
Abstract
Background
Statins, potent compounds that inhibit cholesterol synthesis in the liver have been reported to induce bone formation, both in tissue culture and in rats and mice. To re-examine potential anabolic effects of statins on bone formation, we compared the activity of simvastatin (SVS) to the known anabolic effects of PTH in an established model of ovariectomized (OVX) Swiss-Webster mice.
Methods
Mice were ovariectomized at 12 weeks of age (T0), remained untreated for 5 weeks to allow development of osteopenia (T5), followed by treatment for 8 weeks (T13). Whole, trabecular and cortical femoral bone was analyzed by micro-computed tomography (micro CT). Liquid chromatography/mass spectrometry (LC/MS) was used to detect the presence of SVS and its active metabolite, simvastatin β-hydroxy acid (SVS-OH) in the mouse serum.
Results
Trabecular BV/TV at T13 was 4.2 fold higher in animals treated with PTH (80 micro-g/kg/day) compared to the OVX-vehicle treated group (p < 0.001). However, the same comparison for the SVS-treated group (10 mg/kg/day administered by gavage) showed no significant difference (p = NS). LC/MS detected SVS and SVS-OH in mouse serum 20 minutes after gavage of 100 mg SVS. A serum osteocalcin assay (OC) demonstrated that neither bone formation nor osteoblast activity is significantly enhanced by SVS treatment in this in vivo study.
Conclusions
While PTH demonstrated the expected anabolic effect on bone, SVS failed to stimulate bone formation, despite our verification by LC/MS of the active SVS-OH metabolite in mouse serum. While statins have clear effects on bone formation in vitro, the formulation of existing 'liver-targeted' statins requires further refinement for efficacy in vivo.
Similar content being viewed by others


Background
Statins, potent compounds that inhibit cholesterol synthesis in the liver by blocking 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA), have been reported to induce osteoblast activity and lead to bone formation, both in tissue culture and in rats and mice [1–6]. These findings, if confirmed and extended to include humans, have the potential to introduce immediate new treatment options to patients with low bone mass and osteoporosis. Such treatment is readily available, since an estimated 3 million patients in the US alone take statins daily to treat hyperlipidemia with few side effects [7]. Osteoporosis is increasingly recognized as an eminent public health problem, as the mean age of populations in most Western countries increases and costs related to osteoporosis are expected to grow six-fold by the year 2040 [8]. Data from the National Health and Nutrition Examination Survey, using a definition of osteoporosis developed by the World Health Organization, estimate that in the United States up to 20% of Caucasian women over 50 have osteoporosis and up to 50% have low bone mass. Non-caucasian women and men tend to have lower rates of osteoporosis but contribute up to 25% of the fractures annually [9].
The only bone anabolic agent known to increase bone mineral density in both animal models and clinical settings is human parathyroid hormone 1–34 (PTH), which has recently been approved by the FDA in patients with advance osteoporosis [10]. Bone-related basic and clinical research has been searching intensively for alternative anabolic agents for the treatment of osteoporosis.
Due to the potential clinical impact of statins as a treatment option for patients with low bone mass and osteoporosis, it is essential to evaluate its anabolic activity in several in vitro and experimental animal models. We have recently reported the anabolic action of PTH in an ovariectomized (OVX) Swiss-Webster mouse model, which like the rat model, displays a substantial amount of trabecular bone in its femoral metaphyses and a quantifiable loss of bone following OVX [11]. The aim of this study was to investigate the effect of simvastatin on the microarchitecture of bone in an established mouse model of osteoporosis, and compare its anabolic action with PTH. We also verified the presence of systemically administered simvastatin in the mouse serum, as this has previously not been reported and is likely to be of critical importance when evaluating the effect of simvastatin on bone.
Methods
Animals
72 eleven-week-old virgin female Swiss-Webster mice were purchased from Taconic Farms (Germantown, NY) and maintained at the animal research facility at the Beth Israel Deaconess Medical Center. Animals were fed Purina Formulab Diet containing 1% Ca (Formulab, Richmond, IN) and water ad libitum throughout the experiment. Mice were sacrificed by CO2 inhalation.
