
Abstract
Background
Cornelia de Lange syndrome (CdLS) is a rare autosomal-dominant disorder characterised by facial dysmorphism, growth and psychomotor developmental delay and skeletal defects. To date, causative mutations in the NIPBL (cohesin regulator) and SMC1A (cohesin structural subunit) genes account for > 50% and 6% of cases, respectively.
Methods
We recruited 50 patients with a CdLS clinical diagnosis or with features that overlap with CdLS, who were negative for mutations at NIPBL and SMC1A at molecular screening. Chromosomal rearrangements accounting for the clinical diagnosis were screened for using array Comparative Genomic Hybridisation (aCGH).
Results
Four patients were shown to carry imbalances considered to be candidates for having pathogenic roles in their clinical phenotypes: patient 1 had a 4.2 Mb de novo deletion at chromosome 20q11.2-q12; patient 2 had a 4.8 Mb deletion at chromosome 1p36.23-36.22; patient 3 carried an unbalanced translocation, t(7;17), with a 14 Mb duplication of chromosome 17q24.2-25.3 and a 769 Kb deletion at chromosome 7p22.3; patient 4 had an 880 Kb duplication of chromosome 19p13.3, for which his mother, who had a mild phenotype, was also shown to be a mosaic.
Conclusions
Notwithstanding the variability in size and gene content of the rearrangements comprising the four different imbalances, they all map to regions containing genes encoding factors involved in cell cycle progression or genome stability. These functional similarities, also exhibited by the known CdLS genes, may explain the phenotypic overlap between the patients included in this study and CdLS. Our findings point to the complexity of the clinical diagnosis of CdLS and confirm the existence of phenocopies, caused by imbalances affecting multiple genomic regions, comprising 8% of patients included in this study, who did not have mutations at NIPBL and SMC1A. Our results suggests that analysis by aCGH should be recommended for CdLS spectrum cases with an unexplained clinical phenotype and included in the flow chart for diagnosis of cases with a clinical evaluation in the CdLS spectrum.
Similar content being viewed by others

Background
Cornelia de Lange syndrome (CdLS) is a rare, genetically heterogeneous (OMIM #122470, #300590 and #610759), multiple congenital anomaly/intellectual disability disease [1, 2], characterised by distinctive facial dysmorphism, pre- and post-natal growth deficiency, psychomotor delay, intellectual disability and malformations of the upper limbs (ranging from small hands to complete limb reduction). CdLS also often involves specific medical complications such as gastroesophageal reflux, hypoacusia and seizures. Its clinical presentation ranges from mild/borderline to severe [3], and this variability has led to the definition of a list of consensus diagnostic criteria integrated into a global scoring system of phenotype severity [4], which are an accepted standard [5–7].
Known CdLS-associated genes encode structural and regulatory proteins of the cohesin pathway, which is involved in chromosome segregation, DNA repair, gene expression and chromosome conformation [8]. The first major gene to be identified was NIPBL, which is located at chromosome 5p13.2, encodes a member of the adherin family [9, 10], and mutations in this gene are responsible for > 50% of CdLS patients. All types of NIPBL point mutations have been described, although truncating mutations are generally associated with a more severe phenotype than missense and regulatory mutations [5, 9–24]. Microdeletions involving one or more exons of the NIPBL genomic region, and large rearrangements extending to the NIPBL flanking regions, and correlating with severe syndromic presentation, have also been reported [25–27].
Locus heterogeneity in CdLS has been demonstrated by the X-linked form caused by mutation of the SMC1A gene, which encodes a subunit of the cohesin complex [28]. SMC1A alterations contribute up to 6% of all CdLS cases and include only missense mutations or in-frame deletions that preserve the protein reading frame [14, 24, 28–31]. So far, only one patient has been found to have a mutation in the SMC3 gene, which encodes the other SMC cohesin component, and its epidemiological impact has not yet been defined [29]. Very recently, mutations in the HDAC8 gene, a vertebrate SMC3 deacetylase, have been identified in CdLS probands [32], and mutations in RAD21 gene have been found in six patients with CdLS features [33]. The remaining CdLS cases may be due to as yet undetected mutations in the known genes or by other causative anomalies.
The genomic technology of array Comparative Genomic Hybridisation (aCGH), which monitors losses or gains in chromosome regions that may harbour novel candidate genes, is not yet a standard test for investigation of NIPBL- and SMC1A-mutation-negative CdLS patients [11, 18, 34, 35], but results obtained with the technique to date are consistent with those of > 30 conventional cytogenetic and FISH-targeted studies that have shown chromosomal abnormalities associated with the CdLS phenotype involving almost all of the chromosomes (reviewed [36]). One study has used aCGH to study probands with CdLS-like features, who had been previously screened for mutations in the two major causative genes; however, this was performed in a relatively small patient cohort [35].
This aCGH study of 50 probands including patients fulfilling CdLS diagnostic criteria and those not completely fulfil the criteria [4], and negative for mutations at the NIPBL and SMC1A loci, led to the detection of four carriers of large genomic imbalances that are candidates to explain the clinical phenotype and represent a fraction (8%) of patients with features overlapping those of CdLS. We herein describe how the analysis of the gene content of these imbalances, affecting different genomic regions, links the altered dosage of specific gene classes, shared by all rearrangements, to a common CdLS-like phenotype.
Methods
Patients
CdLS is characterized by a wide phenotypic spectrum; despite some features are quite typical the patients present with a highly variable phenotype ranging from severe to very mild. Out of the fifty probands (26 males and 24 females) investigated in this study diagnosis by our clinical geneticists (AS, RT, GZ) was CdLS for those (60%) fulfilling the international CdLS diagnosis criteria [4] or CdlS-like for the remaining (40%,) not fully satisfying the CdLS criteria. According to the CdLS scoring system [4] the overall phenotype of the patients was severe (~10%) or moderate-mild (~90% with slight prevalence of moderate phenotype). All patients were found negative for NIPBL and SMC1A mutations by DHPLC, direct sequencing and MLPA analyses.
