
Abstract
Background
In the annulus, collagen fibers that make up the lamellae have a wavy, planar crimped pattern. This crimping plays a role in disc biomechanical function by allowing collagen fibers to stretch during compression. The relationship between morphologic changes in the aging/degenerating disc and collagen crimping have not been explored.
Methods
Ultrastructural studies were performed on annulus tissue from 29 control (normal) donors (aged newborn to 79 years) and surgical specimens from 49 patients (aged 16 to 77 years). Light microscopy and specialized image analysis to visualize crimping was performed on additional control and surgical specimens. Human intervertebral disc tissue from the annulus was obtained in a prospective morphologic study of the annulus. Studies were approved by the authors' Human Subjects Institutional Review Board.
Results
Three types of morphologic changes were found to alter the crimping morphology of collagen: 1) encircling layers of unusual matrix disrupted the lamellar collagen architecture; 2) collagen fibers were reduced in amount, and 3) collagen was absent in regions with focal matrix loss.
Conclusions
Although proteoglycan loss is well recognized as playing a role in the decreased shock absorber function of the aging/degenerating disc, collagen changes have received little attention. This study suggests that important stretch responses of collagen made possible by collagen crimping may be markedly altered by morphologic changes during aging/degeneration and may contribute to the early tissue changes involved in annular tears.
Similar content being viewed by others


Background
Collagen in the intervertebral disc, tendons, ligaments, heart valve leaflets, intestine and cornea displays a wavy collagen pattern termed "crimping". This crimp morphology contributes an important component to the mechanical responsiveness of these tissues [1–4]. When these tissues experience loading, the collagen crimp architecture is gradually straightened; this makes possible the slight elongation of collagen at the loaded site.
The biomechanical behavior of the disc is closely linked to this specialized crimp morphology (Figure 1) [2, 3]. The collagen fibers which form the annulus possess the same planar crimped geometry as is seen in the tendon. In the disc, however, crimp parameters vary according to the radial distance through the annulus. The lamellar structure of the annulus consists of continuous layers of collagen fibers that encircle the nucleus pulposus. In successive lamellar layers, the fiber orientation changes with respect to the spinal column axis. From the outer annulus inward, the interlamellar angle decreases linearly. Within each lamellar bundle, the collagen fibers are aligned in parallel arrays, and the fibers display a planar crimped waveform with all fibers in register. A gradient of crimp angle also is present in the disc, with crimp angle increasing and the crimp period decreasing from the periphery of the disc inward.

Diagrammatic illustration of the organization of the intervertebral disc with attention to lamellar structure in the annulus and crimping in the collagen fibers. (Reproduced with permission [3].
This complex and sophisticated architectural organization of lamellar disc structure serves to meet the unique biomechanical needs of the healthy disc. Although it is an important aspect of disc structure and function, this topic, which addresses the interface of microscopic anatomy and biomechanical function, has not been studied with specific reference to changes in the disc architecture during aging and degeneration. In the present report we suggest that the important stretch response of collagen made possible by collagen crimping is altered by morphologic changes in disc architecture during aging and degeneration. This alteration in the crimping behavior of collagen may thus contribute to the underlying tissue changes which culminate in fissures and annual tears in the degenerating disc.
Methods
Control (normal) donor disc specimens were obtained from disc surgeries and the NCI Cooperative Human Tissue Network (CHTN). Studies were approved by the human subjects Institutional Review Board. Disc tissue studied here was from the annulus. Patient specimens were obtained from surgical procedures performed on individuals with herniated discs. Electron microscopy was used in the study of a total of 78 subjects (29 were from control donors [aged newborn to 79 years], and 49 were from surgical specimens from patients [aged 16 to 77 years] undergoing disc surgery). Light microscopic studies utilized the annulus from 40 control donors and 53 surgical specimens. The majority of the specimens studied here were from anterior sites of lumbar discs.