Experimental protocol
The experimental protocol was approved by the Institutional Animal Care and Use Committee of the Beth Israel Deaconess Medical Center. The study design is shown schematically in Figure 1. The mice were randomly subdivided into five groups of bilaterally ovariectomized (OVX) and three groups of SHAM OVX animals, with 9 animals in each group. A group of each of OVX and SHAM animals was sacrificed one week postoperatively (T1/OVX and T1/SHAM, respectively) and served as baseline controls. Four additional weeks were allowed to pass before initiation of treatment in the remaining groups in order to permit significant bone loss to occur in the OVX animals. At this time, one group of OVX (T5/OVX) and one group of sham OVX mice (T5/SHAM) were sacrificed to evaluate pre-treatment bone loss.

Schematic representation of the experimental design
An eight-week daily treatment (5 days a week) consisting of daily gavage of either 10 mg/kg of simvastatin (Apin Chemical Limited UK, Oxfordshire, England) or vehicle (VEH) only (0.5% methylcellulose in H2O) was administered to the T13/OVX/SVS and T13/OVX/VEH groups, respectively.
An additional control group of OVX mice received daily treatment (5 days a week) consisting of subcutaneous (sc) injections of 80 μg/kg per day of hPTH (1–34) (Advanced ChemTech, Louisville, KY, USA). All groups were sacrificed after completion of the 13 week study.
Micro-computed tomography analysis
For a detailed qualitative and quantitative 3-D evaluation, whole femoral bones were examined by a desktop μCT system (μCT 20, Scanco Medical AG, Bassersdorf, Switzerland) equipped with a 10 μm focal spot microfocus X-ray tube as a source [12]. For image acquisition, the specimen was mounted on a turntable shifted automatically in an axial direction over 216° (180° plus half fan angle on either side), taking 600 projections. To scan the entire femoral width (3.4–5.1 mm), including the femoral head, a total of 100–150 micro-tomographic slices were acquired at a 34 μm slice increment. CT images were reconstructed in 512 × 512 pixel matrices using a standard convolution-backprojection procedure with a Shepp and Logan filter. Images were stored in 3-D arrays with an isotropic voxel size of 34 μm. A constrained 3-D Gaussian filter was used to partly suppress the noise in the volumes and the mineralized tissue was segmented from soft tissues by a global thresholding procedure [13]. Morphometric parameters were determined using a direct 3-D approach [14] in three different pre-selected analysis regions: whole bone (including the articular ends), secondary spongiosa in the distal metaphysis, and diaphyseal cortical bone. Parameters determined in the metaphyseal trabecular bone included bone volume density (BV/TV), bone surface density (BS/TV), trabecular number (Tb.N), trabecular thickness (Tb.Th), trabecular spacing (Tb.Sp) and connectivity density. Cortical volume (TV), percent marrow volume (%MV) and cortical thickness (Ct.Th) were assessed in a 1 mm-thick slab in the mid-diaphysis.
Mass Spectrometry
Bond Elut C8 SPE cartridges (1 ml/100 mg) were purchased from Varian/Sample Preparation Products (Harbor City, CA, USA). ESI-MS (electrospray ionization mass spectrometry) were recorded on a Micromass Platform LCZ system equipped with an electrospray ionization source and a Waters 2690 Separation Module. For analytical RP-HPLC (Reverse-phase high performance liquid chromatography Vydac C18 columns were used; the solvent system used included Water/TFA (1000:1, eluent A) and Acetonitrile/TFA (1000:1, eluent B). Linear elution-gradient was used ("Waters Gradient 6", t = 0 min, 10% B; t = 1 min, 45% B; t = 21 min, 60% B; t = 26 100% MeOH; t = 35 min, 10% B). Flow rates were 0.2 ml/min and the detection wavelengths were set to 220 and 240 nm.
