
Abstract
Background
We earlier showed that 4-phenylbutyrate (PB) can induce cathelicidin LL-37 expression synergistically with 1,25-dihydroxyvitamin D3 in a lung epithelial cell line. We aimed to evaluate a therapeutic dose of PB alone or in combination with vitamin D3 for induction of LL-37 expression in immune cells and enhancement of antimycobacterial activity in monocyte-derived macrophages (MDM).
Methods
Healthy volunteers were enrolled in an 8-days open trial with three doses of PB [250 mg (Group-I), 500 mg (Group-II) or 1000 mg (Group-III)] twice daily (b.d.) together with vitamin D3 {5000 IU once daily (o.d.)}, PB (500 mg b.d.) (Group-IV) or vitamin D3 (5000 IU o.d.) (Group-V), given orally for 4 days. Blood was collected on day-0, day-4 and day-8; plasma was separated, peripheral blood mononuclear cells (PBMC), non-adherent lymphocytes (NAL) and MDM were cultured. LL-37 transcript in cells and peptide concentrations in supernatant were determined by qPCR and ELISA, respectively. In plasma, 25-hydorxyvitamin D3 levels were determined by ELISA. MDM-mediated killing of Mycobacterium tuberculosis (Mtb) (H37Rv) was performed by conventional culture method.
Results
MDM from Group-II had increased concentration of LL-37 peptide and transcript at day-4, while Group-I showed increased transcript at day-4 and day-8 compared to day-0 (p < 0.05). Both Group-I and -II exhibited higher levels of transcript on day-4 compared to Group-III and Group-V (p < 0.035). Increased induction of peptide was observed in lymphocytes from Group-II on day-4 compared to Group-I and Group-IV (p < 0.05), while Group-IV showed increased levels on day-8 compared to Group-I and Group-III (p < 0.04). Intracellular killing of Mtb on day-4 was significantly increased compared to day-0 in Group-I, -II and -V (p < 0.05).
Conclusion
The results demonstrate that 500 mg b.d. PB with 5000 IU o.d. vitamin D3 is the optimal dose for the induction of LL-37 in macrophages and lymphocytes and intracellular killing of Mtb by macrophages. Hence, this dose has potential application in the treatment of TB and is now being used in a clinical trial of adults with active pulmonary TB (NCT01580007).
Similar content being viewed by others


Background
Tuberculosis (TB), caused by Mycobacterium tuberculosis (Mtb), is a predominant public health problem worldwide and responsible for about 3 million deaths annually [1]. The prevalence of TB is increasing due to the spread of antibiotic-resistant strains of Mtb[2, 3]. The deleterious consequences of co-infection with HIV is the emergence of XDR (extremely drug resistance) cases in Africa and other countries in recent years [4]. Advances in anti-tuberculous therapies and alternative treatment strategies are urgently required for the treatment of TB patients and exposed individuals at high risk of developing TB [5, 6].
Antimicrobial peptides (AMPs) are important effectors of the innate defense system [7]. AMPs can limit growth and virulence properties of microbes directly or indirectly by enhancing the host immune system. Induction of endogenous AMP expression by administration of extrinsic compounds may be an attractive approach for alternative therapy in combating infectious diseases. Cathelicidins [8] and Defensins [9] and are major classes of AMPs, in mammals LL-37 is the only human cathelicidin and is expressed by both circulating white blood cells and epithelial surfaces including lungs [10].
The active form of vitamin D3 was shown to regulate the production of AMPs, i.e. cathelicidin LL-37, which plays an important role in the innate immune defense against infections including TB [11, 12]. These studies have encouraged many investigators to initiate vitamin D trials in TB patients with renewed enthusiasm [5, 13–15] (http://NCT01130311; http://NCT00507000; http://NCT00366470; http://NCT00677339; http://NCT00918086). In a systemic review and meta-analysis of observational studies, evidence of an association with vitamin D3 deficiency and active TB was demonstrated [16].