Surgical specimens were transported to the lab within 30 minutes of surgical removal in sterile modified minimal essential medium with Earle's salts (MEM, Gibco, Grand Island, NY) with 1% (v/v) L-glutamine (Irvine Scientific, Santa Ana, CA), 1% (v/v) nonessential amino acids (Irvine Scientific), and 1% (v/v) penicillin-streptomycin (Irvine Scientific). CHTN specimens were shipped overnight in sterile MEM. Upon receipt into the lab, specimens from control donors were carefully dissected to obtain specimens from the outer annulus, inner annulus and nucleus.
For electron microscopy, specimens were minced and fixed in Karnovsky's fixative, post-fixed with osmium tetroxide supplemented with 0.1% ruthenium red to enhance proteoglycan visualization in the extracellular matrix [5, 6], embedded in Spurr resin, thin sectioned and grid stained with uranyl acetate and lead citrate. In order to obtain representative specimens to examine with electron microscopy, 4–6 blocks of tissue were trimmed, thick-sectioned and examined with light microscopy and 1–2 blocks (in which the plane of sectioning was parallel to the long axis of the collagen fibrils) selected for thin sectioning. Sections were viewed on a Phillips CM10 transmission electron microscope.
For light microscopy, specimens were fixed in 10% neutral buffered formalin, embedded in paraffin or glycol methacrylate, and stained with picrosirius red-alcian blue, Masson trichrome, or Goldner's stain. Three-dimensional intensity graphical presentation were obtained using BioScan® OPTIMAS™ (BioScan, Edmonds, WA). Morphologic images were captured with a CoolCam 2000 video system and frame grabber (Cool Camera Company, Decatur, GA) and prints were prepared with a Sony Mavigraph color video printer (Sony Electronics, San Jose, CA).
Results
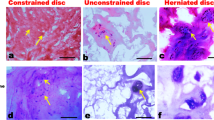
Opportune sectioning planes through the annulus reveal collagen crimping at both the ultrastructural and light microscopic level. Figure 2A illustrates the normal annular morphology of a healthy young intervertebral disc. Disc cells are flattened and lie in a modest lacunar space within collagen lamellar bundles. Prominent wavy expanses of collagen (collagen crimping) are present. Aging and degenerating disc matrix, however, shows striking alterations in tissue architecture. Changes include the presence of layers of unusual extracellular matrix encircling disc cells and interterritorial matrix alterations which extend into the surrounding lamellar structure (Figure 2B). In these regions collagen fibers are fewer in number and fibers are separated from each other reducing collagen crimping areas. Sites of more advanced degeneration can contain areas where there is prominent focal loss of matrix (Figure 2C). Only sparse regions of collagen remain in such sites.

Figs. 2A and 2B: Photomicrographs of light microscopic features of the annulus in a disc from a three month old infant (A) and a control disc from a 68 year old female (B and C). Arrows in A mark regions of prominent collagen crimping. In B note the presence of abnormal matrix encircling disc cells and decreased matrix in intraterritorial regions. In Fig. 2C a region of prominent focal loss of matrix is present near encapsulated cells. (A, glycol methacrylate embedment, Goldner's stain; B and C, paraffin with Masson-trichrome; X 980).
At the ultrastructural level, wavy collagen bands characterize regions of good matrix (Figure 3A). In more degenerated areas, collagen fiber number is much less (Figure 3B).

When viewed with polarizing light microscopy, regions of the disc with collagen crimping stand out prominently, and visualization of crimping patterns can be enhanced by application of specialized image analysis techniques which provide a graphical representation of image intensities (Figure 4). In regions of good disc morphology, collagen crimping patterns produce image intensity graphs where crimp patterns show clear peaks and valleys in register (Figure 4A and 4B). In areas where degenerative changes in the matrix are prominent, however, collagen crimping is markedly disrupted (Figure 4D). Note that the area examined in Figure 4D shows some crimp morphology (arrows), thus confirming that the plane of sectioning was appropriate for the detection of collagen crimping. Figure 4C shows that rare collagen bundles in register with each other are still present in this area, thus verifying that the tissue section plane was appropriate for detection of collagen crimping.

