
Abstract
Background
The large gene superfamily of ABC (ATP-binding cassette) transporters encodes membrane proteins involved in trafficking processes across biological membranes and further essential cell biological functions. ABC transporters are evolutionary ancient and involved in the biochemical defence against toxicants. We report here a genome-wide survey of ABC proteins of Daphnia pulex, providing for the first time information on ABC proteins in crustacea, a primarily aquatic arthropod subphylum of high ecological and economical importance.
Results
We identified 64 ABC proteins in the Daphnia genome, which possesses members of all current ABC subfamilies A to H. To unravel phylogenetic relationships, ABC proteins of Daphnia were compared to those from yeast, worm, fruit fly and human. A high conservation of Daphnia of ABC transporters was observed for proteins involved in fundamental cellular processes, including the mitochondrial half transporters of the ABCB subfamily, which function in iron metabolism and transport of Fe/S protein precursors, and the members of subfamilies ABCD, ABCE and ABCF, which have roles in very long chain fatty acid transport, initiation of gene transcription and protein translation, respectively. A number of Daphnia proteins showed one-to-one orthologous relationships to Drosophila ABC proteins including the sulfonyl urea receptor (SUR), the ecdysone transporter ET23, and the eye pigment precursor transporter scarlet. As the fruit fly, Daphnia lacked homologues to the TAP protein, which plays a role in antigene processing, and the cystic fibrosis transmembrane conductance regulator (CFTR), which functions as a chloride channel. Daphnia showed two proteins homologous to MDR (multidrug resistance) P-glycoproteins (ABCB subfamily) and six proteins homologous to MRPs (multidrug resistance-associated proteins) (ABCC subfamily). However, lineage specific gene duplications in the ABCB and ABCC subfamilies complicated the inference of function. A particularly high number of gene duplications were observed in the ABCG and ABCH subfamilies, which have 23 and 15 members, respectively.
Conclusion
The in silico characterisation of ABC transporters in the Daphnia pulex genome revealed that the complement of ABC transporters is as complex in crustaceans as that other metazoans. Not surprisingly, among currently available genomes, Daphnia ABC transporters most closely resemble those of the fruit fly, another arthropod.
Similar content being viewed by others


Background
ATP-binding cassette (ABC) proteins constitute one of the largest protein superfamilies and are present in all organisms from bacteria to human [1, 2]. Prototypical ABC proteins are membrane-bound transporters coupling ATP hydrolysis to the translocation of substrates across biological membranes [3, 4]. In addition to transporters, ABC proteins also comprise ion channels, regulators of ion channels, receptors, and proteins with roles in ribosome assembly and translation. The human genome has 48 genes encoding ABC proteins, of which 17 have been linked to hereditary diseases, including cystic fibrosis, adrenoleukodystrophy, Stargardt disease and disorders of cholesterol metabolism [2, 5, 6].
ABC proteins share a conserved domain architecture. A functional transporter requires the cooperation of two transmembrane domains (TMD) and two cytosolic nucleotide binding domains (NBDs, also called ATP-binding cassettes). Eukaryotic ABC proteins are either full transporters combining all required domains in one polypeptide (2 TMDs and 2 NBDs), or half-transporters consisting of 1 TMD and 1 NBD that need to form homo- or heterodimers to constitute a functional pump. According to their domain architecture and sequence, metazoan ABC transporters are divided into subfamilies, of which seven (A to G) exist in human [7]. An eighth subfamily (H) has been defined following the analysis of the Drosophila melanogaster genome [2]. The H subfamily is missing in mammals, but has one member in zebrafish [8]. The members of subfamilies E and F are not transporters and differ from other ABC proteins in that they possess two NBDs but lack TMDs. ABCE proteins are inhibitors of RNAse L and involved in the assembly of the preinitiation complex [9–11], while ABCF proteins have roles in ribosome assembly and protein translation [12, 13].
The first eukaryotic ABC transporter discovered was the human (h) drug efflux transporter MDR (multidrug resistance) P-glycoprotein (hABCB1/MDR1), the name of which reflects that its expression in cancers can cause a decreased cellular drug accumulation (initially referred to as drug permeability, 'P'), resulting in the resistance of tumours against chemotherapy [14, 15]. Subsequent studies have identified further ABC proteins that are drug efflux pumps and can cause MDR in cancers, including the multidrug resistance associated protein (hABCC1/MRP1) [16] and the breast cancer related protein (hABCG2/BCRP) [17]. Drug efflux transporters are found in ABC subfamilies B, C and G [18], and in normal tissues often show an apical expression in epithelia involved in excretion or forming boundaries of the body, reflecting their role in the biochemical defence against toxicants [19]. ABC drug efflux transporters have a wide phylogenetic distribution and are found in vertebrates as well as in deuterostome invertebrates (sea squirt Ciona intestinalis [8]; sea urchin Strongylocentrotus purpuratus [20]), protostome invertebrates (nematode Caenorhabditis elegans [21]; fruitfly Drosophila melanogaster [2]), protozoans and yeast [22, 23]. Homologous proteins are also present in plants, though their roles in multidrug efflux have not been firmly established [24, 25].
The planktic crustacean Daphnia is globally distributed and has central importance for the ecology of lakes and ponds. The currently accomplished sequencing of the Daphnia pulex genome will thus enhance research in disciplines which traditionally have made use of daphnids, e.g., ecology, physiology, toxicology, population genetics and behaviour. Moreover, studies with an evolutionary perspective are further expected to benefit, because Daphnia is not only the first crustacean, but also the first non-insect arthropod to have its genome sequence determined. The aim of this study was to provide a survey of the ABC transporter gene family of Daphnia pulex. A complete or close to complete list of ABC transporters in Daphnia will facilitate the identification of genes that play a role during the adaptation of Daphnia to environmental toxicants. Furthermore, ABC genes have been suggested as biochemical factors contributing to the phenomenon of resistance against chemotherapeutics in parasites [22, 26] and insects [27, 28]. In the aquaculture industry, parasitic crustaceans constitute a problem of considerable economic importance. The potential development of resistances against therapeutics used to control the crustacean parasite sea louse (Lepeophtheirus salmonis) in salmon farming is currently becoming a concern, and it has recently been proposed that ABC transporters in the sea louse could represent potential biochemical resistance factors to emamectin, a therapeutic used to control sea louse infestations in salmon [29, 30]. At the same time, the sulfonylurea receptor has been suggested to represent the target for the chitin synthesis inhibitor diflubezuron [31], a compound also used to treat sea lice infections. Thus, ABC transporters might have relevance in crustaceans both as biochemical defence mechanisms against toxicants, and as targets of toxicity.