Our group has earlier shown that cathelicidin is downregulated in the mucosal epithelia during acute diarrhea [17, 18] and in infection with Neisseria gonorrhea[19]. We showed that the downregulation of the rabbit cathelicidin (CAP-18) in the large intestine can be counteracted by oral treatment with sodium butyrate, a short chain fatty acid and phenylbutyrate (PB), an analogue of butyrate in experimental model of shigellosis [17, 20]. PB further counteracted the downregulation of CAP-18 in the lung and tracheal epithelia through systemic effects [20]. We further demonstrated that PB can induce LL-37 expression synergistically with 1,25-dihydroxyvitamin D3 at both protein and mRNA levels in a lung epithelial cell line [21]. It is thus likely that oral supplementation with PB will boost innate immunity in the lung mucosa by increasing expression of innate defense proteins. A dose of PB corresponding to that used for rabbits [20] for induction of cathelicidin was calculated by allometric scaling for use in adults. The calculations suggested that an effective dose for a 60 kg human would be approximately 600 mg of PB twice daily.
Our aim in this study was to determine the optimal dose of PB with or without vitamin D3 in adults for induction of LL-37 in immune cells and increase in functional capacity of macrophages in killing of Mtb. Thus, healthy adult volunteers were treated with different doses of PB and in combination with a defined concentration of vitamin D3.
Methods
Study design
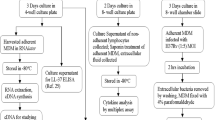
Healthy participants (age range 18 to 55 yrs; n = 15) working at International Centre for Diarrheal Disease Research, Bangladesh (icddr,b) and National Institute of the Diseases of the Chest and Hospital (NIDCH), Dhaka, Bangladesh were recruited in this study. The study was approved by the Ethical Review Committee of icddr,b. Informed consent was obtained from the participants after explaining the nature and purpose of the study. Individuals were excluded if they had symptoms or clinical signs of infection, e.g. fever or diarrhea within the last 2–3 months, if taking corticosteroids, diuretics, or supplementary vitamin D3 (either alone or as part of a multivitamin preparation). The volunteers were asked to abstain from taking any vitamins or supplements for at least two weeks prior to and during the study period. There were five groups, each consisting of 3 participants. Groups-I, -II and -III received the following doses respectively: 250 mg, 500 mg or 1 g of PB b.d. plus 5000 IU vitamin D3 o.d. for 4 consecutive days. Group-IV received 500 mg PB b.d. and Group-V received 5000 IU vitamin D3 o.d. for 4 consecutive days. All healthy volunteers were followed for another 4 days after the treatment to monitor possible side effects. Four-phenylbutyrate (Tributyrate) and the placebo tablet were obtained from Fyrklövern Scandinavia AB, Mönsterås, Sweden; vitamin D3 (Vigantol oil) and the placebo oil were obtained from Merck KGaA, Darmstadt, Germany. The healthy volunteer trial was conducted to determine an optimal dose for the clinical trial in TB patients (http://NCT01580007).
Blood collection and cell culture
Peripheral blood was collected at 3 time points (day-0, day-4 and day-8) from each participant (25 mL/individual). Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood by Ficoll-Hypaque density gradient centrifugation and plasma was collected and stored at −20°C for later measurement of 25-hydroxyvitamin D3, calcium, (serum glutamate pyruvate transaminase (SGPT; liver function marker) and creatinine (kidney function marker). The PBMC pellet was washed and suspended in a culture medium containing 10% autologous plasma in RPMI-1640, 1% L-glutamine, 1% sodium pyruvate, 0.5% amphotericin B and 1% penicillin-streptomycin (Gibco, Grand Island, NY, USA) and plated in two parallel 4-well tissue culture plates (NUNC, Roskilde, Denmark). One tissue culture plate was utilized for analyses of LL-37 peptide and transcript and the other plate was used for macrophage-mediated Mtb killing assay.
PBMCs were counted and incubated in culture plates for 3 days. Thereafter, supernatant containing the non-adherent cells (mostly lymphocytes, 80-90%) was removed, centrifuged to collect the clear supernatant which was the extracellular fluid (ECF) of PBMC. The non-adherent lymphocytes were treated with saponin for 10 minutes, centrifuged and supernatant collected as intracellular fluid (ICF) from lymphocytes. Supernatant and cell pellet were stored for further analysis. The remaining adherent cells in the culture plate were MDM, which were harvested and treated with saponin. After centrifugation, supernatant (ICF) and cells were stored until further analysis. RNALater (Qiagen GmbH, Hilden, Germany) was added to the cell pellet for RNA extraction. In another experiment, one part of PBMC was stimulated ex vivo with Bacille Calmette-Guerin, (BCG, 10 μg/mL: Japan BCG Laboratory, Tokyo, Japan) for 3 days, supernatant containing the non-adherent cells was removed, centrifuged to collect the clear supernatant which was the ECF of BCG stimulated PBMC.