Written informed consent to the research study, which was approved by the Ethical Clinical Research Committee of Istituto Auxologico Italiano, and to the publication of the face photo(s) was obtained from one of the parents.
Array-CGH analysis
The probands were investigated by means of aCGH. Genome scans were performed using the Human Genome CGH Microarray Kit 244 K (Agilent Technologies, Palo Alto, CA), which consists of ~236,000 60-mer oligonucleotide probes covering the entire genome at an average spatial resolution of ~30 kb. The samples were labelled and hybridised following the protocols provided by Agilent, and the arrays were analysed using the Agilent Scanner Control (v 7.0) and Feature Extraction software (v 9.5.1). Graphical overviews were obtained using CGH Analytic software (v4.0.81). Aberration calls were identified using the ADM-2 algorithm.
An in silico analysis of the unbalanced regions indicated by aCGH was performed using the March, 2006, release of the UCSC Genome Browser (http://genome.ucsc.edu/) and the Database of Genomic Variants (http://projects.tcag.ca/variation).
FISH
Chromosome preparations were obtained by standard cytogenetic techniques using peripheral blood lymphocytes cultured by 72 h. BAC probes were selected on the basis of their physical location (http://www.genome.ucsc.edu/ release March, 2006), and provided by Invitrogen Ltd., UK. Their physical positions were verified on control metaphase chromosomes derived from peripheral blood lymphocytes. FISH experiments were performed using standard procedures [37].
Results
The 50 probands negative for NIPBL and SMC1A mutations were considered an ideal cohort to scan for the presence of genomic gains/losses by aCGH, in the search for novel genes responsible for phenotypes with features that overlap CdLS.
We identified four probands with large or de novo copy number variants (CNVs) (Table 1), whose clinical data at birth and at age of evaluation are summarised in Table 2 (see also Additional file 1: Table S1 for auxological parameters). The four probands display a high degree of phenotypic heterogeneity, but all share the minimal diagnostic CdLS clinical features. Three of them (Probands 1, 2 and 3) fulfil the diagnostic CdLS criteria, whereas the fourth (Proband 4) does not fulfil completely the criteria having synophrys and only two (and not three) other facies criteria (Additional file 2: Table S2).
Proband 1
The first proband was a 9-year-old girl who showed both intra-uterine (IUGR) and post-natal growth retardation (PNGR) (Table 2 and Additional file 1: Table S1).
Post-natal growth was poor and psychomotor development retarded due to feeding problems (sucking and swallowing difficulties), and surgical correction of a bilateral inguinal hernia was performed at 3 months of age. At the age of 3.5 months, she developed behavioural disorders including hyperactivity, frequent outbursts/temper tantrums and self-injurious behaviour, with self-hitting and self-biting.
When she was 6 years old, a clinical examination revealed generalised hirsutism and dysmorphic features, such as synophrys, long and downward-slanting palpebral fissures, epicanthic folds, long and curved eyelashes, malar hypoplasia, hypoplastic nasal bone, the columella below the alae nasi, a thin upper lip and downturned corners of the mouth and a high-arched palate (Figure 1a). She also had slight limb involvement with bilateral thenar and hypothenar hypoplasia. At the age of 9 years and 10 months, she had developed brachycephaly and her dysmorphic facial features were unchanged. Heart sonography revealed a secundum small atrial septal defect. She no longer presented gastrointestinal problems except for mild rectal bleeding due to multiple juvenile polyps (> 15) throughout the colon.

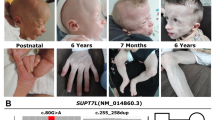
Facial appearance, genomic imbalances and genes with altered copy number in probands. (a, d, g, j) The facial appearance of patients 1 (age 6 years), 2 (age 15 years), 3 (age 22 years) and 4 (age 9 years). (b, e, h, k) aCGH profiles. (Left) Ideogram of the chromosome(s) involved in the imbalances with the log2 probe ratio plotted as a function of chromosomal position; h) profile of patient 4 (left) and his mother (right). (c, f, i, m) The gene content of each genomic imbalance. Magnification of the deleted/duplicated region indicating the distal and proximal breakpoint positions (horizontal dotted lines) and a selection of gene content. Blue colour indicates genes cited in the main text. The red bar in m) corresponds to the CTD-3009K5 BAC clone used in the FISH analysis (not to scale). l) FISH analysis using the CTD-3009K5 BAC clone mapping to chromosome 19p13.3 shows a duplicated signal in all cells of proband 4 (left) and in approximately 76% of the nuclei in a maternal sample, confirming the presence of the rearrangement in a mosaic state (right). White arrows indicate nuclei with a duplicated signal.
The aCGH analysis showed a large, 4.2 Mb, de novo deletion of chromosome 20q11.2-q12 (Table 1, Figure 1b) that was not found in her healthy parents (not shown). More than 50 genes are localised to the deleted region (Figure 1c).
Proband 2
Proband 2 was a 22-year-old woman who had been affected by both IUGR and PNGR (Table 2 and Additional file 1: Table S1). Clinical evaluation revealed slight hirsutism and dysmorphic features, including microcephaly with temporal narrowing, synophrys, long eyelashes, a large nasal tip, anteverted nares, a long philtrum, a large mouth, a thin upper lip and micrognathia (Figure 1d). Limb involvement was mild, marked by small hands, a proximally placed thumb and slight toenail dysplasia. The patient suffered from gastroesophageal reflux, and mitral valve prolapse.