From an evolutionary perspective, the wide distribution of ABC transporters capable of drug efflux transport suggests they are of ancestral origin. However, ABC subfamilies containing drug transporters (B, C, and G) also comprise proteins with other functions. For instance, the arguably best known drug efflux pump hABCB1/MDR1 belongs to the B subfamily that also contains mammalian transporters of bile salts and phospholipids, and yeast transporters of pheromones. Because of the lack of clear orthologous relationships between ABC proteins of the nematode worm Caenorhabditis elegans and those of other genomes, it has been suggested that drug efflux pumps have evolved independently several times [21]. The annotation of ABC transporters in the Daphnia genome, provided by this work, represents an important resource for future biochemical, toxicological and physiological studies of ABC drug efflux transporters
Results and discussion
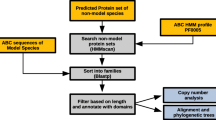
To identify gene loci encoding ABC transporters, multiple tblastn searches were performed on the Dappu v1.1 draft genome sequence assembly (September, 2006) [32] using NBDs of different Drosophila melanogaster ABC proteins as queries (one search per subfamily). The most plausible gene model was selected among machine-generated models available at wFleaBase http://wFleaBase.org and the JGI genome portal http://www.jgi.doe.gov/Daphnia, and its NBDs extracted for phylogenetic analysis that further included NBDs of fruit fly and human ABC transporters. This resulted in a subfamily-specific clustering of NBDs, with N- and C-terminal NBDs of full transporters generally allocated to distinct sub-clusters (data not shown). Based on the clustering of NBDs, Daphnia transporters were assigned to ABC subfamilies. Gene models were refined on the basis of sequence homology and EST support, and the subfamily assignment confirmed by protein BLAST analyses on the National Center for Biotechnology Information website. Using this strategy, 64 loci of putative Daphnia ABC genes were identified, for 48 of which evidence of mRNA expression was present in EST databases implemented in the JGI genome portal and wFeaBase (10,392 assembled cDNA on genome scaffolds) (Table 1). The majority of potential Daphnia ABC genes lacking EST support (14 of 16) were found in subfamilies G and H (Table 1), in which a high number of gene duplications was observed (see below for further discussion). Significant sequence homologies to the query NBDs were observed at further 15 loci that were excluded from further analysis [see additional file 1]. Of these loci, four corresponded to obvious pseudogenes, while two most likely represent bacterial contamination. At the remaining nine excluded loci, the best obtainable gene models were fragmentary, i.e. showing lack/incompleteness of vital domains, with many models being affected by sequence gaps. All excluded loci lacked EST evidence of expression.
To analyse the evolutionary position of the 64 putative Daphnia ABC transporters identified, phylogenetic analyses compared the complement of ABC proteins in the Daphnia genome to that in the genomes of yeast (Saccharomyces cervisiae), fruit fly (Drosophila melanogaster), worm (Caenorhabditis elegans) and human (Homo sapiens), using predicted (Daphnia) or database derived protein sequences [see additional file 2]. For two reasons, separate analyses were carried out for each subfamily. Firstly, the domain architecture of ABC transporters is highly variable among subfamilies, which complicates bioinformatic analyses on whole length sequences of the whole protein family. Secondly, while the NBDs of different transporters show a similar length and organisation, their sequence is too conserved to provide a meaningful degree of resolution in phylogenetic analysis.
ABCA
ABCA subfamily proteins are full transporters characterised by distinctive conserved traits, in particular a large extracellular loop between the first two transmembrane helices of each TMD, and a family specific motif located C-terminal of each NBF [33]. Four ABCA subfamily transporters presenting these hallmarks (data not shown) have been identified in Daphnia (Table 1). An analysis of the evolutionary relationship of these transporters to human, worm, and fruit fly ABCA proteins is shown in Fig. 1. Dappu-(Daphnia pulex)312055 and Dappu-312056 are neighbouring genes showing a head-to-tail orientation (Table 1) and display 59% amino acid identity, suggesting they are the result of a tandem duplication. Dappu-312055 and Dappu-312056 group together with hABCA1/ABC1, hABCA2, hABCA4/ABCR and hABCA7. Dappu-346971 groups together with the hABCA5 cluster (hABCA5, 6, 8, 9, and 10). Dappu-347506 groups together with hABCA3. In mammals, ABCA proteins adopt critical roles in the control of cellular lipid transport processes. Loss-of-function scenarios in human monogenetic diseases and mouse knockout models have revealed roles of hABCA1/ABC1 in HDL biogenesis, of hABCA3 in lung surfactant production, of hABCA4/ABCR in retinal integrity and of ABCA12 in keratinisation processes in the skin [34]. Thus, while ABCA proteins share a functional relation to lipid trafficking, individual transporters in this subfamily have adopted highly specialised roles in phospho- and sphingolipid export machineries [34]. In consequence, it is not possible to assign specific putative roles to the Daphnia ABCA transporters, though it appears likely that they are involved in lipid trafficking processes.

Phylogenetic tree of ABCA proteins in four eukaryotic genomes. Predicted amino acid sequences were aligned using ClustalX, and the alignment used to generate a phylogenetic tree using the neighbour joining method [74]. Bootstrapping was used to determine the relative support of the various branches (5,000 replicates, support expressed as percent) [74]. Dp, Daphnia pulex,; Dm Drosophila melanogaster; Ce Caenorhabditis elegans, h Homo sapiens; Sc Saccharomyces cervisia.