Vitamin D3, calcium, albumin, SGPT and creatinine in plasma
In plasma, 25-hydroxyvitamin D3 was measured by a commercial ELISA kit (IDS, Fountain Hills, Arizona, USA) that determines 25-hydroxyvitamin D3 (100%), 25-hydroxyvitamin D2 (75%) and 24,25-dihydroxyvitamin D3 (≥100%). Calcium, albumin and creatinine were assessed by Roche automated clinical chemistry analyzer, Hitachi 902. Serum glutamate-pyruvate transaminase (SGPT) was measured by Beckman-Coulter AU680 (Japan).
LL-37 ELISA
LL-37 levels in ICF of macrophage and lymphocyte lysates as well as in ECF of PBMC and BCG stimulated PBMC were measured by ELISA. Duplicate samples were tested and concentrations were calculated using a standard curve generated from synthetic LL-37 (Innovagen, Lund, Sweden). Brief procedure of ELISA is as follow. Polystyrene microtiter plates (Maxisorp by NUNC, Naperville, IL, USA) were coated with monoclonal anti-LL-37 [22] (5 μg/ml) in carbonate buffer (15 mM sodium carbonate, 35 mM sodium bicarbonate and 0.02% sodium azide [pH 9.6]) and incubated overnight at 4°C. After washing, non-specific binding was blocked with 0.1% gelatin in tris-buffered saline (pH 7.5) for 1 hour at RT. Standards and samples were then added and incubated overnight at 4°C. Biotinylated rabbit anti-LL-37 (1 μg/mL) (Innovagen) was added and incubated for 2 h at room temperature (RT), followed by the incubation with Streptavidin-alkaline phosphatase conjugate (Chemicon, Melbourne, Australia) for another 2 h at RT. Four-methylumbelliferyl phosphate was used as substrate (Molecular Probes, Europe BV, Leiden, The Netherlands) and fluorescence was measured at an excitation wavelength of 360 nm and emission wavelength of 450 nm.
Quantitative real time RT-PCR amplification of LL-37 mRNA
RNA was extracted from macrophages and lymphocytes utilizing RNeasy Mini kit as described by the manufacturer (Qiagen GmbH). cDNA was synthesized using Superscript III First-Strand Synthesis System (Invitrogen, Grand Island, NY, USA). The CAMP gene encoding transcript LL-37 relative to the housekeeping 18S rRNA was measured in triplicate from the cDNA samples by real-time quantitative RT-PCR using CFX96 Real-Time PCR Detection Systems (Bio-Rad,) and the 18 s rRNA–housekeeping gene kit (Applied Biosystems, Foster City, CA, USA). The sequences of forward and reverse primers for LL-37 transcript were 5’-TCACCAGAGGATTGTGACTTCAA-3’ and 5’-TGAGGGTCACTGTCCCCATAC-3’, respectively (Primer Express; Applied Biosystems). The results were analyzed by using the relative standard method [19].
Macrophage mediated killing of Mycobacterium tuberculosis
MDM in the culture plates were infected with previously prepared log phased Mtb H37Rv strain (from Tuberculosis Research Center, Chennai, India) in culture medium without antibiotics [23]. Initially 3 different multiplicity of infection (MOI; 1:10; 1:25 and 1:50) were tested and the ratio of 1:25 was found to be the optimal MOI, resulting in a clear bacterial killing. After 2 hours of exposure the culture plates were washed 3 times with warm RPMI to remove the extracellular bacteria and the infected MDM were cultured for three additional days in a medium with autologous plasma and antibiotics (penicillin-streptomycin, amphotericin B) (Gibco). Thereafter, the cells were lysed with 0.3% saponin-PBS and vigorous pipetting to collect viable intracellular Mtb. The lysates were cultured on Middle Brook 7H11 agar medium supplemented with 10% Middle Brook OADC (oleic acid, albumin, dextrose and catalase) enrichment (Becton Dickinson, Sparks, MD, USA). Bacterial viability was calculated by counting colony forming units (CFU) on agar plates after 21–28 days of culture at 37°C. Before initiation of any intervention, the day-0 MDM-mediated killing served as the control for all groups.