Neurological assessment showed severe psychomotor and intellectual disability, with some behavioural disorders such as hyperactivity.
The aCGH analysis revealed the presence of a ~4.8 Mb interstitial deletion of chromosome 1p36.23-36.22 1 (Table 1, Figure 1e). Her healthy parents have a normal molecular karyotype (not shown), indicating the de novo origin of the rearrangement. The large deleted region includes > 50 genes and does not overlap with the 1p36 syndrome regions (Figure 1f).
Proband 3
In the first years of life, proband 3, a 22-year-old man, suffered from cryptorchidism and feeding problems with swallowing difficulties, as well as post-natal growth delay and cognitive impairment, with both psychomotor and intellectual disability (Table 2 and Additional file 1: Table S1).
His current phenotype is characterised by hirsutism and facial features consisting of microbrachycephaly, a long face, low-set ears, synophrys, thick eyebrows, long eyelashes, a depressed nasal bridge, a large columella, a long and prominent philtrum, a thin upper lip, a high palate and downturned corners of the mouth (Figure 1g). Limb involvement includes post-axial polydactily of the left hand and foot.
The proband is a carrier of an unbalanced 46,XY translocation der(7)t(7;17)(p22.3;q24.2), inherited from his father, who carries a t(7;17)(p22.3;q24.2) balanced translocation. The aCGH analysis performed to characterise the double segmental imbalances revealed a 14 Mb duplication at chromosome 17q24.2-25.3 and a 769 kb deletion of 7p22.3 (Table 1, Figure 1h). Both affected genomic intervals include a number of genes (Figure 1i).
Proband 4
This 10-year-old boy, who had presented prenatally with IUGR, was found at clinical evaluation to have dysmorphic facial features including synophrys, a long philtrum, a thin upper lip and ear lobe creases (Figure 1l), along with musculoskeletal anomalies (clynodactyly of the fifth finger and scoliosis); other features were polythelia and cryptorchidism (Table 2 and Additional file 1: Table S1). He also had psychological and cognitive disorders, including hyperactivity, emotional problems and mild intellectual disability. The mother of the proband had similar facial features (synophrys, a long hypoplastic philtrum, a thin upper lip and ear lobe creases), and a monolateral hypoplasic kidney. She has different neurocognitive and psychological disorders, characterised by dyslexia and bulimia.
The aCGH analyses of the proband and his parents revealed an 880 Kb chromosome 19p13.3 duplication in the son and mother who, on the basis of the signals ratio, appeared to be a mosaic (Table 1, Figure 1m).
To estimate the mosaicism rate with a cell-to-cell-based technique, an interphase FISH experiment was carried out on the mother’s nuclei using the CTD-3009K5 BAC clone, which maps within the duplicated region (Figure 1m). Hybridisation signals were scored on 100 nuclei each from mother and son. A signal of either duplicated or increased intensity was identified in all of the son’s cells, whereas in the sample from the mother, cells containing nuclei with this abnormal pattern were prevalent (76%), but the remaining fraction showed a normal hybridisation pattern, confirming the presence of mosaicism for the mutation (Figure 1n).
Forty-three RefSeq genes are located in the duplicated interval (Figure 1o).
Discussion
CdLS is a genetically heterogeneous disorder, with only 50–60% of clinically diagnosed probands shown to have mutations in one of the known cohesin-associated genes NIPBL, SMC1A, SMC3, HDAC8 or RAD21. This partial knowledge of the molecular basis of CdLS parallels the wide clinical spectrum, which ranges from extremely mild to severe and includes “borderline” cases, which are often at the interface with other syndromic conditions caused by defects in interconnected cohesion pathways [33]. The genes for cohesin structural subunits and regulators perform crucial roles in the maintenance of genome stability through surveillance of chromatid cohesion throughout the cell cycle, double-strand DNA break repair and long-range regulation of transcription (see [38] for a review). While transcriptional activation and regulation occurs only in cycling and postmitotic cells, the more ancient roles of cohesion in adhesion of sister chromatids and DNA repair are performed throughout the cell cycle (see [39] for a review). Filtering atypical/borderline cases from the overall set of patients with a presumptive or possible CdLS clinical diagnosis is a major challenge. The technology used in this study (aCGH) can identify genomic regions implicated in CdLS and overlapping phenotypes by identifying CNVs that may harbour genes encoding the large number of proteins that may interact with those of the cohesin pathway. Here, we describe four probands with a CdLS-like phenotype and without evidence of mutations in NIPBL and SMC1A. The aCGH analyses detected unbalanced rearrangements of various sizes and involving chromosomal regions that have not previously been associated with the Cornelia de Lange phenotypic spectrum.
The four imbalances detected were: i) a de novo deletion at chromosome 20q11.2-q12 (4.2 Mb); ii) a de novo 1p36.23-36.22 deletion (4.8 Mb); iii) a der(7)t(7;17)(p22.3;q24.2) with a 14 Mb duplication in 17q24.2-25.3 and a 769 Kb deletion in 7p22.3; and iv) a familial 880 Kb duplication in 19p13.3, apparently de novo in the mother, who presented with a mosaic state and transmitted the duplication to the affected child.
A number of patients carrying pathogenic deletions or duplications partially overlapping those of our cases have been deposited in DECIPHER or ISCA databases. However, the paucity of the accessory clinical data does not permit conclusive comparisons for genotype phenotype correlations.
