 0757990 entitled "Adapting recombinant anti-tick vaccines to livestock in Africa" and the Consejería de Educación y Ciencia, JCCM, Spain (project PAI06-0046-5285) and was facilitated through the Integrated Consortium on Ticks and Tick-borne Diseases (ICTTD-3), financed by the International Cooperation Program of the European Union, coordination action project No. 510561.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
MC carried out the expression, fermentation, protein purification and vaccine formulation. CA conducted the vaccine trial and determined serum antibody levels by ELISA. VN determined the sequence of Ba86 and Bm86. JF did sequence analyses. JF and FJ conceived the study, and participated in its design and coordination and drafted the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Canales, M., Almazán, C., Naranjo, V. et al. Vaccination with recombinant Boophilus annulatus Bm86 ortholog protein, Ba86, protects cattle against B. annulatus and B. microplus infestations. BMC Biotechnol 9, 29 (2009). https://doi.org/10.1186/1472-6750-9-29
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1472-6750-9-29