Sample preparation
All sample preparation procedures were carried out in an ice-water bath. Stock solution of simvastatin was prepared in ethanol and of simvastatin hydroxy acid in methanol-water (90:10) at 4°C. Calibration standards were prepared fresh for each analysis, by dilutions in methanol-water (70:30). The doses of the standards at the respective points on the calibration graphs were 1, 10, 30, 60, 100, 150 ng. Recovery of simvastatin and of simavastatin hydroxy acid from mouse plasma following the extraction procedure below, was assessed by comparison of samples at three doses (30, 80 and 150 ng), which were used to spike 0.1 ml mouse plasma followed by extraction and injection into LC-MS, to similar doses directly injected to the LC-MS. The quantitation was based on integration of the area under the respective peaks in the UV spectra obtained at 240 nm (approximately the λmax for both simvastatin and simavastatin hydroxy acid). Each of the samples from the in vivo tests (0.15 ml), or the recovery test above, was diluted with 0.4 ml of water prior to extraction.
Extraction procedure
This procedure was based on the previously published protocol by Wu et al. [15], implementing minor modifications. Each solid-phase extraction cartridge (1 ml/100 mg) was conditioned with Methanol-water (50:50). The diluted sample (see Sample preparation) was loaded on to each cartridge, followed by low vacuum (1 min), and each cartridge was subsequently washed with 1 ml H2O, 1 ml 5% formic acid, 1 ml H2O and dried for 1 min. Each cartridge was then eluted with 1 ml of methanol-water (70:30) solution and with 1 ml of acetonitrile, followed by low vacuum (1 min). The combined eluates were evaporated to dryness in a SpeedVac at 45°C. The dry residues were reconstituted in 100 μl of ammonium acetate (10 mm, pH 4.0)-acetonitrile (30:70). The reconstituted extracts were transferred into autosampler vials and 50 μl of each sample were injected on to the LC column (equivalent to 75 μl of plasma sample).
Osteocalcin assay
Blood was sampled prior to sacrifice to determine serum levels of osteocalcin (OC). Serum intact OC was measured by a two-site immunoradiometric assay (IRMA kit, Immutopics Inc., San Clemente, CA) against the mid-region C-terminal and amino-terminal position of mouse OC with a known sensitivity of 0.1 ng/mL. Intra- and interassay CVs were 2.3% and 4.4%, respectively.
Statistical Analysis
A power analysis was conducted in planning the experiments to determine the sufficient number of mouse femurs in this investigation. A total sample size of 72 animals (n = 9 per treatment condition) was required to provide 90% power (α = 0.05, β = 0.1) to detect a significant difference of 1 standard deviation (effect size = 1.0) between the Simvastatin, PTH, OVX, and SHAM treatments with respect to each trabecular and cortical bone parameter using one-way analysis of variance (ANOVA) with a two-tailed significance level of 0.05. Sample size calculations were determined using the nQuery Advisor software program (version 4.0, Statistical Solutions, Boston, MA).
All continuous variables were checked for normality using the Kolmogorov-Smirnov goodness-of-fit test and were found to follow a normal distribution closely [16]. Therefore, results are reported in terms of means ± standard deviation (SD). Treatment groups were compared with respect to trabecular and cortical bone parameters obtained by μCT by use of analysis of variance (ANOVA) with the Fisher least significant difference (LSD) method for post-hoc multiple testing [17]. A two-tailed value of p < 0.05 was established as the criterion for statistical significance. Statistical analysis was performed using the SPSS (version 11.0, SPSS Inc., Chicago, IL) and SAS (version 6.12, SAS Institute, Cary, NC) software packages.