Statistical analysis
Statistical analyses were performed using SigmaStat 3.1 for Windows (Systat Software Inc., Point Richmond, CA, USA) and SPSS 17.0 for Windows (SPSS Inc, Chicago, Illinois, USA). Results were expressed as mean with standard deviation. Sex and age was matched for each group. Data distribution patterns were checked by using scatter plots, and normality and homogeneity of variances were checked by descriptive statistics. Two-way repeated measure ANOVA was performed when both the sphericity and normality of the data was met. When condition for sphericity was violated or normality of data failed, one Way Analysis of Variance (ANOVA) was performed followed by the Tukey multiple comparison test for between groups and within group variables. Kruskal-Wallis ANOVA on Ranks was performed when data was not normally distributed. Analysis of Co-variance was performed when data at entry level was significantly different. P-values < 0.05 were considered significant.
Results
Plasma levels of vitamin D3and calcium
The average of 25-hydroxyvitamin D3 levels in plasma increased in all groups except for Group-IV (treated with only PB). However, the increase was not significant (Figure 1). Sex and age was matched for each group. Similarly, no changes were observed in albumin-adjusted calcium levels during the study period in any of the groups (data not shown).

Plasma 25-hydroxyvitamin D 3 concentration in healthy adults before and after supplementation. Group-I: 250 mg PB b.d. + 5000 IU vitamin D3 o.d.; Group-II: 500 mg PB b.d. + 5000 IU vitamin D3 o.d.; Group-III: 1000 mg PB b.d. + 5000 IU vitamin D3 o.d.; Group-IV: 500 mg PB b.d.; Group-V: 5000 IU vitamin D3 o.d. PB: Phenylbutyrate. b.d.: twice daily. o.d. : once daily. Data were analyzed by One-way ANOVA.
SGPT level before treatment was normal in all healthy volunteers except for one male volunteer in Group-I, who had 66.49 IU/L that was above the normal upper range of 56 IU/L. However, the SGPT level (12.08 IU/L) declined on day-4 to normal level. None of the volunteers showed any increase in SGPT level after ingestion of PB alone or in combination with vitamin D3 on day-4 or −8 (Additional file 1: Table S1).
Synergistic effects of PB and vitamin D3on LL-37 expression in macrophages
Significant differences in concentration of MDM derived LL-37 transcripts and peptides were observed within days. A significant increase in both peptide (p = 0.05) and transcript (p = 0.011) was noted in Group-II in day-4 compared to day-0 (Table 1 and 2). Group-I showed significantly higher levels of LL-37 transcript but not peptide at day-4 (p = 0.015) and day-8 (p = 0.042) compared to day-0. Group-IV showed a marginal increase in LL-37 transcript levels on day-4 compared to day-0 (p = 0.07) (Table 1).
When comparing between groups, Group-I and Group-II exhibited significantly higher levels of LL-37 transcript on day-4 compared to Group-III (p = 0.025 and p = 0.03 respectively) and Group-V (p = 0.03 and p = 0.035 respectively) (Table 1). Furthermore, Group-II showed higher levels of peptide compared to Group-IV (p = 0.01) (Table 2).
PB and vitamin D3induce LL-37 expression in lymphocytes
Lymphocyte-derived LL-37 transcript did not show any significant induction within days (Table 3). However, LL-37 peptide induction on day-4 was significantly higher in Group-II compared to Group-I (p = 0.03) and Group-IV (p = 0.05) (Table 4). Group-IV showed increased production of peptide on day-8 compared to Group-III (p = 0.03) and Group-I (p = 0.04).
No effects of PB and vitamin D3on LL-37 expression in PBMC
There was no significant induction of LL-37 production in ECF of total PBMC within days or between groups. Stimulation of PBMC with live BCG also did not show any remarkable increase in LL-37 release in the ECF (0.53 ± 0.17 ng/106 PBMC) compared to the ECF of unstimulated PBMC (0.50 ± 0.17 ng/106 PBMC) (p = 0.92).