ABCB
The ABCB family can be divided into a group of full transporters (FT) that includes the drug efflux pump hABCB1/MDR1, and a group of half transporters (HT). Two ABCB FTs and five ABCB HTs were found in the Daphnia genome (Table 1). The evolutionary analysis of ABCB FT assigned the human, worm, and arthropod (combined Drosophila and Daphnia) transporters into clearly distinguished clades (Fig. 2), suggesting that this subfamily has diversified through lineage-specific gene duplications. Early reports have noted the similarity in sequence of three Drosophila ABCB FTs and hABCB1/MDR1, and named these genes mdr49, mdr50, and mdr65 [35, 36]. While no further information is available on mdr50, the Drosophila gene most closely related to the two Daphnia ABCB FTs, a number of studies suggests a role of mdr49 and to a lesser extent also mdr65 in the biochemical defence against toxicants. The disruption of the mdr49 gene resulted in an increase in colchicine resistance [35]. A genetic polymorphism related to α-amatin resistance in Drosophila was mapped to the region of the mdr65 gene [37]. Colchicine exposure and heat shock increases the expression of mdr49, but not mdr65, in Drosophila larvae, while both genes were induced in tumours [38]. Mdr49 was further found to be induced by polycyclic aromatic hydrocarbons, and shown to be involved in the transport of these chemicals [39].

Phylogenetic tree of ABCB subfamily full transporters in five eukaryotic genomes, derived according to the procedure outlined in the legend to Fig. 1.
The phylogenetic analysis of ABCB HT revealed comparatively clear orthologue relations (Fig. 3). Human mitochondrial transporters hABCB6, hABCB7, hABCB8/MABC1 and hABCB10/MABC2 function in iron metabolism and transport of Fe/S protein precursors. The hABCB6, hABCB7, and hABCB8/MABC1 proteins each have one orthologue in Daphnia (Dappu-347268, Dappu-347270, and Dappu-347266, respectively). The hABCB10/MABC2 protein has two Daphnia co-orthologues, Dappu-347275 and Dappu-347276, which show high similarity (83% amino acid identity). The human transporter associated with antigen processing (TAP) is a heterodimer of two ABCB proteins, hABCB2/TAP1 and ABCB3/TAP2 [40]. TAP translocates peptides derived from proteasomal degradation from the cytosol to the lumen of the endoplasmic reticulum, where their loading onto major histocompatibility complex (MHC) class I molecules occurs [40]. The function of hABCB9/TAPL (TAP-like) is currently unknown and its subcellular localisation is still under discussion [41]. It has been shown that TAPL is present in the lysosomal compartment [42], and it has been recently suggested that it might be involved in peptide presentation to MHC class II in dendritic cells [41]. As invertebrates lack the mammalian adaptive immune response, the lack of TAP/TAPL homologues in Daphnia and Drosophila (Fig. 3) is not unexpected. However, the presence of proteins orthologous to TAP/TAPL in C. elegans (Fig. 3) suggests that these protein transporters might have further roles unrelated to antigen presentation.

Phylogenetic tree of ABCB subfamily half transporters in five eukaryotic genomes, derived according to the procedure outlined in the legend to Fig. 1.
ABCC
The ABCC subfamily is functionally diverse, comprising the chloride channel CFTR (cystic fibrosis transmembrane conductance regulator), the membrane-bound receptors SURs (sulfonylurea receptors), and broad-specificity transporters called multidrug resistance-associated proteins (MRPs) that translocate a range of substrates including drugs, endogenous compounds and their glutathione and glucuronyl conjugates, glutathione, and cyclic nucleotides [43–45]. ABCC subfamily proteins are full-transporters showing two TMDs and two NBDs. Within the human ABCC family, hABCC8/SUR1, hABCC9/SUR2 and certain MRPs called 'long' MRPs are unique in that they possess an additional N-terminal TMD called TMD0 which is lacking in hABCC7/CFTR and the remaining 'short' MRPs [46, 47]. Searches of the Daphnia genome identified seven ABCC transporters (Table 1). The phylogenetic tree of Daphnia, human, worm, and Drosophila ABCC proteins is characterised by comparatively low support of the more basal nodes, most probably reflecting the heterogeneity of the ABCC subfamily (Fig 4). Dappu-442500 is found in the same, well supported clade as human and Drosophila SURs, suggesting it is a SUR homologue. Indeed, Dappu-442500 displays a general architecture of SUR and 'long' MRPs, possessing the additional N-terminal TMD0 (Table 1). Moreover, Dappu-442500 shows a significant conservation of two SUR typical N-terminal motifs, the sulphonylurea receptor family signature (PR01092) and the sulphonylurea receptor type 1 family signature (PR01093), with predicted amino acid similarities to hABCC8/SUR1 of 73% and 61%, respectively (data not shown). Together with previous reports of SUR-typical functional traits of the Drosophila protein CG5772 [48, 49], the data strongly suggests that Dappu-442500 is indeed a SUR. Further Daphnia ABCC proteins possessing an additional N-terminal TMD0 are Dappu-347281 and Dappu-347323 (Table 1). In our phylogenetic analysis Dappu-347281 groups together with Drosophila CG6214 (Fig 4), which is a long MRP resembling hABCC1/MRP1 but awaiting in-depth functional characterisation [50, 51]. According to the tree obtained in this study, Dappu-347323 is a putative orthologue of hABCC10/MRP7 and Drosophila CG7806 (Fig. 4). hABCC10/MRP7 is capable of conferring a hABCC1/MRP1-type multidrug resistance phenotype in cellular models, but its physiological function is at present poorly understood [52]. Functional data are lacking on CG7806. The remaining Daphnia MRPs, Dappu-347288, Dappu-347292, Dappu-347295, and Dappu-347548, show the structural traits of 'short' MRPs (Table 1). The hABCC4/MRP4 protein groups together with Dp347288 in a clade of moderately low bootstrap support (Fig. 4) and is the ABCC transporter showing the greatest similarity to the Daphnia protein (42% amino acid identity). Dappu-347292, Dappu-347295, and Dappu-347548 group together with hABCC5/MRP5 in a clade of high bootstrap support (Fig. 4). The hABCC4/MRP4 and hABCC5/MRP5 proteins have been reported to be able to transport cyclic nucleotides in vitro, but the in vivo relevance of this observation is controversial and their physiological roles remain to be unravelled [53]. Dappu-347288 groups together in one clade with hABCC4/MRP4 and a cluster of Drosophila ABCC proteins including CG10505 (Fig. 4). CG10505 is regulated by heavy metals via the metal-responsive transcription factor 1, and has been shown to be involved in biochemical detoxification of zinc and copper [54]. This parallels previous studies showing that cell lines overexpressing hABCC1/MRP1 are resistant to arsenite and antimony (reviewed in [47]). Together these data suggest Daphnia MRPs could represent potential biochemical factors in the defence against toxicants; however, in the absence of functional data and in the view of the complex phylogeny of the ABCC subfamily this remains at present speculative.