To the best of our knowledge, only the chromosome 20q imbalance has previously been described in three patients not classified as having CdLS (although they have some features in common with CdLS) who carry a pure 20q deletion that completely or partially overlaps that identified in our proband 1 (Additional file 3: Figure S1 and Additional file 4: Table S3) [40–42]. No patients have been reported in the literature sharing imbalances in the same regions as the other probands. Four patients have been described bearing a terminal or interstitial deletion of 1p36, with proximal breakpoints falling within the region deleted in our proband 2 (patients A, B, and F in [43], and patient D1P3 in [44]), with whom they share a very small part of the deleted region, consistent with the lack of a common clinical picture. Five patients with a duplicated 19p13.3 region have been described, but all carry a deletion of a different genomic region, thus making them not directly comparable to our proband 4 [45–49].
A large number of the genes located in the regions involved in the imbalances identified in this study have been associated with clinical conditions, making it likely that the phenotypes of our probands are the result of contiguous gene syndromes that mimic the multifaceted CdLS syndrome. Our four probands display a high degree of phenotypic heterogeneity, but all share the minimal diagnostic CdLS clinical features including pre- and post-natal growth retardation (Additional file 1: Table S1), mild to severe psychomotor and cognitive impairment and a cranio-facial appearance (Figure 1) characterised by microcephaly or plagiocephaly, long eyelashes, synophrys, thin upper lip and downturned corners of the mouth, long and prominent philtrum, and hirsutism (Table 2).
It is interesting to note that all of the genomic regions involved in the imbalances described here harbour some dosage-altered genes whose functions are directly or indirectly related to those of the known CdLS genes (NIPBL, SMC1A, SMC3) (Figure 1). The genes of interest (CEP250, DSN1, MAD2L2, APITD1/CENP-S, RECQL5, ANAPC11, MED16, MBD3) encode proteins involved in controlling cell cycle progression, including components of the centrosome, the kinetochore, the mitotic spindle assembly checkpoint, the anaphase-promoting complex, and proteins involved in the methylation and unwinding of DNA (Table 3) [50–63].
In addition to the presence of genes whose products functionally overlap with those of known CdLS genes, further findings supporting the hypothesis that our probands are phenocopies of CdLS include: i) the localisation of the CEP170 gene which encodes a component of the centrosome [64], within a region (chromosome 1q44) shown to be deleted in a CdLS proband by Borck et al. [11]; ii) the localisation of the TNKS gene, involved in sister chromatid cohesion, within the chromosome 8p23.1 region in the CdLS proband reported by Baynam et al. [65].
Moreover, genes with similar functions (RECQL4, BUB1B, BUB3, CENPA, CENPL, SMARCA4, SMARCC1 and ATRX) have also been found to be dysregulated in expression studies of CdLS patients with mutations in NIPBL[66].
Conclusions
We found that a considerable fraction (8%) of the NIPBL and SMC1A mutation-negative probands with features overlapping with CdLS included in this study were carriers of chromosomal imbalances that may underlie their phenotypes.
The four probands had different chromosomal imbalances, but all involved a number of genes related to progression through the cell cycle and the safeguarding of chromosomal stability (Table 3). Given the similar functions of delangin and the proteins of the cohesin network, we hypothesise that an imbalance of these genes, which probably act in concert with other functionally related genes, contributes to the observed CdLS-like phenotypes.
Our data raise the issue of the complex clinical diagnosis of a syndrome such as CdLS which, through the multifunctional proteins encoded by its known causative genes, has an impact on a myriad of interconnected pathways. It is therefore not surprising that the clinical diagnosis of a CdLS-like phenotype often includes cases of carriers of chromosomal imbalances affecting multiple genomic regions.
References
Brachmann W, Ein Fall von symmetrischer Monodaktylie durch Ulnadefekt, mit symmetrischer Flughautbildung in den Ellenbeugen, sowie anderen Abnormitäten: A case of symmetrical monodactyly representing ulnar deficiency, with symmetrical antecubital webbing and other abnormalities (dwarfish, cervical ribs, hirsutism). Jahrbuch Kinderheilkunde und physische Erziehun. 1916, 84: 225-235.
De Lange C: Sur un type nouveau de degenerescence (typus Amstelodamensis). Arch. Med. Enfants. 1933, 36: 713-719.
Van Allen MI, Filippi G, Siegel-Bartelt J, Yong SL, McGillivray B, Zuker RM, Smith CR, Magee JF, Ritchie S, Toi A: Clinical variability within Brachmann-de Lange syndrome: a proposed classification system. Am J Med Genet. 1993, 47: 947-958. 10.1002/ajmg.1320470704.
Kline AD, Krantz ID, Sommer A, Kliewer M, Jackson LG, FitzPatrick DR, Levin AV, Selicorni A: Cornelia de Lange syndrome: clinical review, diagnostic and scoring systems, and anticipatory guidance. Am J Med Genet A. 2007, 143A: 1287-1296. 10.1002/ajmg.a.31757.
Selicorni A, Russo S, Gervasini C, Castronovo P, Milani D, Cavalleri F, Bentivegna A, Masciadri M, Domi A, Divizia MT, Sforzini C, Tarantino E, Memo L, Scarano G, Larizza L: Clinical score of 62 Italian patients with Cornelia de Lange syndrome and correlations with the presence and type of NIPBL mutation. Clin Genet. 2007, 72: 98-108. 10.1111/j.1399-0004.2007.00832.x.
Oliver C, Bedeschi MF, Blagowidow N, Carrico CS, Cereda A, Fitzpatrick DR, Gervasini C, Griffith GM, Kline AD, Marchisio P, Moss J, Ramos FJ, Selicorni A, Tunnicliffe P, Wierzba J, Hennekam RC: Cornelia de Lange syndrome: extending the physical and psychological phenotype. Am J Med Genet A. 2010, 152: 1127-1135.
Rohatgi S, Clark D, Kline AD, Jackson LG, Pie J, Siu V, Ramos FJ, Krantz ID, Deardorff MA: Facial diagnosis of mild and variant CdLS: Insights from a dysmorphologist survey. Am J Med Genet A. 2010, 152: 1641-1653.