Results
Effects of OVX on bone loss
Consistent with the findings in our recent study [11], μCT analyses of femoral trabecular and cortical bone showed little change over the duration of the study in any of the structural variables for SHAM OVX Swiss-Webster mice (groups T1/SHAM, T5/SHAM, T13/SHAM). OVX mice demonstrated significantly lower trabecular bone volume density (BV/TV) compared to SHAM controls at 1 week (15 ± 6% vs. 26 ± 5%, P < 0.001) 5 weeks (5 ± 3% vs. 28 ± 12%, P < 0.001), and showed a reduction of 85% at 13 weeks (5 ± 4% vs. 34 ± 11%, P < 0.001, Table 1). In addition, trabecular thickness, spacing and number were significantly different in OVX mice compared to SHAM controls at 5 and 13 weeks (all P < 0.001). However, cortical parameters including total volume, marrow volume/total volume, and thickness were not significantly different between OVX and SHAM controls throughout the 13-week study course (all P > 0.20, Table 1).
Effect of PTH and SVS treatment on bone
At 13 weeks, trabecular bone volume density (BV/TV) in mice treated with PTH alone (T13/OVX/PTH) was 4.2-fold higher than vehicle-treated animals (T13/OVX/VEH) (Table 2). PTH treatment was associated with significant increases in Tb.Th and Tb.N as well as an expected decrease in Tb.Sp. In addition, significant differences in cortical %MV/TV and thickness were observed at 13 weeks (P < 0.01, Table 2).
In comparison, trabecular BV/TV in SVS-treated animals at 13 weeks (T13/OVX/SVS) was not significantly increased over that observed in vehicle-treated animals (T13/OVX/VEH) (Figure 2). Furthermore, none of the other trabecular bone parameters (Tb.Th, Tb.Sp and Tb.N) revealed a significant difference after 13 weeks of SVS treatment when compared to T13/OVX/VEH animals, (all P = NS, Table 2).

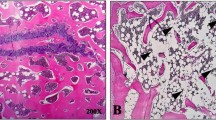
3D μCT images of distal femoral metaphyseal trabecular bone of 13 week SHAM, OVX-VEH-treated, OVX PTH treated, OVX SVS treated Swiss-Webster mice. Images were obtained from animals with median cancellous BV/TV values.
Similarly, whole bone volume density at T13 was higher in animals treated with PTH compared to the OVX group (76 ± 4% vs. 67 ± 3%, p < 0.001), whereas SVS treatment had no significant effect (68 ± 3% vs. 67 ± 3%, p = NS). In addition, cortical thickness evaluated at T13 was higher in the PTH treatment group compared to the OVX group (0.31 ± 0.03 mm vs. 0.26 ± 0.02 mm, p < 0.001), whereas SVS treatment exhibited no effect (0.27 ± 0.01 mm vs. 0.26 ± 0.02 mm, p = NS, Table 2).
Osteocalcin
Mean serum osteocalcin (OC) of SHAM and OVX treated animals did not change significantly when compared at 1, 5 and 13 weeks. PTH treated mice, however revealed an increase (596 ± 138 ng/ml) over T13/OVX/VEH animals (252 ± 57 ng/ml, P < 0.001, Figure 2). SVS treatment at 13 weeks, however, caused no significant increases in OC compared to T13/OVX/VEH animals (309 ± 74 ng/ml vs. 252 ± 57 ng/ml, p = NS, Figure 3).

Mean Serum Osteocalcin levels at 13 weeks. While PTH treatment (596 ± 138 ng/ml) was significantly different (P < 0.001,*) from OVX (252 ± 57 ng/ml) and SVS (309 ± 74 ng/ml), SVS treatment was not significantly different (NS) from OVX vehicle treated animals after 13 weeks.
Detection of the active Simvastatin hydroxy acid metabolite in mouse serum after gavage administration
In this study, SVS was administered by gavage, and requires hepatic conversion to the metabolically active β-hydroxy acid (SVS-OH). Because SVS failed to have any effect on any static μCT parameter measured at either site in mouse femur, we conducted separate experiments designed to detect the presence of SVS-OH in mouse serum after SVS gavage by liquid chromatography/mass spectrometry (LC/MS).