Phylogenetci tree of ABCC proteins in five eukaryotic genomes, derived according to the procedure outlined in the legend to Fig. 1.
ABCD
The ABCD subfamily contains half transporters located to the peroxisome that are involved in the import of fatty acids and/or fatty acyl-CoAs into this organelle [55]. The simultaneous posttranscriptional silencing of three C. elegans ABCD transporters disrupted offspring production in a previous study, suggesting developmental roles of peroxisomal ABC transporters [56]. Mutations in the hABCD1/ALDP gene are the principal inherited defect in adrenoleukodystrophy, a clinically heterogeneous X-linked recessive disorder characterised by adrenal insufficiency and neuronal demyelination [57]. This study identified three ABCD transporters in the Daphnia genome, Dappu-347330, Dappu-347326, and Dappu-303977 (Table 1). The phylogenetic analysis revealed that three clades of ABCDs exist in metazoans, each of which has one Daphnia member (Fig 5). The high degree of conservation (amino acid identity of Daphnia ABCD proteins to the closest human homologue between 46% and 55%) and the clear structure of the phylogenetic tree are consistent with the notion that the function of ABCD proteins in Daphnia might resemble that in other metazoans.

Phylogenetic tree of ABCD proteins in five eukaryotic genomes, derived according to the procedure outlined in the legend to Fig. 1.
ABCE and ABCF
The subfamilies ABCE and ABCF are comprised of atypical ABC proteins that contain a pair of linked nucleotide binding domains and lack transmembrane domains [58]. The proteins of both subfamilies are highly conserved across evolutionary diverse taxa, suggesting their role in fundamental cell biological processes. Most eukaryotes possess one ABCE protein, and Daphnia conforms to this rule (Dappu-189585, Table 1). Human ABCE1/RNaseLI was initially identified as an inhibitor of RNase L [59]. Recent data indicate that human and yeast ABCE proteins have further a central role in translation initiation [60]. ABCF proteins have roles in ribosome assembly and/or protein translation [13]. Four ABCF proteins have been identified in Daphnia (Table 1). The phylogenetic analysis of ABCE and ABCF proteins was carried out together (Fig. 6). As expected, Dappu-189585 fell into the clade containing other ABCE proteins. ABCF proteins were divided into four well-supported clades, one of which contained only yeast proteins, while the remaining three each contained one of the human ABCFs (Fig. 6). Of the Daphnia ABCF proteins, of which Dappu-347357 and Dappu-347363 show 53% amino acid identity and are homologues to hABCF1, while Dappu-304799 and Dappu-347354 are homologues to hABCF2 and hABCF3, respectively.

Phylogenetic tree of ABCE and ABCF proteins in five eukaryotic genomes, derived according to the procedure outlined in the legend to Fig. 1.
ABCG
As half transporters, ABCG proteins need to form homo- or hetero dimers to form a functional pump. In contrast to other ABC half transporters, ABCG proteins show a reverse domain architecture, with the TMD being located C-terminally of the NBD. In human, hABCG1/WHITE1 and hABCG4 are involved in cellular cholesterol efflux to high density lipoprotein, hABCG5 and hABCG8 mediate the intestinal and biliary efflux of cholesterol, plant, and shellfish sterols, while hABCG2/BCRP is a drug efflux pump [61]. The most intensively studied ABCG protein is the white protein in Drosophila, which dimerises with ABCG proteins brown and scarlet to function as a transporter of eye pigment precursors [62]. In yeast and in plants, certain ABCG subfamily proteins are reverse full transporters and called PDR (pleiotropic drug resistance) proteins [63]. Because of their duplicated domain structure (NBD-TM-NBD-TM), yeast PDRs were not included in the phylogenetic analyses of this study.
The identification of Daphnia ABC transporters revealed that in this species ABCG proteins form the largest ABC subfamily (23 members, Table 1). This parallels the situation in Drosophila [2]. The phylogenetic analysis of the ABCG subfamily revealed that the high number of ABCG genes in Daphnia and Drosophila is due to extensive lineage specific gene duplications (Fig. 7). It is know that the genomes of flies and worms contain a large number of duplicated genes, with a greater number of tandem or locally duplicated genes in the C. elegans than the Drosophila genome [64]. A large number of C. elegans annotated genes might be pseudogenes [65, 66]. A similar situation seems to exist in Daphnia, at least with respect to the ABCG (and ABCH, see next section) family, which comprises several clusters of putative genes located in close vicinity displaying high predicted amino acid sequence similarity (up to 86%), and which further show a high number of loci lacking EST support (10 of 23 ABCG proteins Table 1). It seems premature, however, to conclude that the non-expressed putative ABC gene loci (Table 1) are pseudogenes. For instance, a detailed functional analysis of tandem duplicated ABC genes in the worm has revealed differential function of the duplicates, with expression being highly tissue or stage specific [67].

Phylogenetic tree of ABCG proteins in five eukaryotic genomes, derived according to the procedure outlined in the legend to Fig. 1.
In the phylogenetic analysis of ABCG proteins from the Daphnia and other genomes, Dappu-347419 and nine further Daphnia proteins forms a clade (Fig 7, top of tree) that consists of a cluster of eight neighbouring genes on scaffold 7 and two neighbouring genes on scaffold 37 (Table 1). Further down, Dappu-347444 groups together with Drosophila proteins CG3164 and atet (ABC transporter expressed in trachea, [68]) as well as hABCG1 and 4. Dappu-347416 is a homologue of the Drosophila ET23 gene product protein CG3327, an ABCG transporter believed to regulate intracellular ecdysone concentrations during development [69]. Phylogenetic analyses grouped Dappu-258299 together in one clade with hABCG5 and 8, and two Drosophila proteins of unknown function. Among the Drosophila eye pigment transporters, the protein brown (CG17632) does not have a Daphnia orthologue, Dappu-347393 is an orthologue of the scarlet protein (CG4314), while the Drosophila white protein has nine co-orthologues in Daphnia. Assuming that these proteins might adopt roles in eye pigment transport in Daphnia appears a reasonable hypothesis. However, it is noteworthy that recent evidence in Drosophila suggests additional neurobiological functions for the white protein [70, 71].