Nasmyth K, Haering CH: Cohesin: its roles and mechanisms. Annu Rev Genet. 2009, 43: 525-558. 10.1146/annurev-genet-102108-134233.
Tonkin ET, Wang TJ, Lisgo S, Bamshad MJ, Strachan T: NIPBL, encoding a homolog of fungal Scc2-type sister chromatid cohesion proteins and fly Nipped-B, is mutated in Cornelia de Lange syndrome. Nat Genet. 2004, 36: 636-641. 10.1038/ng1363.
Krantz ID, McCallum J, DeScipio C, Kaur M, Gillis LA, Yaeger D, Jukofsky L, Wasserman N, Bottani A, Morris CA, Nowaczyk MJ, Toriello H, Bamshad MJ, Carey JC, Rappaport E, Kawauchi S, Lander AD, Calof AL, Li HH, Devoto M, Jackson LG: Cornelia de Lange syndrome is caused by mutations in NIPBL, the human homolog of Drosophila melanogaster Nipped-B. Nat Genet. 2004, 36: 631-635. 10.1038/ng1364.
Borck G, Redon R, Sanlaville D, Rio M, Prieur M, Lyonnet S, Vekemans M, Carter NP, Munnich A, Colleaux L, Cormier-Daire V: NIPBL mutations and genetic heterogeneity in Cornelia de Lange syndrome. J Med Genet. 2004, 41: e128-10.1136/jmg.2004.026666.
Gillis LA, McCallum J, Kaur M, DeScipio C, Yaeger D, Mariani A, Kline AD, Li HH, Devoto M, Jackson LG, Krantz ID: NIPBL mutational analysis in 120 individuals with Cornelia de Lange syndrome and evaluation of genotype-phenotype correlations. Am J Hum Genet. 2004, 75: 610-623. 10.1086/424698.
Miyake N, Visser R, Kinoshita A, Yoshiura K, Niikawa N, Kondoh T, Matsumoto N, Harada N, Okamoto N, Sonoda T, Naritomi K, Kaname T, Chinen Y, Tonoki H, Kurosawa K: Four novel NIPBL mutations in Japanese patients with Cornelia de Lange syndrome. Am J Med Genet A. 2005, 135: 103-105.
Borck G, Zarhrate M, Cluzeau C, Bal E, Bonnefont JP, Munnich A, Cormier-Daire V, Colleaux L: Father-to-daughter transmission of Cornelia de Lange syndrome caused by a mutation in the 5′ untranslated region of the NIPBL Gene. Hum Mutat. 2006, 27: 731-735. 10.1002/humu.20380.
Bhuiyan ZA, Klein M, Hammond P, van Haeringen A, Mannens MM, Van Berckelaer-Onnes I, Hennekam RC: Genotype-phenotype correlations of 39 patients with Cornelia De Lange syndrome: the Dutch experience. J Med Genet. 2006, 43: 568-575.
Yan J, Saifi GM, Wierzba TH, Withers M, Bien-Willner GA, Limon J, Stankiewicz P, Lupski JR, Wierzba J: Mutational and genotype-phenotype correlation analyses in 28 Polish patients with Cornelia de Lange syndrome. Am J Med Genet A. 2006, 140: 1531-1541.
Kline AD, Grados M, Sponseller P, Levy HP, Blagowidow N, Schoedel C, Rampolla J, Clemens DK, Krantz I, Kimball A, Pichard C, Tuchman D: Natural history of aging in Cornelia de Lange syndrome. Am J Med Genet C Semin Med Genet. 2007, 145: 248-260.
Schoumans J, Wincent J, Barbaro M, Djureinovic T, Maguire P, Forsberg L, Staaf J, Thuresson AC, Borg A, Nordgren A, Malm G, Anderlid BM: Comprehensive mutational analysis of a cohort of Swedish Cornelia de Lange syndrome patients. Eur J Hum Genet. 2007, 15: 143-149. 10.1038/sj.ejhg.5201737.
Vrouwe MG, Elghalbzouri-Maghrani E, Meijers M, Schouten P, Godthelp BC, Bhuiyan ZA, Redeker EJ, Mannens MM, Mullenders LH, Pastink A, Darroudi F: Increased DNA damage sensitivity of Cornelia de Lange syndrome cells: evidence for impaired recombinational repair. Hum Mol Genet. 2007, 16: 1478-1487. 10.1093/hmg/ddm098.
Chong K, Keating S, Hurst S, Summers A, Berger H, Seaward G, Martin N, Friedberg T, Chitayat D: Cornelia de Lange syndrome (CdLS): prenatal and autopsy findings. Prenat Diagn. 2009, 5: 489-494.
Castronovo P, Delahaye-Duriez A, Gervasini C, Azzollini J, Minier F, Russo S, Masciadri M, Selicorni A, Verloes A, Larizza L: Somatic mosaicism in Cornelia de Lange syndrome: a further contributor to the wide clinical expressivity?. Clin Genet. 2010, 78: 560-564. 10.1111/j.1399-0004.2010.01408.x.
Hosokawa S, Takahashi N, Kitajima H, Nakayama M, Kosaki K, Okamoto N: Brachmann-de Lange syndrome with congenital diaphragmatic hernia and NIPBL gene mutation. Congenit Anom (Kyoto). 2010, 50: 129-132. 10.1111/j.1741-4520.2010.00270.x.
Oliveira J, Dias C, Redeker E, Costa E, Silva J, Reis Lima M, den Dunnen JT, Santos R: Development of NIPBL locus-specific database using LOVD: from novel mutations to further genotype-phenotype correlations in Cornelia de Lange Syndrome. Hum Mutat. 2010, 31: 1216-1222. 10.1002/humu.21352.