PB and/or vitamin D3 mediate enhanced killing of Mtb ex vivo
Group-II demonstrated significantly higher intracellular killing of Mtb by MDM at day-4 (p = 0.027) compared to day-0. Group-I and -V exhibited a significant increase in intracellular killing on day-4 (p < 0.001 and p = 0.019 respectively) and day-8 (p < 0.001 and p = 0.051 respectively) compared to day-0 (Figure 2). The other Groups did not show any increase in the killing activity of MDM (Figure 2).

Viable Mtb CFU count in Monocyte derived macrophages (MDM). MDM from different group of volunteers were incubated with Mtb H37Rv strain for 2 h, after that extracellular bacteria were removed and cultured for 3 days, cells were lysed and plated for variable colony (CFU) count. Group-I: 250 mg PB b.d. + 5000 IU vitamin D3 o.d.; Group-II: 500 mg PB b.d. + 5000 IU vitamin D3 o.d.; Group-III: 1000 mg PB b.d. + 5000 IU vitamin D3 o.d.; Group-IV: 500 mg PB b.d.; Group-V: 5000 IU vitamin D3 o.d.. PB: Phenylbutyrate. b.d.: twice daily. o.d. : once daily. The straight horizontal line indicates means. Data were analyzed by two-way (treatment and time) repeated measures ANOVA. * p < 0.05, and *** p < 0.001.
Discussion
In this study we demonstrated that oral supplementation of healthy adult volunteers with 500 mg PB b.d. plus 5000 IU vitamin D3 o.d. (Group-II) consistently induced LL-37 in both macrophages and lymphocytes, and also exhibited increased MDM derived intracellular killing of M. tuberculosis.
In an animal model of shigellosis we have previously shown that Shigella infection causes downregulation of the rabbit cathelicidin CAP-18 in the epithelia of rectum, lung and trachea [17, 20]. Oral feeding of butyrate (0.14 mmol/dose) or PB (0.14 mmol/dose) up-regulates CAP-18 expression in the lung and rectal epithelia of these rabbits and reduces shedding of Shigella in stool [20]. We have also demonstrated that PB, induces the CAMP gene expression synergistically with 1,25-dihydroxyvitamin D3 at both protein and mRNA levels in the VA10 lung epithelial cell line [21]. We have now further shown in humans, that oral intake of PB alone or in combination with vitamin D3 can also induce LL-37 in immune cells. In this study, three potential doses of PB were selected for healthy adults. Group I (250 mg b.d. with 5000 IU vitamin D3 o.d.) showed significant increase in LL-37 transcript but not peptide in MDM; no changes in peptide or transcript was noted in NAL. Group-II (500 mg b.d. plus 5000 IU vitamin D3 o.d.) showed increase in both peptide and transcript in MDM and peptide in NAL. In Group-III with the higher dose of PB (1 g b.d.) along with vitamin D3 there was no increase in LL-37 transcript or peptide in cells after PB supplementation. Butyrate is known to inhibit RNA and protein synthesis at high concentrations [24–26]. We may speculate that in a similar fashion PB at high doses is inhibitory to the expression of LL-37 both at transcriptional and translational levels. The dose of Group-II appears superior in inducing both peptide and mRNA concentrations in MDM and peptide in lymphocytes.
Various cell types including T lymphocytes are known to express LL-37 [27, 28]. However, to our knowledge this is the first study to report enhanced in vivo induction of LL-37 in lymphocytes after oral supplementation with PB alone or in combination with vitamin D3. This finding thus underscores the significance of PB therapy in TB infection since T cells play a major role in the host defense against tuberculosis [29].
The present study showed that oral intake of vitamin D3 (Group-V) alone or in combination with PB (Group-I and -II) exhibited a marked increase in intracellular killing of Mtb. Vitamin D3-induced LL-37 expression is an important factor in fighting TB [11, 12]. Killing of Mtb by macrophages is directly correlated with CAMP gene expression encoding LL-37 and plasma levels of 25-hydroxyvitamin D3[12]. Furthermore, it was shown that activation of Toll-Like Receptor 2/1 in human macrophages up-regulated expression of the genes encoding vitamin D receptor and vitamin D-hydroxylase, leading to induction of LL-37 with subsequent killing of intracellular Mtb[11]. Recent studies have shown that human cathelicidin is a key mediator of 1,25-dihydroxyvitamin D3-induced autophagy and hence provide a mechanistic insight into the role of cathelicidin in combating Mtb[30, 31]. Interestingly, in vitro studies have shown that short chain fatty acids, butyrate and propionate can induce both apoptosis and autophagy [32]. PB is an analogue of butyrate and it is quite likely that PB in combination with Vitamin D3 may also induce autophagy and LL-37 mediated killing of Mtb ex vivo.