ABCH
The ABCH subfamily is lacking members in mammals and C. elegans, and has been identified for the first time in Drosophila [2]. At present, teleost fish are the only vertebrates known to possess ABCH transporters [6, 8]. ABCH proteins are inverse half-transporters showing the same domain architecture as the members of the ABCG subfamily. The function of ABCH proteins is yet unknown. In Daphnia, this subfamily is with 15 members the second largest ABC subfamily (Table 1). The phylogenetic analysis assigned Daphnia and Drosophila proteins in distinct clades (Fig. 8), suggesting that the diversity of ABCH subfamily in Daphnia has arisen from lineage specific gene duplications. As observed with the ABCG subfamily, clusters of neighbouring genes displaying high similarity are found among Daphnia ABCH proteins (Table 1).

Phylogenetic tree of ABCH proteins in five eukaryotic genomes, derived according to the procedure outlined in the legend to Fig. 1.
Conclusion
The identification and phylogenetical analysis of ABC transporters in the Daphnia pulex genome, the first available crustacean genome, has revealed interesting traits of this important group of proteins in Daphnia. The complement of ABC proteins shows parallels between two arthropods Daphnia and Drosophila in that both lack homologues to the human proteins TAP, protein translocators involved in antigen processing, and CFTR, a chloride channel regulated by ATP. The high conservation in Daphnia of ABC proteins involved in fundamental cellular processes, such as the mitochondrial ABCB half transporters and the members of subfamilies ABCD, ABCE and ABCF, confirms an earlier study [21] in strongly suggesting an evolutionary ancestral origin of these proteins. Clear orthologous relationships were observed in further occasions, providing the basis for a tentative function assignment to Daphnia homologues of SUR, a membrane receptor regulating a potassium channel, and to Daphnia homologues of the Drosophila genes ET23 and scarlet. The presence of proteins in Daphnia that resemble the ABC drug efflux transporters hABCB1/MDR1 and hABCC1/MRP1 is in accordance with the hypothesis that these Daphnia proteins might adopt roles in the biochemical defence against toxicants; however, the lack of clear orthologous relationships makes the inference of function in these cases uncertain. A high number of duplicated genes was observed in the ABCG and ABCH subfamilies of Daphnia. This parallels the situation in Drosophila and Caenorhabditis, which also show lineage specific duplications, albeit in other subfamilies. More research is needed to unravel the physiological significance of gene duplications in Daphnia.
Methods
To identify loci of genes encoding ABC transporters, tblastn searches were performed on v1.1 (September, 2007) of the Daphnia pulex draft genome sequence assembly [32] using the highly conserved NBD [72] of different Drosophila melanogaster ABC proteins as queries. One search was carried out per subfamily, each using the sequence of the NBD (as defined by interpro domain IPR003439) of a representative Drosophila protein (A: CG1718; B: CG3879 (mdr49); C: CG9270; D: CG12703; E: CG5651; F: CG9330; G: white; H: CG9990). If the Drosophila transporter had two NBDs, the N-terminal domain was used. Hits from individual subfamily specific tblastn searches (E-value of 10-5) significantly overlapped, with each search retrieving loci of genes of the query and other subfamilies. Increasing the E-value to 10-4 increased the degree of overlap between individual searches, but had no effect of the total number of loci retrieved, suggesting that our search strategy gave an exhaustive representation of those sequences in the current version of the Daphnia pulex genome that show homology to ABC-transporter NBD domains. For each locus identified in tblastn searches, one preliminary gene model representing the NBD hit with reasonable fidelity was chosen from the gene models automatically generated by the JGI pipeline. To assign putative Daphnia ABC genes to ABC subfamilies, the NBDs of preliminary gene models were extracted using the ScanProsite facility [73] with predicted protein sequences and the prosite profile PS50893. NBDs were then subjected to a phylogenetic analysis together with those of fruit fly and human ABC transporters, using neighbour joining and bootstrapping with 5000 replicates in the program package MEGA4 [74]. NBFs of the fruit fly and human ABC transporters formed subfamily specific clusters, with separate groupings for the N- and C-terminal NBFs of full transporters (data not shown). This allowed unequivocal assignment of Daphnia transporters to ABC subfamilies. Gene models were then refined on the basis of homology and EST support. Both the tandem architecture of ABC transporters of certain subfamilies and the commonness of tandem gene duplications in Daphnia made manual adjustments to the gene models necessary, as machine generated gene models would sometimes combine exons from distinct genes. The subfamily assignment of Daphnia ABC proteins was confirmed by protein BLAST analyses of the manually corrected models on the National Center for Biotechnology Information website. Separate phylogenetic analyses on Daphnia, yeast, worm, fruitfly and human ABC transporters were then carried out per subfamily, using the same methodology as above [74] on full sequences.
References
Dassa E, Bouige P: The ABC of ABCs: a phylogenetic and functional classification of ABC systems in living organisms. Res Microbiol. 2001, 152: 211-229. 10.1016/S0923-2508(01)01194-9.
Dean M, Hamon Y, Chimini G: The human ATP-binding cassette (ABC) transporter superfamily. J Lipid Res. 2001, 42: 1007-1017.
Hollenstein K, Frei DC, Locher KP: Structure of an ABC transporter in complex with its binding protein. Nature. 2007, 446: 213-216. 10.1038/nature05626.
Dawson RJ, Locher KP: Structure of a bacterial multidrug ABC transporter. Nature. 2006, 443: 180-185. 10.1038/nature05155.
Gottesman MM, Ambudkar SV: Overview: ABC transporters and human disease. J Bioenerg Biomembr. 2001, 33: 453-458. 10.1023/A:1012866803188.
Dean M, Annilo T: Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Annu Rev Genomics Hum Genet. 2005, 6: 123-142. 10.1146/annurev.genom.6.080604.162122.
Klein I, Sarkadi B, Váradi A: An inventory of the human ABC proteins. Biochim Biophys Acta. 1999, 1461: 237-262. 10.1016/S0005-2736(99)00161-3.
Annilo T, Chen ZQ, Shulenin S, Costantino J, Thomas L, Lou H, Stefanov S, Dean M: Evolution of the vertebrate ABC gene family: analysis of gene birth and death. Genomics. 2006, 88: 1-11. 10.1016/j.ygeno.2006.03.001.