Pié J, Gil-Rodríguez MC, Ciero M, López-Viñas E, Ribate MP, Arnedo M, Deardorff MA, Puisac B, Legarreta J, de Karam JC, Rubio E, Bueno I, Baldellou A, Calvo MT, Casals N, Olivares JL, Losada A, Hegardt FG, Krantz ID, Gómez-Puertas P, Ramos FJ: Mutations and variants in the cohesion factor genes NIPBL, SMC1A, and SMC3 in a cohort of 30 unrelated patients with Cornelia de Lange syndrome. Am J Med Genet A. 2010, 152: 924-929.
Bhuiyan ZA, Stewart H, Redeker EJ, Mannens MM, Hennekam RC: Large genomic rearrangements in NIPBL are infrequent in Cornelia de Lange syndrome. Eur J Hum Genet. 2007, 15: 505-508. 10.1038/sj.ejhg.5201776.
Ratajska M, Wierzba J, Pehlivan D, Xia Z, Brundage EK, Cheung SW, Stankiewicz P, Lupski JR, Limon J: Cornelia de Lange syndrome case due to genomic rearrangements including NIPBL. Eur J Med Genet. 2010, 53: 378-382. 10.1016/j.ejmg.2010.08.002.
Russo S, Masciadri M, Gervasini C, Azzollini J, Cereda A, Zampino G, Haas O, Scarano G, Tenconi R, Di Rocco M, Finelli P, Selciorni A, Larizza L: Intragenic and large NIPBL rearrangements revealed by MLPA. Eur J Hum Genet. 2012, 20: 734-741. 10.1038/ejhg.2012.7.
Musio A, Selicorni A, Focarelli ML, Gervasini C, Milani D, Russo S, Vezzoni P, Larizza L: X-linked Cornelia de Lange syndrome owing to SMC1L1 mutations. Nat Genet. 2006, 38: 528-530. 10.1038/ng1779.
Deardorff MA, Kaur M, Yaeger D, Rampuria A, Korolev S, Pie J, Gil-Rodríguez C, Arnedo M, Loeys B, Kline AD, Wilson M, Lillquist K, Siu V, Ramos FJ, Musio A, Jackson LS, Dorsett D, Krantz ID: Mutations in cohesin complex members SMC3 and SMC1A cause a mild variant of cornelia de Lange syndrome with predominant mental retardation. Am J Hum Genet. 2007, 80: 485-494. 10.1086/511888.
Liu J, Feldman R, Zhang Z, Deardorff MA, Haverfield EV, Kaur M, Li JR, Clark D, Kline AD, Waggoner DJ, Das S, Jackson LG, Krantz ID: SMC1A expression and mechanism of pathogenicity in probands with X-Linked Cornelia de Lange syndrome. Hum Mutat. 2009, 30: 1535-1542. 10.1002/humu.21095.
Mannini L, Liu J, Krantz ID, Musio A: Spectrum and consequences of SMC1A mutations: the unexpected involvement of a core component of cohesin in human disease. Hum Mutat. 2010, 31: 5-10. 10.1002/humu.21129.
Deardorff MA, Bando M, Nakato R, Watrin E, Itoh T, Minamino M, Saitoh K, Komata M, Katou Y, Clark D, Cole KE, De Baere E, Decroos C, Di Donato N, Ernst S, Francey LJ, Gyftodimou Y, Hirashima K, Hullings M, Ishikawa Y, Jaulin C, Kaur M, Kiyono T, Lombardi PM, Magnaghi-Jaulin L, Mortier GR, Nozaki N, Petersen MB, Seimiya H, Siu VM, Suzuki Y, Takagaki K, Wilde JJ, Willems PJ, Prigent C, Gillessen-Kaesbach G, Christianson DW, Kaiser FJ, Jackson LG, Hirota T, Krantz ID, Shirahige K: HDAC8 mutations in Cornelia de Lange syndrome affect the cohesin acetylation cycle. Nature. 2012, 489: 313-317. 10.1038/nature11316.
Deardorff MA, Wilde JJ, Albrecht M, Dickinson E, Tennstedt S, Braunholz D, Mönnich M, Yan Y, Xu W, Gil-Rodríguez MC, Clark D, Hakonarson H, Halbach S, Michelis LD, Rampuria A, Rossier E, Spranger S, Van Maldergem L, Lynch SA, Gillessen-Kaesbach G, Lüdecke HJ, Ramsay RG, McKay MJ, Krantz ID, Xu H, Horsfield JA, Kaiser FJ: RAD21 mutations cause a human cohesinopathy. Am J Hum Genet. 2012, 90: 1014-1027. 10.1016/j.ajhg.2012.04.019.
Hayashi S, Ono M, Makita Y, Imoto I, Mizutani S, Inazawa J: Fortuitous detection of a submicroscopic deletion at 1q25 in a girl with Cornelia-de Lange syndrome carrying t(5;13)(p13.1;q12.1) by array-based comparative genomic hybridization. Am J Med Genet A. 2007, 143: 1191-1197.
Gervasini C, Pfundt R, Castronovo P, Russo S, Roversi G, Masciadri M, Milani D, Zampino G, Selicorni A, Schoenmakers EF, Larizza L: Search for genomic imbalances in a cohort of 24 Cornelia de Lange patients negative for mutations in the NIPBL and SMC1L1 genes. Clin Genet. 2008, 74: 531-538. 10.1111/j.1399-0004.2008.01086.x.
DeScipio C, Kaur M, Yaeger D, Innis JW, Spinner NB, Jackson LG, Krantz ID: Chromosome rearrangements in cornelia de Lange syndrome (CdLS): report of a der(3)t(3;12)(p25.3;p13.3) in two half sibs with features of CdLS and review of reported CdLS cases with chromosome rearrangements. Am J Med Genet A. 2005, 137: 276-282.