Reverse-phase high performance liquid chromatography (RP-HPLC) tandem electrospray ionization mass spectrometry (ESI-MS) method published by Wu et al. [15] was used to analyze samples of mouse serum for the presence of SVS and SVS-OH. The identity of the eluted peak in RP-HPLC was established by the determination of the mass associated with it by ESI-MS. The retention times for SVS and SVS-OH were 19.5 ± 0.5 and 15.35 ± 0.20 min, respectively. Threshold of detection on the LC-MS at λ240 nm (AUFS = 0.1), the most sensitive wavelength, for both SVS and the SVS-OH was ~1 ng. Calibration curves for both the SVS and the SVS-OH extracted from the area under the curve in the HPLC profile was linear in the range of 1–150 ng. These curves were used to quantitate the drug and metabolite levels in mouse plasma. Based on the spiking of mouse plasma ex-vivo with known amounts of drug and subjecting the samples for the extraction procedure, the efficiency of drugs recovery was estimated to be 60–75% for SVS and 70–85% for SVS-OH. Figure 4 is a representative example of the LC-MS analysis of a sample collected from a mouse 30 min after treatment with 100 mg/kg SVS. In this sample both the RP-HPLC tracing and the ESI-MS analysis of the peaks corresponding to the drug and its metabolite confirm the presence of both in the serum.

Liquid chromatography/mass spectrometry (LC/MS) for SVS (M + Na+ = 442 kD) and SVS-OH (M + Na+ = 460 kD) of mouse serum 30 min after gavage with 100 mg/kg SVS.
Both residual SVS and its in situ-generated metabolite SVS-OH were detected in the mouse plasma as early as 10 min after gavage administration of 100 mg/kg of SVS (the shortest sampling interval). The levels of SVS and SVS-OH in the plasma 10 min after administration were 97.3 and 15.3 ng/μl, respectively. These findings establish the bioavailability of the drug following this route of administration.
Discussion
The aim of this study was to investigate the effect of Simvastatin on bone in an established mouse model of osteoporosis. This Swiss-Webster outbred mouse strain utilized in this study has proven to be a valuable model for investigating the efficacy of both anabolic and anti-resorptive therapies for the treatment of osteoporosis. Here, we have used PTH as a positive control [11], and compared its effects to SVS. While PTH demonstrated the expected anabolic effect on bone, SVS failed to stimulate bone formation, despite our verification by LC/MS of the active SVS-OH metabolite in mouse serum.
The SVS-OH metabolite is first detectable in mouse serum by 10 minutes after gavage administration, and serum levels of SVS-OH continued to be observed by both mass-spectrometric and UV measurements through the last serum samples collected at 50 minutes post-gavage. Other experiments failed to detect SVS-OH at longer post-gavage timepoints of 3 and 24 hours. These data are in general agreement with studies in both rats and canines that calculate peak serum levels of SVS-OH at 40–60 minutes after gavage administration [18]. Thus, although we failed to document any bone anabolic activity of SVS in a mouse model of OVX-induced osteoporosis, LC/MS analysis of the SVS-OH metabolite in mouse serum document the efficacy of our gavage administration of SVS to experimental mice.
Extrapolating from mice to humans, the dose of SVS administered in this study was approximately 400-fold higher than the average dose given to humans. The results suggest that Simvastatin does not enhance bone formation in this mouse model in vivo, despite our ability to verify the presence of the SVS-OH metabolite in mouse serum by LC/MS analysis. Furthermore, the osteocalcin assay did not suggest any increase in osteoblast activity in SVS treated mice, consistent with its failure to increase bone density.
It is generally accepted that the hydroxy acid form of statins are potent inhibitors of 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase, an enzyme that plays a critical role in cholesterol metabolism, where they block substrate accessibility to HMG-CoA reductase, effectively subverting cholesterol levels of HDL [19–21]. In addition to lowering cholesterol, statins seem to have a number of additional effects, such as the nitric oxide-mediated promotion of new blood vessel growth [22], protection against oxidative modification of low-density lipoprotein, as well as anti-inflammatory effects and a reduction in C-reactive protein levels [23]. All statins limit cholesterol biosynthesis by inhibiting the committed step in the biosynthesis of isoprenoids and sterols [24].