Zhao Z, Fang LL, Johnsen R, Baillie DL: ATP-binding cassette protein E is involved in gene transcription and translation in Caenorhabditis elegans. Biochem Biophys Res Commun. 2004, 323: 104-111. 10.1016/j.bbrc.2004.08.068.
Dong J, Lai R, Nielsen K, Fekete CA, Qiu H, Hinnebusch AG: The essential ATP-binding cassette protein RLI1 functions in translation by promoting preinitiation complex assembly. J Biol Chem. 2004, 279: 42157-42168. 10.1074/jbc.M404502200.
Bisbal C, Martinand C, Silhol M, Lebleu B, Salehzada T: Cloning and characterization of a RNAse L inhibitor. A new component of the interferon-regulated 2-5A pathway. J Biol Chem. 1995, 270: 13308-13317. 10.1074/jbc.270.22.13308.
Marton MJ, Vazquez de Aldana CR, Qiu H, Chakraburtty K, Hinnebusch AG: Evidence that GCN1 and GCN20, translational regulators of GCN4, function on elongating ribosomes in activation of eIF2alpha kinase GCN2. Mol Cell Biol. 1997, 17: 4474-4489.
Tyzack JK, Wang X, Belsham GJ, Proud CG: ABC50 interacts with eukaryotic initiation factor 2 and associates with the ribosome in an ATP-dependent manner. J Biol Chem. 2000, 275: 34131-34139. 10.1074/jbc.M002868200.
Riordan JR, Ling V: Purification of P-glycoprotein from plasma membrane vesicles of chinese hamster ovary cell mutants with reduced colchicine permeability. J Biol Chem. 1979, 254: 12701-12705.
Gerlach JH, Endicott JA, Juranka PF, Henderson G, Sarangi F, Deuchars KL, Ling V: Homology between P-glycoprotein and a bacterial heamolysin transport protein suggests a model for multidrug resistance. Nature. 1986, 324: 485-489. 10.1038/324485a0.
Cole SP, Bhardwaj G, Gerlach JH, Mackie JE, Grant CE, Almquist KC, Stewart AJ, Kurz EU, Duncan AM, Deeley RG: Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science. 1992, 258: 1650-1654. 10.1126/science.1360704.
Doyle LA, Yang W, Abruzzo LV, Krogmann T, Gao Y, Rishi AK, Ross DD: A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc Natl Acad Sci USA. 1998, 95: 15665-15670. 10.1073/pnas.95.26.15665.
Gottesman MM, Fojo T, Bates SE: Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat Rev Cancer. 2002, 2: 48-58. 10.1038/nrc706.
Bodo A, Bakos E, Szeri F, Varadi A, Sarkadi B: The role of multidrug transporters in drug availability, metabolism and toxicity. Toxicol Lett. 2003, 140–141: 133-143. 10.1016/S0378-4274(02)00497-6.
Goldstone JV, Hamdoun A, Cole BJ, Howard-Ashby M, Nebert DW, Scally M, Dean M, Epel D, Hahn ME, Stegeman JJ: The chemical defensome: environmental sensing and response genes in the Strongylocentrotus purpuratus genome. Dev Biol. 2006, 300: 366-384. 10.1016/j.ydbio.2006.08.066.
Sheps JA, Ralph S, Zhao Z, Baillie DL, Ling V: The ABC transporter gene family of Caenorhabditis elegans has implications for the evolutionary dynamics of multidrug resistance in eukaryotes. Genome Biol. 2004, 5: R15-10.1186/gb-2004-5-3-r15.
Jones PM, George AM: Multidrug resistance in parasites: ABC transporters, P-glycoproteins and molecular modelling. Int J Parasitol. 2005, 35: 555-566. 10.1016/j.ijpara.2005.01.012.
Jungwirth H, Kuchler K: Yeast ABC transporters – a tale of sex, stress, drugs and aging. FEBS Lett. 2006, 580: 1131-1138. 10.1016/j.febslet.2005.12.050.
Rea PA: Plant ATP-binding cassette transporters. Annu Rev Plant Biol. 2007, 58: 347-375. 10.1146/annurev.arplant.57.032905.105406.
Verrier PJ, Bird D, Burla B, Dassa E, Forestier C, Geisler M, Klein M, Kolukisaoglu U, Lee Y, Martinoia E, et al: Plant ABC proteins – a unified nomenclature and updated inventory. Trends Plant Sci. 2008, 13: 151-159. 10.1016/j.tplants.2008.02.001.
Wolstenholme AJ, Fairweather I, Prichard R, von Samson-Himmelstjerna G, Sangster NC: Drug resistance in veterinary helminths. Trends Parasitol. 2004, 20: 469-476. 10.1016/j.pt.2004.07.010.
Lanning CL, Fine RL, Corcoran JJ, Ayad HM, Rose RL, Abou-Donia MB: Tobacco budworm P-glycoprotein: biochemical characterization and its involvement in pesticide resistance. Biochim Biophys Acta. 1996, 1291: 155-162.
Aurade R, Jayalakshmi SK, Sreeramulu K: Stimulatory effect of insecticides on partially purified P-glycoprotein ATPase from the resistant pest Helicoverpa armigera. Biochem Cell Biol. 2006, 84: 1045-1050. 10.1139/O06-194.
Tribble ND, Burka JF, Kibenge FS: Evidence for changes in the transcription levels of two putative P-glycoprotein genes in sea lice (Lepeophtheirus salmonis) in response to emamectin benzoate exposure. Mol Biochem Parasitol. 2007, 153: 59-65. 10.1016/j.molbiopara.2007.02.002.
Tribble ND, Burka JF, Kibenge FS, Wright GM: Identification and localization of a putative ATP-binding cassette transporter in sea lice (Lepeophtheirus salmonis) and host Atlantic salmon (Salmo salar). Parasitology. 2007, 135 (2): 243-255.
Abo-Elghar GE, Fujiyoshi P, Matsumura F: Significance of the sulfonylurea receptor (SUR) as the target of diflubenzuron in chitin synthesis inhibition in Drosophila melanogaster and Blattella germanica. Insect Biochem Mol Biol. 2004, 34: 743-752.
JGI: Joint Genome Institute. [http://www.jgi.doe.gov/Daphnia/]
Peelman F, Labeur C, Vanloo B, Roosbeek S, Devaud C, Duverger N, Denefle P, Rosier M, Vandekerckhove J, Rosseneu M: Characterization of the ABCA transporter subfamily: identification of prokaryotic and eukaryotic members, phylogeny and topology. J Mol Biol. 2003, 325: 259-274. 10.1016/S0022-2836(02)01105-1.