Lichter P, Cremer T: Chromosome analysis by non-isotopic in situ hybridization. Human cytogenetics. A practical approach. Edited by: Rooney DE, Czepulkowski BH. 1992, Oxford: IRL Press at Oxford University Press, 157-192.
Liu J, Krantz ID: Cornelia de Lange syndrome, cohesin, and beyond. Clin Genet. 2009, 76: 303-314. 10.1111/j.1399-0004.2009.01271.x.
Dorsett D, Ström L: The ancient and evolving roles of cohesin in gene expression and DNA repair. Curr Biol. 2012, 22: R240-R250. 10.1016/j.cub.2012.02.046.
Callier P, Faivre L, Marle N, Thauvin-Robinet C, Sanlaville D, Gosset P, Prieur M, Labenne M, Huet F, Mugneret F: Major feeding difficulties in the first reported case of interstitial 20q11.22-q12 microdeletion and molecular cytogenetic characterization. Am J Med Genet A. 2006, 140A: 1859-1863. 10.1002/ajmg.a.31395.
Iqbal MA, Al-Owain M: Interstitial del(20)(q11.2q12) - clinical and molecular cytogenetic characterization. Am J Med Genet A. 2007, 143A: 1880-1884. 10.1002/ajmg.a.31844.
Hiraki Y, Nishimura A, Hayashidani M, Terada Y, Nishimura G, Okamoto N, Nishina S, Tsurusaki Y, Doi H, Saitsu H, Miyake N, Matsumoto N: A de novo deletion of 20q11.2-q12 in a boy presenting with abnormal hands and feet, retinal dysplasia, and intractable feeding difficulty. Am J Med Genet A. 2011, 155A: 409-414.
Redon R, Rio M, Gregory SG, Cooper RA, Fiegler H, Sanlaville D, Banerjee R, Scott C, Carr P, Langford C, Cormier-Daire V, Munnich A, Carter NP, Colleaux L: Tiling path resolution mapping of constitutional 1p36 deletions by array-CGH: contiguous gene deletion or “deletion with positional effect” syndrome?. J Med Genet. 2005, 42: 166-171. 10.1136/jmg.2004.023861.
D’Angelo CS, Gajecka M, Kim CA, Gentles AJ, Glotzbach CD, Shaffer LG, Koiffmann CP: Further delineation of nonhomologous-based recombination and evidence for subtelomeric segmental duplications in 1p36 rearrangements. Hum Genet. 2009, 125: 551-563. 10.1007/s00439-009-0650-9.
Byrne JLB, Korn GA, Dev VG, Bunch GM, Brooks K, Friedman JM, Harrod MJE: Partial trisomy 19p. Am J Hum Genet A. 1980, 32: 64-
Salbert BA, Solomon M, Spence JE, Jackson-Cook C, Brown J, Bodurtha J: Partial trisomy 19p: case report and natural history. Clin Genet. 1992, 41: 143-146.
Brown J, Horsley SW, Jung C, Saracoglu K, Janssen B, Brough M, Daschner M, Beedgen B, Kerkhoffs G, Eils R, Harris PC, Jauch A, Kearney L: Identification of a subtle t(16;19)(p13.3;p13.3) in an infant with multiple congenital abnormalities using a 12-colour multiplex FISH telomere assay, M-TEL. Eur J Hum Genet. 2000, 8: 903-910. 10.1038/sj.ejhg.5200545.
Quigley DI, Kaiser-Rogers K, Aylsworth AS, Rao KW: Submicroscopic deletion 9(q34.3) and duplication 19(p13.3): identified by subtelomere specific FISH probes. Am J Med Genet A. 2004, 125: 67-72.
Lybaek H, Ørstavik KH, Prescott T, Hovland R, Breilid H, Stansberg C, Steen VM, Houge G: An 8.9 Mb 19p13 duplication associated with precocious puberty and a sporadic 3.9 Mb 2q23.3q24.1 deletion containing NR4A2 in mentally retarded members of a family with an intrachromosomal 19p-into-19q between-arm insertion. Eur J Hum Genet. 2009, 17: 904-910. 10.1038/ejhg.2008.261.
Mayor T, Hacker U, Stierhof YD, Nigg EA: The mechanism regulating the dissociation of the centrosomal protein C-Nap1 from mitotic spindle poles. J Cell Sci. 2002, 115 (Pt 16): 3275-3284.
Yang Y, Wu F, Ward T, Yan F, Wu Q, Wang Z, McGlothen T, Peng W, You T, Sun M, Cui T, Hu R, Dou Z, Zhu J, Xie W, Rao Z, Ding X, Yao X: Phosphorylation of HsMis13 by Aurora B kinase is essential for assembly of functional kinetochore. J Biol Chem. 2008, 283: 26726-26736. 10.1074/jbc.M804207200.
Cheeseman IM, Desai A: Molecular architecture of the kinetochore-microtubule interface. Nat Rev Mol Cell Biol. 2008, 9: 33-46. 10.1038/nrm2310.
Medendorp K, Vreede L, van Groningen JJ, Hetterschijt L, Brugmans L, Jansen PA, van den Hurk WH, de Bruijn DR, van Kessel AG: The mitotic arrest deficient protein MAD2B interacts with the clathrin light chain A during mitosis. PLoS One. 2010, 5: e15128-10.1371/journal.pone.0015128.
Amano M, Suzuki A, Hori T, Backer C, Okawa K, Cheeseman IM, Fukagawa T: The CENP-S complex is essential for the stable assembly of outer kinetochore structure. J Cell Biol. 2009, 186: 173-182. 10.1083/jcb.200903100.