Photomicrographs of disc tissue from the annulus in a region of healthy matrix (Fig. 4B, surgical specimen from a 46 year old female) and from a region with marked matrix changes (Fig. 4D, surgical specimen from a 54 year old male). In Figures 4A and 4C, collagen crimping patterns throughout the same microscopic fields shown in Figs. 4B and 4D are converted to graphic images with three-dimensional intensity images using BioScan® OPTIMAS™ using the following settings for both images: rotation: x, 25.000; y, 1.000; sampling, x: 45, y:45, and viewpoint Z: 3000.0 . White arrows in Fig. 4D mark small regions showing collagen crimping, but degenerative changes in the tissue disrupt crimping in the adjacent regions. (Figs. 4B and 4D, polarized light, picrosirius red/alcian blue, X 102).
Discussion
In order for the disc to successfully withstand mechanical forces, its matrix must have the appropriate structural features, and this matrix must be maintained by appropriate turnover of its molecules. Previous studies from our laboratory have identified similar matrix changes during disc aging and degeneration in disc tissue from both control subjects and patients undergoing disc surgery [7, 8]. Observations presented here focus upon secondary collagen changes involving crimping morphology which nonetheless have important consequences for tissue biomechanical function.
During aging and degeneration of the intervertebral disc, disruption of annular fibers during stress can result in fissures which may extend out to the disc margin [9]. Such radial tears may be a source of the nonspecific early spinal symptoms in disc degeneration [9]. Although the importance of annular tears has long been recognized, the basis for these tears at the architectural level has not been investigated.
In the present work, study of a collection of disc specimens from control and surgical patients has shown that the architectural features of collagen crimping in the annulus can be disrupted in three fashions. First, a common morphologic change during disc aging/degeneration is the accumulation of wide layers of unusual matrix which encircle disc cells and clusters of disc cells [7, 8]. These regions disrupt normal collagen fibrillar layers and thus reduce the functional stretch afforded by collagen crimping in regions of normal collagen. Secondly, in areas where the matrix has become sparse, only a few collagen fibers are present. Such regions would have diminished capability of affording appropriate stretch function to resist tearing under compression. Finally, marked extracellular matrix loss in focal areas altered normal lamellar collagen architecture. The presence of similar types of changes in both control and surgical disc tissue is not surprising in light of previous studies by Boden et al [10, 11] in which degenerative changes were identified with MRI in lumbar and cervical discs of older asymptomatic subjects.
In other ultrastructural studies of disc tissue, we have reported considerable variation in cross-sectional diameters of collagen fibrils in both territorial and inter-territorial extracellular matrix [8]. Cross-sectional diameter variability was present in 34% of control and 59% of surgical specimens which were examined. Such variations may also have a possible effect on the ability of the disc to respond to biomechanical stress.
A limitation of the present study is the qualitative nature of our study. Future studies would be greatly strengthened by quantitative analysis of variations of crimping periodicity in both surgical specimens and in representative sections of complete discs. Such quantitative studies are important since histologic/ultrastructural examination only focuses upon a small portion of tissue. Additional important information lacking from our study is determination of the extent to which the changes described here actually alter the biomechanical properties of the disc. We hope that our small study here will lead to more complete studies of this important topic since there is much to be learned about how disc degeneration relates to increased risk of herniation and pain.
Little is understood about collagen turnover in either the healthy or diseased intervertebral disc. Studies by Antoniou et al have pointed out that it is very difficult to distinguish between synthesis of new matrix, degradation products, and resident matrix collagen [12]. It has been suggested that the turnover in the disc may be in excess of 100 years [9]. Newer methods used by Antoniou et al reveal that synthesis of Type I and II collagen and of aggrecan all decrease from age 2 to 15, but Type I collagen appears to decline more rapidly and to plateau during ages 15–40 years, whereas Type II collagen and aggrecan continue to decline during this period [12]. At older ages (60–80 years), there is increased denaturation of Type II collagen and a small increase in the synthesis of Type I procollagen. The maintenance of appropriate matrix composition in the disc is complicated by a marked decline in the number of disc cells with aging and an increase in cellular death by apoptosis [7].