Wenzel JJ, Piehler A, Kaminski WE: ABC A-subclass proteins: gatekeepers of cellular phospho- and sphingolipid transport. Front Biosci. 2007, 12: 3177-3193. 10.2741/2305.
Wu CT, Budding M, Griffin MS, Croop JM: Isolation and characterization of Drosophila multidrug resistance gene homologs. Mol Cell Biol. 1991, 11: 3940-3948.
Gerrard B, Stewart C, Dean M: Analysis of Mdr50: a Drosophila P-glycoprotein/multidrug resistance gene homolog. Genomics. 1993, 17: 83-88. 10.1006/geno.1993.1286.
Begun DJ, Whitley P: Genetics of alpha-amanitin resistance in a natural population of Drosophila melanogaster. Heredity. 2000, 85 (Pt 2): 184-190. 10.1046/j.1365-2540.2000.00729.x.
Tapadia MG, Lakhotia SC: Expression of mdr49 and mdr65 multidrug resistance genes in larval tissues of Drosophila melanogaster under normal and stress conditions. Cell Stress Chaperones. 2005, 10: 7-11. 10.1379/CSC-67R.1.
Vache C, Camares O, Cardoso-Ferreira MC, Dastugue B, Creveaux I, Vaury C, Bamdad M: A potential genomic biomarker for the detection of polycyclic aromatic hydrocarbon pollutants: multidrug resistance gene 49 in Drosophila melanogaster. Environ Toxicol Chem. 2007, 26: 1418-1424. 10.1897/06-552R.1.
Abele R, Tampe R: Modulation of the antigen transport machinery TAP by friends and enemies. FEBS Lett. 2006, 580: 1156-1163. 10.1016/j.febslet.2005.11.048.
Herget M, Tampe R: Intracellular peptide transporters in human – compartmentalization of the "peptidome". Pflugers Arch. 2007, 453: 591-600. 10.1007/s00424-006-0083-4.
Zhang F, Zhang W, Liu L, Fisher CL, Hui D, Childs S, Dorovini-Zis K, Ling V: Characterization of ABCB9, an ATP binding cassette protein associated with lysosomes. J Biol Chem. 2000, 275: 23287-23294. 10.1074/jbc.M001819200.
Aleksandrov AA, Aleksandrov LA, Riordan JR: CFTR (ABCC7) is a hydrolyzable-ligand-gated channel. Pflugers Arch. 2007, 453: 693-702. 10.1007/s00424-006-0140-z.
Moreau C, Prost AL, Derand R, Vivaudou M: SUR, ABC proteins targeted by KATP channel openers. J Mol Cell Cardiol. 2005, 38: 951-963. 10.1016/j.yjmcc.2004.11.030.
Kruh GD, Belinsky MG: The MRP family of drug efflux pumps. Oncogene. 2003, 22: 7537-7552. 10.1038/sj.onc.1206953.
Conti LR, Radeke CM, Shyng SL, Vandenberg CA: Transmembrane topology of the sulfonylurea receptor SUR1. J Biol Chem. 2001, 276: 41270-41278. 10.1074/jbc.M106555200.
Deeley RG, Westlake C, Cole SP: Transmembrane transport of endo- and xenobiotics by mammalian ATP-binding cassette multidrug resistance proteins. Physiol Rev. 2006, 86: 849-899. 10.1152/physrev.00035.2005.
Akasaka T, Klinedinst S, Ocorr K, Bustamante EL, Kim SK, Bodmer R: The ATP-sensitive potassium (KATP) channel-encoded dSUR gene is required for Drosophila heart function and is regulated by tinman. Proc Natl Acad Sci USA. 2006, 103: 11999-12004. 10.1073/pnas.0603098103.
Nasonkin I, Alikasifoglu A, Ambrose C, Cahill P, Cheng M, Sarniak A, Egan M, Thomas PM: A novel sulfonylurea receptor family member expressed in the embryonic Drosophila dorsal vessel and tracheal system. J Biol Chem. 1999, 274: 29420-29425. 10.1074/jbc.274.41.29420.
Tarnay JN, Szeri F, Ilias A, Annilo T, Sung C, Le Saux O, Varadi A, Dean M, Boyd CD, Robinow S: The dMRP/CG6214 gene of Drosophila is evolutionarily and functionally related to the human multidrug resistance-associated protein family. Insect Mol Biol. 2004, 13: 539-548. 10.1111/j.0962-1075.2004.00512.x.
Grailles M, Brey PT, Roth CW: The Drosophila melanogaster multidrug-resistance protein 1 (MRP1) homolog has a novel gene structure containing two variable internal exons. Gene. 2003, 307: 41-50. 10.1016/S0378-1119(03)00455-4.
Kruh GD, Guo Y, Hopper-Borge E, Belinsky MG, Chen ZS: ABCC10, ABCC11, and ABCC12. Pflugers Arch. 2007, 453: 675-684. 10.1007/s00424-006-0114-1.
Borst P, de Wolf C, Wetering van de K: Multidrug resistance-associated proteins 3, 4, and 5. Pflugers Arch. 2007, 453: 661-673. 10.1007/s00424-006-0054-9.
Yepiskoposyan H, Egli D, Fergestad T, Selvaraj A, Treiber C, Multhaup G, Georgiev O, Schaffner W: Transcriptome response to heavy metal stress in Drosophila reveals a new zinc transporter that confers resistance to zinc. Nucleic Acids Res. 2006, 34: 4866-4877. 10.1093/nar/gkl606.
Theodoulou FL, Holdsworth M, Baker A: Peroxisomal ABC transporters. FEBS Lett. 2006, 580: 1139-1155. 10.1016/j.febslet.2005.12.095.
Petriv OI, Pilgrim DB, Rachubinski RA, Titorenko VI: RNA interference of peroxisome-related genes in C. elegans: a new model for human peroxisomal disorders. Physiol Genomics. 2002, 10: 79-91.
Berger J, Gartner J: X-linked adrenoleukodystrophy: clinical, biochemical and pathogenetic aspects. Biochim Biophys Acta. 2006, 1763: 1721-1732. 10.1016/j.bbamcr.2006.07.010.
