
Abstract
Background
Cytogenetic studies were carried out on samples of Eigenmannia virescens (Sternopygidae, Gymnotiformes) obtained from four river systems of the Eastern Amazon region (Para, Brazil).
Results
All four populations had 2n = 38, with ZZ/ZW sex chromosomes (Z, acrocentric; W, submetacentric). Constitutive heterochromatin (CH) was found at the centromeric regions of all chromosomes. The W chromosome had a heterochromatic block in the proximal region of the short arm; this CH was positive for DAPI staining, indicating that it is rich in A-T base pairs. The nucleolar organizer region (NOR) was localized to the short arm of chromosome pair 15; this result was confirmed by fluorescent in situ hybridization (FISH) with human 45S rDNA, and CMA3 staining indicated that the region is G-C rich. FISH with telomeric probes did not show any evidence of interstitial telomeric sequences (ITS).
Conclusion
Previous studies have shown that the species Eigenmannia sp. 2 and E. virescens have differentiated sex chromosomes, and diverse sex chromosome systems have been described for E. virescens specimens obtained from different Brazilian rivers. A comparative analysis of the present data and prior reports suggests that the sex chromosomes of Eigenmannia may have arisen independently in the different populations.
Similar content being viewed by others

Background
Eigenmannia Jordan & Evermann (1896) is a genus of electric fishes of the family Sternopygidae (Gymnotiformes). This genus is endemic to the main hydrographic basins of the neotropical region (e.g., Madalena, Orinoco, Amazonas and Paraná rivers), and includes species possessing an electric organ that generates wavelike electrical discharges that are used for communication and electrolocation [1–6]. Eigenmannia is presently classified into eight valid species (Table 1) that may be divided in two groups [7, 8]: Eigenmannia microstoma contains members that have large, dark bodies at sexual maturity, such as E. microstoma, E. humboldtii, E. limbata and E. nigra; while Eigenmannia virescens contains members that have two or three longitudinal lines on their bodies at sexual maturity, such as E. virescens, E. trilineata, E. vicentespelaea and E. sp. D (undescribed species). The position of Eigenmannia macrops in these groups is still undefined [8]. Despite this classification, however, Eigenmannia is considered a taxonomically confusing genus because the relative lack of morphological variation among the species makes it difficult to define diagnostic traits [7, 9, 10]. In recent years, many cytogenetic studies have been carried out on Eigenmannia samples obtained from different Brazilian basins. The diploid numbers of these samples have ranged from 2n = 28 in Eigenmannia sp. 1 [11] to 2n = 46 in Eigenmannia sp. [12]. Both ZZ/ZW and XX/XY sex chromosome systems have been described for E. virescens [13, 14], and a multiple system, X1X1X2X2/X1X2Y, was reported for Eigenmannia sp. 2 [15]. All these data are in Table 2.
Eigenmannia virescens Valenciennes (1842) appears to have one of the largest geographic distributions among the Gymnotiformes (Table 1). In the present study, we examined the karyotypes of E. virescens samples from four rivers of the Eastern Amazon region. We herein report our results, compare them with those described in the literature for species of Eigenmannia from other localities, and discuss the differences in the sex chromosomes of samples from different hydrographic basins.
Methods
Fifteen fishes of species Eigenmannia virescens (Figure 1) were collected from white-water rivers of the Eastern Amazon region during the rainy and dry seasons of 2005 to 2008 (Figure 2 and Table 3). The animals had body sizes ranging from 12 to 20 cm, and body masses ranging from 10 to 23 g. Before killing the fishes we used Benzocaine hydrochloride as an anaesthesic. Metaphase chromosomes were obtained following standard procedures [16]. The slides were analyzed using the following techniques: conventional Giemsa staining (Merck); C-banding [17]; Ag-NOR staining [18]; fluorochrome staining with CMA3 [19] and DAPI [20]; and fluorescent in situ hybridization (FISH) with biotin-labeled human 45S rDNA [21] and telomeric probes (All Telomere Probes, Oncor). The chromosomes were classified according to a previously published strategy [22].