Conclusions
In summary, use of our large disc tissue collection was an advantage in the present study since it provided examination of numerous disc specimens sampled through various planes of sectioning. Application of image intensity graphs provided objective tracings of collagen patterns. Although proteoglycan loss has long been acknowledged to be an important part of disc aging and degeneration, the work presented here suggests that important stretch responses of collagen made possible by collagen crimping may be markedly altered by morphologic changes in disc architecture during aging/degeneration; such changes may contribute to the early tissue changes involved in annular tears. This work points to the need to more fully understand the dynamic relationship between degenerative changes in disc architecture at the microscopic level and resultant alterations in biomechanical function.
References
Berkovitz B: Collagen crimping in the intra-articular disc and articular surfaces of the human temporomandibular joint. Arch Oral Biol. 2000, 45: 749-756. 10.1016/S0003-9969(00)00045-5.
Cassidy J, Hiltner A, Baer E: Hierarchical structure of the intervertebral disc. Conn. Tiss. Res. 1989, 23: 75-88.
Cassidy J, Hiltner A, Baer E: The response of the hierarchical structure of the intervertebral disc to uniaxial compression. J Materials Science: Materials in Medicine. 1990, 1: 69-80.
Gathercole L, Keller A: Crimp morphology in the fibre-forming collagens. Matrix. 1991, 11: 214-234.
Hunziker E, Ludi A, Herrmann W: Preservation of cartilage matrix proteoglycans using cationic dyes chemically related to ruthenium hexaammine trichloride. J Histochemistry Cytochemistry. 1992, 40: 909-917.
Nuehring L, Steffens W, Rowland G: Comparison of the ruthenium hexammine trichloride method to other methods of chemical fixation for preservation of avian physeal cartilage. Histochem. J. 1991, 23: 201-214.
Gruber H, Hanley E: Analysis of aging and degeneration of the human intervertebral disc – Comparison of surgical specimens with normal controls. Spine. 1998, 23: 751-757. 10.1097/00007632-199804010-00001.
Gruber H, Hanley E: Ultrastructure of the human intervertebral disc during aging and degeneration: Comparison of surgical specimens with controls. Spine.
Fraser F, Bleasel J, Moskowitz R: Spinal degeneration: Pathogenesis and Medical Management. In: The Adult Spine. Principles and Practices. Edited by: Ducker TB, Hadler NM, Kostuik JP, Weinstein JN, Whitecloud TSI. 1997, Philadelphia, Lippincott-Raven, 735-759. Second
Boden SD, Davis DO, Dina TS, Patronas NJ, Wiesel SE: Abnormal magnetic-resonance scans of the lumbar spine in asymptomatic subjects: A prospective investigation. J. Bone Joint Surg. 1990, 72A: 403-408.
Boden SD, McCowin PR, Davis DO, Dina TS, Mark AS, Wiesel S: Abnormal magnetic-resonance scans of the cervical spine in asymptomatic subjects. A prospective investigation. J Bone Joint Surg. 1990, 72A: 1178-1184.
Antoniou J, Steffen T, Nelson F, et al: The human lumbar intervertebral disc – Evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J. Clin. Invest. 1996, 96: 996-1003.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2474/3/9/prepub
Acknowledgements
The authors thank Jane Ingram and Tracie McClain for assistance with light microscopy, and Daisy Ridings, Pat McCoy and Winston Wiggins for assistance with electron microscopy.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
ENH contributed disc tissue from surgeries on patients, and participated in the study and reviewed findings with HEG. HEG conceived of the study and participated in its design, coordination, reviewed all light and electron microscopic material, and wrote the manuscript. All authors read and approved the final manuscript
Competing interests
None declared
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Gruber, H.E., Hanley, E.N. Observations on morphologic changes in the aging and degenerating human disc: Secondary collagen alterations. BMC Musculoskelet Disord 3, 9 (2002). https://doi.org/10.1186/1471-2474-3-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2474-3-9