The major limitation of this study is the small sample size and the short duration of the PB and vitamin D3 supplementation. The lack of increase in plasma 25-hydroxyvitamin D3 level after 4 days supplementation was not unexpected since 1–2 months supplementation is needed to reach steady state vitamin D3 levels.
Conclusion
In conclusion, oral combination dose of 500 mg PB b.d. with 5000 IU vitamin D3 o.d. appears to be the optimum dose to induce both LL-37 peptide and transcript expression in functional immune cells as well as enhance intracellular Mtb killing in macrophages. The dose of 250 mg PB b.d. with 5000 IU vitamin D3 o.d. also increased intracellular killing however it enhanced only LL-37 transcript levels (not peptide) in macrophages but not in lymphocytes. This pilot study has generated results for a potential dose in a clinical trial of adults with active pulmonary TB (http://NCT01580007).
Ethical approval
The study was approved by the Ethical Review Committee of International Centre for Diarrheal Disease Research, Bangladesh (icddr,b).
Abbreviations
- AMPs:
-
Antimicrobial peptides
- b.d.:
-
Twice daily
- CFU:
-
Colony forming units
- ECF:
-
Extracellular fluid
- HDAC:
-
Histone deacetylase
- ICF:
-
Intracellular fluid
- MDM:
-
Monocyte-derived macrophages
- Mtb:
-
Mycobacterium tuberculosis
- o.d.:
-
Once daily
- PB:
-
4-Phenylbutyrate
- PBMCs:
-
Peripheral blood mononuclear cells
- TB:
-
Tuberculosis.
References
Smith PE, Moss AR: Tuberculosis: pathogenesis, protection, and control, epidemiology of tuberculosis. Epidemiol Tuberc. 1994, 1: 47-60.
Parsons LM, Driscoll JR, Taber HW, Salfinger M: Drug resistance in tuberculosis. Infect Dis Clin North Am. 1997, 11 (4): 905-928. 10.1016/S0891-5520(05)70397-4.
Bradford WZ, Daley CL: Multiple drug-resistant tuberculosis. Infect Dis Clin North Am. 1998, 12 (1): 157-172. 10.1016/S0891-5520(05)70415-3.
Gandhi NR, Shah NS, Andrews JR, Vella V, Moll AP, Scott M, Weissman D, Marra C, Lalloo UG, Friedland GH: HIV coinfection in multidrug- and extensively drug-resistant tuberculosis results in high early mortality. Am J Respir Crit Care Med. 2010, 181 (1): 80-86. 10.1164/rccm.200907-0989OC.
Wejse C, Gomes VF, Rabna P, Gustafson P, Aaby P, Lisse IM, Andersen PL, Glerup H, Sodemann M: Vitamin D as supplementary treatment for tuberculosis: a double-blind, randomized, placebo-controlled trial. Am J Respir Crit Care Med. 2009, 179 (9): 843-850. 10.1164/rccm.200804-567OC.
Martins M: Targeting the human macrophage with combinations of drugs and inhibitors of Ca2+ and K + transport to enhance the killing of intracellular multi-drug resistant Mycobacterium tuberculosis (MDR-TB)–a novel, patentable approach to limit the emergence of XDR-TB. Recent Pat Antiinfect Drug Discov. 2011, 6 (2): 110-117. 10.2174/157489111796064524.
Zasloff M: Antimicrobial peptides of multicellular organisms. Nature. 2002, 415 (6870): 389-395. 10.1038/415389a.
Zaiou M, Gallo RL: Cathelicidins, essential gene-encoded mammalian antibiotics. J Mol Med (Berl). 2002, 80 (9): 549-561. 10.1007/s00109-002-0350-6.
Ganz T: Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol. 2003, 3 (9): 710-720. 10.1038/nri1180.
Kai-Larsen Y, Agerberth B: The role of the multifunctional peptide LL-37 in host defense. Front Biosci. 2008, 13: 3760-3767.
Liu PT, Stenger S, Li H, Wenzel L, Tan BH, Krutzik SR, Ochoa MT, Schauber J, Wu K, Meinken C: Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science. 2006, 311 (5768): 1770-1773. 10.1126/science.1123933.