Eigenmannia virescens collected from rivers in the Eastern Amazon region (bar: 1 cm). Source: Laboratório de Citogenética-UFPA.

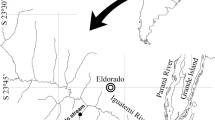
Map of the localities where the Eigenmannia virescens samples were collected. Source: Arcgis 9.1.
Results
All analyzed individuals of E. virescens (Figure 1) were found to have 2n = 38, a fundamental number (FN) of 52 for males and 53 for females (Figures 3A and 3C), and a karyotypic formula (KF) of 14m/sm+24a for males and 15m/sm+23a for females. The species was found to have a simple sex chromosome system of ZZ/ZW, where the Z is acrocentric (a) and the W is submetacentric (sm). C-banding revealed the presence of constitutive heterochromatin (CH) in the centromeric regions of all chromosomes (Figures 3B and 3D), and a particularly notable heterochromatic block was found in the proximal region of the short arm of the W chromosome (Figure 3D). The Nucleolar Organizer Region (NOR) was localized to the short arm of pair 15, which is also the location of a secondary constriction that was found to have a size heteromorphism, being less evident in some cases (Figure 3A, box) and more evident in others (Figure 3C, box). DAPI fluorescence was found in the centromeric regions of all chromosome pairs. Consistent with the C-banding results (Figures 4A and 4C), the W chromosome had a strong signal at the proximal region of the short arm (Figure 4C). The CMA3 results were consistent with our identification of the NOR (Figures 4B and 4D). Human 45S rDNA probes hybridized to the short arm of pair 15 (Figure 4E). Finally, FISH with (TTAGGG)n telomeric probes did not show any evidence of an interstitial telomeric sequence (ITS) (Figure 4F).

Karyotypes with 2n = 38, ZZ/ZW, obtained from Eigenmannia virescens collected in Benfica, Abaetetuba and Belém. (A & B) Male karyotypes stained with Giemsa and C-banding, respectively. Box: pair 15, bearing the NOR. (C & D) Female karyotypes stained with Giemsa and C-banding, respectively. Box: pair 15, bearing the NOR. Abbreviations: m-sm = metacentric-submetacentric; st-a = subtelocentric-acrocentric.