Considering the enormous potential impact of statins on the treatment of osteoporosis, several groups have further investigated the skeletal effects of statin use. Several groups have failed to confirm the initially reported anabolic effect of systemically administered statins in vivo in rats [25–27]. One obvious criticism of the negative data in these studies is that they did not verify the actual presence of statin in the animal serum. This limitation may be of importance, as Crawford et al [27] could detect an increase in cortical bone when statin was administered locally. Our study demonstrated the presence of systemically administered the SVS-OH metabolite in the mouse serum by LC/MS analysis.
The exact mechanism by which statins perform their effects on bone cell function is a central issue. It has been hypothesized that statins mediate their effects by increasing the expression in bone of the growth factor bone morphogenic protein-2 (BMP-2) which in turn leads to osteoblast differentiation and bone formation [2]. This mechanism of action has been addressed by Baumann et al. in rats, where the anabolic effect of lovastatin on bone was at least partially due to their inhibiting effect on osteoclast development[28]. Similarly, in the mouse calvaria model Gasser et al. failed to detect any anabolic activity of statins in bone and suggested that the disruption of the capillary integrity and local bleeding might explain some of the previously reported bone responses [26].
Several studies in humans reported a decreased risk of fracture after statin treatment [3, 29, 30]. However larger human studies failed to confirm statins beneficial effect on bone. LaCroix et al. found the risk of hip and arm fracture to be similar among women who used statins and those who did not take lipid lowering medication [31]. Van Staa et al. examining 218,062 individuals with a fracture in the General Practice Database in the United Kingdom, and found no relationship between statin use and nonspine fracture [32]. Interestingly, a report by Meier [3] had used the exact same database as van Staa et al. [32], but sampled a much larger cohort (218,062 individuals with fracture) than Meier (3940), and in addition, selected an age and gender matched control patient for every fracture patient. The clinical data available at the moment does not allow the conclusion that statin treatment has a postive relationship with fracture risk.
Similarly as described in the clinical trials, the early animal studies reported data supportive of the initial findings [1, 4–6, 33, 34]. These results however, were not substantiated in later studies in rodents, where SVS had no or little effect on bone mineral density [26]. However, none of the studies has verified the actual presence of the active SVS-OH metabolite in the animal serum, as shown in this study. In combination with the osteocalcin data, this report provides strong evidence that simvastatin does not have an effect on bone in an established Swiss Webster mouse model of OVX-induced osteoporosis.
Conclusion
In summary, the currently available data on the effects of statins on bone has done little to confirm the finding that statin therapy increases bone mass and thereby lowers fracture risk. Few of the studies in animals and humans have been able to show a predictable beneficial effect on bone mass and bone turnover. This report verifies the presence of the active SVS-OH metabolite in the mouse serum yet failed to demonstrate any beneficial effect on bone. While statins have clear effects on bone formation in vitro, the formulation of existing 'liver-targeted' statins may require further refinement for efficacy on bone formation in vivo.
Authors contributions
D. von Stechow: participated in the study coordination, perfomed the microCT analysis and drafted the manuscript
S. Fish: carried out the animal experiments
D. Yahalom: carried out the mass spec analsysis
I. Bab: participated in the design of the study
M. Chorev: participated in the mass spec analysis
R. Müller: participated in the microCT analysis
J.M. Alexander: conceived of the study, and participated in its design and coordination, and edited the manuscript
References
Banu J, Kalu DN: Effects of cerivastatin and parathyroid hormone on the lumbar vertebra of aging male Sprague-Dawley rats. Bone. 2002, 31: 173-179. 10.1016/S8756-3282(02)00803-7.
Mundy G, Garrett R, Harris S, Chan J, Chen D, Rossini G, Boyce B, Zhao M, Gutierrez G: Stimulation of bone formation in vitro and in rodents by statins. Science. 1999, 286: 1946-1949. 10.1126/science.286.5446.1946.
Meier CR, Schlienger RG, Kraenzlin ME, Schlegel B, Jick H: HMG-CoA reductase inhibitors and the risk of fractures. Jama. 2000, 283: 3205-3210. 10.1001/jama.283.24.3205.