Liu PT, Stenger S, Tang DH, Modlin RL: Cutting edge: vitamin D-mediated human antimicrobial activity against Mycobacterium tuberculosis is dependent on the induction of cathelicidin. J Immunol. 2007, 179 (4): 2060-2063.
Salahuddin N, Ali F, Hasan Z, Rao N, Aqeel M, Mahmood F: Vitamin D accelerates clinical recovery from tuberculosis: results of the SUCCINCT study [supplementary cholecalciferol in recovery from tuberculosis]. A randomized, placebo-controlled, clinical trial of vitamin D supplementation in patients with pulmonary tuberculosis. BMC Infect Dis. 2013, 13: 22-10.1186/1471-2334-13-22.
Martineau AR, Wilkinson RJ, Wilkinson KA, Newton SM, Kampmann B, Hall BM, Packe GE, Davidson RN, Eldridge SM, Maunsell ZJ: A single dose of vitamin D enhances immunity to mycobacteria. Am J Respir Crit Care Med. 2007, 176 (2): 208-213. 10.1164/rccm.200701-007OC.
Martineau AR, Timms PM, Bothamley GH, Hanifa Y, Islam K, Claxton AP, Packe GE, Moore-Gillon JC, Darmalingam M, Davidson RN: High-dose vitamin D(3) during intensive-phase antimicrobial treatment of pulmonary tuberculosis: a double-blind randomised controlled trial. Lancet. 2011, 377 (9761): 242-250. 10.1016/S0140-6736(10)61889-2.
Nnoaham KE, Clarke A: Low serum vitamin D levels and tuberculosis: a systematic review and meta-analysis. Int J Epidemiol. 2008, 37 (1): 113-119.
Raqib R, Sarker P, Bergman P, Ara G, Lindh M, Sack DA, Nasirul Islam KM, Gudmundsson GH, Andersson J, Agerberth B: Improved outcome in shigellosis associated with butyrate induction of an endogenous peptide antibiotic. Proc Natl Acad Sci U S A. 2006, 103 (24): 9178-9183. 10.1073/pnas.0602888103.
Islam D, Bandholtz L, Nilsson J, Wigzell H, Christensson B, Agerberth B, Gudmundsson G: Downregulation of bactericidal peptides in enteric infections: a novel immune escape mechanism with bacterial DNA as a potential regulator. Nat Med. 2001, 7 (2): 180-185. 10.1038/84627.
Bergman P, Johansson L, Asp V, Plant L, Gudmundsson GH, Jonsson AB, Agerberth B: Neisseria gonorrhoeae downregulates expression of the human antimicrobial peptide LL-37. Cell Microbiol. 2005, 7 (7): 1009-1017. 10.1111/j.1462-5822.2005.00530.x.
Sarker P, Ahmed S, Tiash S, Rekha RS, Stromberg R, Andersson J, Bergman P, Gudmundsson GH, Agerberth B, Raqib R: Phenylbutyrate counteracts Shigella mediated downregulation of cathelicidin in rabbit lung and intestinal epithelia: a potential therapeutic strategy. PLoS One. 2011, 6 (6): e20637-10.1371/journal.pone.0020637.
Steinmann J, Halldorsson S, Agerberth B, Gudmundsson GH: Phenylbutyrate induces antimicrobial peptide expression. Antimicrob Agents Chemother. 2009, 53 (12): 5127-5133. 10.1128/AAC.00818-09.
Yoshio H, Tollin M, Gudmundsson GH, Lagercrantz H, Jornvall H, Marchini G, Agerberth B: Antimicrobial polypeptides of human vernix caseosa and amniotic fluid: implications for newborn innate defense. Pediatr Res. 2003, 53 (2): 211-216. 10.1203/00006450-200302000-00003.
Edfeldt K, Liu PT, Chun R, Fabri M, Schenk M, Wheelwright M, Keegan C, Krutzik SR, Adams JS, Hewison M: T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin D metabolism. Proc Natl Acad Sci U S A. 2010, 107 (52): 22593-22598. 10.1073/pnas.1011624108.