Kerr ID: Sequence analysis of twin ATP binding cassette proteins involved in translational control, antibiotic resistance, and ribonuclease L inhibition. Biochem Biophys Res Commun. 2004, 315: 166-173. 10.1016/j.bbrc.2004.01.044.
Zhou A, Hassel BA, Silverman RH: Expression cloning of 2-5A-dependent RNAase: a uniquely regulated mediator of interferon action. Cell. 1993, 72: 753-765. 10.1016/0092-8674(93)90403-D.
Chen ZQ, Dong J, Ishimura A, Daar I, Hinnebusch AG, Dean M: The essential vertebrate ABCE1 protein interacts with eukaryotic initiation factors. J Biol Chem. 2006, 281: 7452-7457. 10.1074/jbc.M510603200.
Kusuhara H, Sugiyama Y: ATP-binding cassette, subfamily G (ABCG family). Pflugers Arch. 2007, 453: 735-744. 10.1007/s00424-006-0134-x.
Mackenzie SM, Brooker MR, Gill TR, Cox GB, Howells AJ, Ewart GD: Mutations in the white gene of Drosophila melanogaster affecting ABC transporters that determine eye colouration. Biochim Biophys Acta. 1999, 1419: 173-185. 10.1016/S0005-2736(99)00064-4.
Crouzet J, Trombik T, Fraysse AS, Boutry M: Organization and function of the plant pleiotropic drug resistance ABC transporter family. FEBS Lett. 2006, 580: 1123-1130. 10.1016/j.febslet.2005.12.043.
Rubin GM, Yandell MD, Wortman JR, Gabor Miklos GL, Nelson CR, Hariharan IK, Fortini ME, Li PW, Apweiler R, Fleischmann W, et al: Comparative genomics of the eukaryotes. Science. 2000, 287: 2204-2215. 10.1126/science.287.5461.2204.
Harrison PM, Echols N, Gerstein MB: Digging for dead genes: an analysis of the characteristics of the pseudogene population in the Caenorhabditis elegans genome. Nucleic Acids Res. 2001, 29: 818-830. 10.1093/nar/29.3.818.
Mounsey A, Bauer P, Hope IA: Evidence suggesting that a fifth of annotated Caenorhabditis elegans genes may be pseudogenes. Genome Res. 2002, 12: 770-775.
Zhao Z, Sheps JA, Ling V, Fang LL, Baillie DL: Expression analysis of ABC transporters reveals differential functions of tandemly duplicated genes in Caenorhabditis elegans. J Mol Biol. 2004, 344: 409-417. 10.1016/j.jmb.2004.09.052.
Kuwana H, Shimizu-Nishikawa K, Iwahana H, Yamamoto D: Molecular cloning and characterization of the ABC transporter expressed in Trachea (ATET) gene from Drosophila melanogaster. Biochim Biophys Acta. 1996, 1309: 47-52.
Hock T, Cottrill T, Keegan J, Garza D: The E23 early gene of Drosophila encodes an ecdysone-inducible ATP-binding cassette transporter capable of repressing ecdysone-mediated gene activation. Proc Natl Acad Sci USA. 2000, 97: 9519-9524. 10.1073/pnas.160271797.
Campbell JL, Nash HA: Volatile general anesthetics reveal a neurobiological role for the white and brown genes of Drosophila melanogaster. J Neurobiol. 2001, 49: 339-349. 10.1002/neu.10009.
Lee HG, Kim YC, Dunning JS, Han KA: Recurring ethanol exposure induces disinhibited courtship in Drosophila. PLoS ONE. 2008, 3: e1391-10.1371/journal.pone.0001391.
Kerr ID: Structure and association of ATP-binding cassette transporter nucleotide-binding domains. Biochim Biophys Acta. 2002, 1561: 47-64. 10.1016/S0304-4157(01)00008-9.
ExPASy (Expert Protein Analysis System) proteomics server of the Swiss Institute of Bioinformatics (SIB), ScanProsite facility. [http://expasy.org/tools/scanprosite/]
Tamura K, Dudley J, Nei M, Kumar S: MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol. 2007, 24: 1596-1599. 10.1093/molbev/msm092.
Acknowledgements
The sequencing and portions of the analyses were performed at the DOE Joint Genome Institute under the auspices of the U.S. Department of Energy's Office of Science, Biological and Environmental Research Program, and by the University of California, Lawrence Livermore National Laboratory under Contract No. W-7405-Eng-48, Lawrence Berkeley National Laboratory under Contract No. DE-AC02-05CH11231, Los Alamos National Laboratory under Contract No. W-7405-ENG-36 and in collaboration with the Daphnia Genomics Consortium (DGC) http://daphnia.cgb.indiana.edu. Supported in part by the Intramural Research Program of the National Institutes of Health, National Cancer Institute, Center for Cancer Research. Additional analyses were performed by wFleaBase, developed at the Genome Informatics Lab of Indiana University with support to Don Gilbert from the National Science Foundation and the National Institutes of Health. Coordination infrastructure for the DGC is provided by The Center for Genomics and Bioinformatics at Indiana University, which is supported in part by the METACyt Initiative of Indiana University, funded in part through a major grant from the Lilly Endowment, Inc. Our work benefits from, and contributes to the Daphnia Genomics Consortium.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
AS identified and annotated ABC proteins in Daphnia and wrote the manuscript. PC facilitated the bioinformatic analyses, particularly with respect to accessing and extracting database information and domain identification. MD contributed expert knowledge on ABC transporters and phylogenetic analysis, took part in the revision of this article. All authors read and approved the final manuscript.
Electronic supplementary material
12864_2008_2054_MOESM1_ESM.doc
Additional File 1: Table S1. Loci that were identified in a tBLASTn search of the Daphnia pulex genome (see Methods) but not considered further because likely representing pseudogenes (1), or consisting of short partial sequences (2), or being likely bacterial contamination (3). (DOC 54 KB)
12864_2008_2054_MOESM2_ESM.doc
Additional File 2: Table S2. Genbank accession numbers of yeast, worm, fruit fly and human ABC transporter sequences used in the phylogenetic analyses. (DOC 56 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Sturm, A., Cunningham, P. & Dean, M. The ABC transporter gene family of Daphnia pulex. BMC Genomics 10, 170 (2009). https://doi.org/10.1186/1471-2164-10-170
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2164-10-170