Oxlund H, Dalstra M, Andreassen TT: Statin given perorally to adult rats increases cancellous bone mass and compressive strength. Calcif Tissue Int. 2001, 69: 299-304.
Skoglund B, Forslund C, Aspenberg P: Simvastatin improves fracture healing in mice. J Bone Miner Res. 2002, 17: 2004-2008.
Whang K, Zhao M, Qaio M, Rossini G, Horn D, Garrett IR, Mundy GR, Chen D: Administration of lovastatin locally in low doses in a novel; delivery system induces prolonged bone formation. J Bone Miner Res. 2000, 15 (Suppl): S225-
Cummings SR, Bauer DC: Do statins prevent both cardiovascular disease and fracture?. Jama. 2000, 283: 3255-3257. 10.1001/jama.283.24.3255.
Schneider EL, Guralnik JM: The aging of America. Impact on health care costs. Jama. 1990, 263: 2335-2340. 10.1001/jama.263.17.2335.
Looker AC, Johnston CC, Wahner HW, Dunn WL, Calvo MS, Harris TB, Heyse SP, Lindsay RL: Prevalence of low femoral bone density in older U.S. women from NHANES III. J Bone Miner Res. 1995, 10: 796-802.
Neer RM, Arnaud CD, Zanchetta JR, Prince R, Gaich GA, Reginster JY, Hodsman AB, Eriksen EF, Ish-Shalom S, Genant HK, Wang O, Mitlak BH: Effect of parathyroid hormone (1–34) on fractures and bone mineral density in postmenopausal women with osteoporosis. N Engl J Med. 2001, 344: 1434-1441. 10.1056/NEJM200105103441904.
Alexander JM, Bab I, Fish S, Müller R, Uchiyama T, Gronowicz G, Nahounou M, Zhao Q, White DW, Chorev M, Gazit D, Rosenblatt M: Human parathyroid hormone 1–34 reverses bone loss in ovariectomized mice. J Bone Miner Res. 2001, 16: 1665-1673.
Rüegsegger P, Koller B, Müller R: A microtomographic system for the nondestructive evaluation of bone architecture. Calcif Tissue Int. 1996, 58: 24-29. 10.1007/s002239900006.
Müller R, Rüegsegger P: Micro-tomographic imaging for the nondestructive evaluation of trabecular bone architecture. Stud Health Technol Inform. 1997, 40: 61-79.
Hildebrand T, Laib A, Müller R, Dequeker J, Rüegsegger P: Direct three-dimensional morphometric analysis of human cancellous bone: microstructural data from spine, femur, iliac crest, and calcaneus. J Bone Miner Res. 1999, 14: 1167-1174.
Wu Y, Zhao J, Henion J, Korfmacher WA, Lapiguera AP, Lin CC: Microsample determination of lovastatin and its hydroxy acid metabolite in mouse and rat plasma by liquid chromatography/ionspray tandem mass spectrometry. J Mass Spectrom. 1997, 32: 379-387. 10.1002/(SICI)1096-9888(199704)32:4<379::AID-JMS461>3.0.CO;2-9.
Conover WJ: Practical nonparametric statistics. 1999, New York: John Wiley, 3
Armitage P, Berry G: Statistical methods in medical research. 1994, Cambridge, MA: Blackwell Science, 3
Vickers S, Duncan CA, Chen IW, Rosegay A, Duggan DE: Metabolic disposition studies on simvastatin, a cholesterol-lowering prodrug. Drug Metabolism and Disposition. 1990, 18: 138-145.
Istvan ES, Deisenhofer J: Structural mechanism for statin inhibition of HMG-CoA reductase. Science. 2001, 292: 1160-1164. 10.1126/science.1059344.
Altieri DC: Statins' benefits begin to sprout. J Clin Invest. 2001, 108: 365-366. 10.1172/JCI200113556.