Soliman MM, Ahmed MM, Salah-Eldin AE, Abdel-Aal AA: Butyrate regulates leptin expression through different signaling pathways in adipocytes. J Vet Sci. 2011, 12 (4): 319-323. 10.4142/jvs.2011.12.4.319.
de Haan JB, Gevers W, Parker MI: Effects of sodium butyrate on the synthesis and methylation of DNA in normal cells and their transformed counterparts. Cancer Res. 1986, 46 (2): 713-716.
Soliman M, Kimura K, Ahmed M, Yamaji D, Matsushita Y, Okamatsu-Ogura Y, Makondo K, Saito M: Inverse regulation of leptin mRNA expression by short- and long-chain fatty acids in cultured bovine adipocytes. Domest Anim Endocrinol. 2007, 33 (4): 400-409. 10.1016/j.domaniend.2006.08.005.
Kin NW, Chen Y, Stefanov EK, Gallo RL, Kearney JF: Cathelin-related antimicrobial peptide differentially regulates T- and B-cell function. Eur J Immunol. 2011, 41 (10): 3006-3016. 10.1002/eji.201141606.
Agerberth B, Charo J, Werr J, Olsson B, Idali F, Lindbom L, Kiessling R, Jornvall H, Wigzell H, Gudmundsson GH: The human antimicrobial and chemotactic peptides LL-37 and alpha-defensins are expressed by specific lymphocyte and monocyte populations. Blood. 2000, 96 (9): 3086-3093.
Ottenhoff TH, Kaufmann SH: Vaccines against tuberculosis: where are we and where do we need to go?. PLoS Pathog. 2012, 8 (5): e1002607-10.1371/journal.ppat.1002607.
Campbell GR, Spector SA: Autophagy induction by vitamin D inhibits both Mycobacterium tuberculosis and human immunodeficiency virus type 1. Autophagy. 2012, 8 (10): 1523-1525.
Yuk JM, Shin DM, Lee HM, Yang CS, Jin HS, Kim KK, Lee ZW, Lee SH, Kim JM, Jo EK: Vitamin D3 induces autophagy in human monocytes/macrophages via cathelicidin. Cell Host Microbe. 2009, 6 (3): 231-243. 10.1016/j.chom.2009.08.004.
Lee JS, Lee GM: Effect of sodium butyrate on autophagy and apoptosis in Chinese hamster ovary cells. Biotechnol Prog. 2012, 28 (2): 349-357. 10.1002/btpr.1512.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2466/13/23/prepub
Acknowledgement
The study was supported by icddr,b, ‘the Swedish Agency for Research Cooperation with Developing Countries (Sida/SAREC Agreement support; grant 384, SWE-2008-065)’; ‘The Swedish Research Council, (grant 58X-11217-14-3)’; ‘The Swedish Strategic Foundation (SSF)’; Swedish Cancer Society; ‘Karolinska Institutet’; ‘The Icelandic Centre for Research (RANNIS)’ and ‘University of Iceland research fund’. icddr,b acknowledges with gratitude the commitment of all donors’ to it’s research efforts. GHG is a visiting scientist supported by ‘The Wenner-Gren Foundations’. We thank the volunteers for participating in this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
Birgitta Agerberth, Gudmundur H. Gudmundsson and Rubhana Raqib have a pending patent application for the use of phenylbutyrate to the treatment of infections. There is no financial arrangement between any of the authors.
Authors’ contributions
RR, BA contributed to the conception and design of the study; AM, RSR, EA and PS performed laboratory analysis, AM, RSR and RR performed the statistical analysis; AM, RSR and RR prepare the first draft of the manuscript. BA, GHG, ZR, SMMK and PS revised the manuscript. All authors read and approved the final version of the manuscript.
Akhirunnesa Mily, Rokeya Sultana Rekha contributed equally to this work.
Electronic supplementary material
12890_2012_394_MOESM1_ESM.pdf
Additional file 1: Table 1: Serum glutamate-pyruvate transaminase and creatinine levels in healthy adults supplemented with phenylbutyrate and vitamin D3 alone or in combination in different doses. (PDF 53 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Mily, A., Rekha, R.S., Kamal, S.M.M. et al. Oral intake of phenylbutyrate with or without vitamin D3upregulates the cathelicidin LL-37 in human macrophages: a dose finding study for treatment of tuberculosis. BMC Pulm Med 13, 23 (2013). https://doi.org/10.1186/1471-2466-13-23
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2466-13-23