Li M, Xu X, Liu Y: The SET2-RPB1 interaction domain of human RECQ5 is important for transcription-associated genome stability. Mol Cell Biol. 2011, 31: 2090-2099. 10.1128/MCB.01137-10.
Islam MN, Fox D, Guo R, Enomoto T, Wang W: RecQL5 promotes genome stabilization through two parallel mechanisms–interacting with RNA polymerase II and acting as a helicase. Mol Cell Biol. 2010, 30: 2460-2472. 10.1128/MCB.01583-09.
Schwendener S, Raynard S, Paliwal S, Cheng A, Kanagaraj R, Shevelev I, Stark JM, Sung P, Janscak P: Physical interaction of RECQ5 helicase with RAD51 facilitates its anti-recombinase activity. J Biol Chem. 2010, 285: 15739-15745. 10.1074/jbc.M110.110478.
Wäsch R, Robbins JA, Cross FR: The emerging role of APC/CCdh1 in controlling differentiation, genomic stability and tumor suppression. Oncogene. 2010, 29: 1-10. 10.1038/onc.2009.325.
Nilsson J, Yekezare M, Minshull J, Pines J: The APC/C maintains the spindle assembly checkpoint by targeting Cdc20 for destruction. Nat Cell Biol. 2008, 10: 1411-1420. 10.1038/ncb1799.
Jin L, Williamson A, Banerjee S, Philipp I, Rape M: Mechanism of ubiquitin-chain formation by the human anaphase-promoting complex. Cell. 2008, 133: 653-665. 10.1016/j.cell.2008.04.012.
Wathugala DL, Hemsley PA, Moffat CS, Cremelie P, Knight MR, Knight H: The Mediator subunit SFR6/MED16 controls defence gene expression mediated by salicylic acid and jasmonate responsive pathways. New Phytol. 2012, 10.1111/j.1469-8137.2012.04138.x. Epub ahead of print
Noh EJ, Lim DS, Lee JS: A novel role for methyl CpG-binding domain protein 3, a component of the histone deacetylase complex, in regulation of cell cycle progression and cell death. Biochem Biophys Res Commun. 2009, 378: 332-337. 10.1016/j.bbrc.2008.11.079.
Saito M, Ishikawa F: The mCpG-binding domain of human MBD3 does not bind to mCpG but interacts with NuRD/Mi2 components HDAC1 and MTA2. J Biol Chem. 2002, 277: 35434-35439. 10.1074/jbc.M203455200.
Hutchins JR, Toyoda Y, Hegemann B, Poser I, Hériché JK, Sykora MM, Augsburg M, Hudecz O, Buschhorn BA, Bulkescher J, Conrad C, Comartin D, Schleiffer A, Sarov M, Pozniakovsky A, Slabicki MM, Schloissnig S, Steinmacher I, Leuschner M, Ssykor A, Lawo S, Pelletier L, Stark H, Nasmyth K, Ellenberg J, Durbin R, Buchholz F, Mechtler K, Hyman AA, Peters JM: Systematic analysis of human protein complexes identifies chromosome segregation proteins. Science. 2010, 328: 593-599. 10.1126/science.1181348.
Baynam G, Goldblatt J, Walpole I: Deletion of 8p23.1 with features of Cornelia de Lange syndrome and congenital diaphragmatic hernia and a review of deletions of 8p23.1 to 8pter? A further locus for Cornelia de Lange syndrome. Am J Med Genet A. 2008, 46A: 1565-1570.
Liu J, Zhang Z, Bando M, Itoh T, Deardorff MA, Clark D, Kaur M, Tandy S, Kondoh T, Rappaport E, Spinner NB, Vega H, Jackson LG, Shirahige K, Krantz ID: Transcriptional Dysregulation in NIPBL and Cohesin Mutant Human Cells. PLoS Biol. 2009, 7: e1000119-10.1371/journal.pbio.1000119.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2350/14/41/prepub
Acknowledgements
We would like to thank the patients families for participating in this study, and the clinicians who provided some of the patients negative for known causative mutations.
This study was supported by a Ministry of Health “Ricerca Corrente” grant to Istituto Auxologico Italiano IRCCS (08C001-2010) and by a “Accordo quadro Università-Regione Lombardia n° 17292” grant to LL and PF.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare they have no competing interests.
Author’s contributions
CG, LL and PF conceived and designed the study and wrote the manuscript. CP and DR performed the aCGH work and the FISH experiments. CG, JA, MM and SR performed the mutation screening of NIPBL and SMC1A. CG, CP, JA and PF contributed to interpretation of the aCGH results. AS, GZ, RT, AC and CM recruited the patients and reviewed the clinical data. All of the authors approved the final version of the manuscript.
Electronic supplementary material
12881_2012_1071_MOESM1_ESM.xls
Additional file 1: Table S1: Auxological parameters (at birth and at age of evaluation) of probands 1–4 compared with those characteristic of CdLS patients. (XLS 29 KB)
12881_2012_1071_MOESM3_ESM.pdf
Additional file 3: Figure S1: 20q11.2q12 map showing region involved in our patient 1 rearrangement compared to molecularly characterized 20q deletions reported in the literature. (PDF 35 KB)
12881_2012_1071_MOESM4_ESM.xls
Additional file 4: Table S3: Clinical signs of proband 1 compared with those characteristic of CdLS patients and patients carrying an overlapping 20q deletion. (XLS 35 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Gervasini, C., Picinelli, C., Azzollini, J. et al. Genomic imbalances in patients with a clinical presentation in the spectrum of Cornelia de Lange syndrome. BMC Med Genet 14, 41 (2013). https://doi.org/10.1186/1471-2350-14-41
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2350-14-41