Rao S, Porter DC, Chen X, Herliczek T, Lowe M, Keyomarsi K: Lovastatin-mediated G1 arrest is through inhibition of the proteasome, independent of hydroxymethyl glutaryl-CoA reductase. Proc Natl Acad Sci U S A. 1999, 96: 7797-7802. 10.1073/pnas.96.14.7797.
Kureishi Y, Luo Z, Shiojima I, Bialik A, Fulton D, Lefer DJ, Sessa WC, Walsh K: The HMG-CoA reductase inhibitor simvastatin activates the protein kinase Akt and promotes angiogenesis in normocholesterolemic animals. Nat Med. 2000, 6: 1004-1010. 10.1038/79510.
Davignon J, Laaksonen R: Low-density lipoprotein-independent effects of statins. Curr Opin Lipidol. 1999, 10: 543-559. 10.1097/00041433-199912000-00010.
Corsini A, Maggi FM, Catapano AL: Pharmacology of competitive inhibitors of HMG-CoA reductase. Pharmacol Res. 1995, 31: 9-27. 10.1016/1043-6618(95)80042-5.
Yao W, Li CY, Farmer RW, Chen JL, Mo A, Cooper R, Chmielewski P, Setterberg RB, Jee WSS, Lundy MW: Simvastatin did not prevent bone loss in ovariectomized rats. J Bone Miner Res. 2001, 16: S294-
Gasser JA: Fluvastatin and cervastatin are not anabolic for bone after local and systemic administration of non-toxic doses in mice and rats. J Bone Miner Res. 2001, 16: S295-
Crawford DT, Qi H, Chidesy-Frink KL, Thompson DD, Ke HZ: Statin increases cortical bone in young male rats by single, local administration but fails to restore bone in ovariectomized (OVX) rats by daily systemic administration. J Bone Miner Res. 2001, 16: S295-
Baumann AP, Grasser W, Petras S, Harwood HJ, Thompson DD, Paralkar V: Inhibition of osteoclast formation by statins. J Bone Miner Res. 2001, 16: S507-
Chan KA, Andrade SE, Boles M, Buist DS, Chase GA, Donahue JG, Goodman MJ, Gurwitz JH, LaCroix AZ, Platt R: Inhibitors of hydroxymethylglutaryl-coenzyme A reductase and risk of fracture among older women. Lancet. 2000, 355: 2185-2188. 10.1016/S0140-6736(00)02400-4.
Wang PS, Solomon DH, Mogun H, Avorn J: HMG-CoA reductase inhibitors and the risk of hip fractures in elderly patients. Jama. 2000, 283: 3211-3216. 10.1001/jama.283.24.3211.
LaCroix AZ, Cauley JA, Jackson R, McGowan J, Pettinger M, Hsia J, Chen Z, Lewis C, Bauer DC, Daugherty S, McNeeley SG, Passaro M: Does statin use reduce the risk of fracture in postmenopausal women? Results from the Women's Health Initiative Observational Study (WHIOS). J Bone Miner Res. 2000, 15 (Suppl): S155-
van Staa TP, Wegman SLJ, F. dV, Leufkens HGM, Cooper C: Use of Statins and risk of fracture. J Bone Miner Res. 2000, 16 (Suppl): S155-
Oxlund H: Simvastatin given perorally increases the cortical bone formation rate in adult rats. J Bone Miner Res. 1999, 15 (Suppl): S313-
Garrett IR, Esparza J, Chen D, Zhao M, Guiterrez G, Escobedo A, Horn D, Mundy GR: Statins mediate their effects on osteoblasts by inhibition of HMG-CoA reductase and ultimately BMP-2. J Bone Miner Res. 2000, 15 (Suppl):
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2474/4/8/prepub
Acknowledgements
This work was supported in part by a grant from the William F Milton Fund at Harvard Medical School.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
None declared.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
von Stechow, D., Fish, S., Yahalom, D. et al. Does simvastatin stimulate bone formation in vivo?. BMC Musculoskelet Disord 4, 8 (2003). https://doi.org/10.1186/1471-2474-4-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2474-4-8