Karyotypes of Eigenmannia virescens : (A & C) DAPI banding consistent with the centromeric C-bands found in both male and female chromosomes. Note the bright staining of the W chromosome. (B & D) CMA3 banding of pair 15, consistent with our localization of the NOR. (E) FISH with rDNA 45S probes in E. virescens metaphase spreads. (F) FISH with telomeric probes in E. virescens metaphase spreads. No evidence of ITS was observed.
Discussion
We found that samples of E. virescens obtained from four localities in the Eastern Amazon region had the same karyotype (2n = 38, ZZ/ZW; KF 14m/sm+24a for males and 15m/sm+23a for females), suggesting the possibility of genetic flow among these populations. This karyotype is similar to the one described for a sample from Marajo Island [14], with the exception that the W chromosome was described as being fully heterochromatic in the Marajo sample, whereas heterochromatin was found only at the proximal region in our sample.
Previous cytogenetic studies in E. virescens from many localities have consistently reported this same diploid number (2n = 38), but there are wide variations in terms of the reported KF, chromosome sex system, and characteristics of the W chromosome (Table 2). The differences in the KF (Table 2) can be explained by the occurrence of many pericentromeric inversions, which may suggest the presence of a postzygotic mechanism for reproductive isolation. As such, each of these populations can be accepted as a valid species, and E. virescens should potentially be considered a complex of morphologically similar species. Furthermore, the description of many different karyotypes for E. virescens may arise from the population structure of this genus, whose members typically live in small populations that have little vagility [25]. These characteristics may facilitate the fixation of chromosome rearrangements.
In terms of the chromosomes themselves, the distribution of CH in E. virescens is similar to that found in most Neotropical fishes [26]. The CH block in the W chromosome may have originated from the amplification of repetitive sequences, which is not an uncommon process [14, 27]. In the Marajo Island, São Francisco River and Middle Amazonas samples, the NORs were localized to chromosome pairs 14, 17 and 16, respectively [14]. In a sample from Paraná River [13] and the present study, the NORs were localized to pair 15. These discrepancies may arise from differences in the classification and position of the chromosomes in the karyotype. Our DAPI and CMA3 staining results, which are the first such reports for this species, agree with the findings from similar studies in other Neotropical fishes [28–31]. More specifically, DAPI staining of the C-banding-positive regions showed that this CH is A-T rich, while CMA3 staining of the NOR showed that the rDNA sequences are interspersed with G-C rich sequences. The rDNA probes hybridized to different-sized regions between homologs obtained from the different individuals sampled from the four localities. This heteromorphism may be the result of differences in the copy numbers of the ribosomal genes [12, 32, 33]. Finally, we found no evidence of ITS signals. This could be due to the modification of these sequences (TTAGGG)n after a fusion event, as a consequence of telomeric loss, or the absence of chromosome rearrangements involving the telomeres. A previous study compared the identification of (TTAGGG)n sequences in salmonids under different hybridizations stringencies [34], and found that lower stringency hybridizations identified more such sequences. This result suggests that such sequences may be modified after their inclusion as an ITS because of the progressive difficult on the hybridization of the probe with the target DNA sequence. This may happened on the karyotypes here studied.
Among the Neotropical fish karyotypes studied to date, 5.9% were found to have differentiated sex chromosome systems. However, the origin of these chromosomes is still unresolved [35]. Previous cytogenetic studies in Eigenmannia identified only two other species as having differentiated sex chromosomes: Eigenmannia sp.2 (2n = 31/32, X1X1X2X2/X1X2Y) and E. virescens (2n = 38, with undifferentiated, XX/XY, and ZZ/ZW sex chromosome systems variously described for samples collected from different localities) (Table 2). According to a previous hypothesis put forth to explain the evolution of the sex chromosomes in E. virescens [14], the differentiation of their sex chromosomes first arose in the Paraná basin, where some populations lack differentiated sex chromosomes. From these undifferentiated chromosomes, amplification of the CH on one homolog of an acrocentric pair led to the development of the XX/XY system found in other populations from that basin (Table 2). Thereafter, a pericentric inversion in one of these acrocentric chromosomes generated the ZZ/ZW system found in the São Francisco River. The authors suggested that the ZZ/ZW sex system found in the Amazon basin is a posterior situation, with heterochromatinization differentiating one of the homologs. In support of this hypothesis, the authors noted that differences in HC blocks could be used to distinguish the W chromosomes of the Marajo Island populations from those of the Middle Amazon River populations.
However, we believe that an alternative hypothesis could explain the evolution of the different sex chromosomes found in Eigenmannia. Based on careful consideration of our results and those from previous reports, we suggest that the ancestral karyotype was similar to that seen in modern populations lacking differentiated sex chromosomes (such as the Mogi-Guaçu sample; Figure 5B), and the systems found in the other populations arose independently from this ancestral system (Figure 5). In Eigenmannia sp.2 (2n = 31/32, X1X1X2X2/X1X2Y), a centric fusion between two acrocentrics (pairs 6 and 11) in a male karyotype would yield the metacentric Y (neo-Y), as previously suggested [[15, 23, 24] and Figure 5A]. The other populations could be generated as follows: a) the addition of heterochromatin to the distal region of the long arm in one of the homologs would yield the XX/XY system seen in the Tiete River sample (Figure 5C); b) a pericentric inversion in one of the homologs would yield the ZZ/ZW system found in the São Francisco River sample (Figure 5D); c) the addition of heterochromatin to the short arm of an acrocentric would yield the W chromosome seen in the sample from Marajo Island (Figure 5E); d) a pericentric inversion followed by the addition of heterochromatin to the pericentromeric region of the proximal long arm of the W would generate the pattern found in the Middle Amazon River sample (Figure 5F); and e) a pericentric inversion followed by heterochromatinization in the proximal region of the short arm of the W would differentiate this chromosome to that seen in the present work, assuming that there is genetic flow among the four populations of the Eastern Amazon region studied herein (Figure 5G).

Sex chromosomes in various species of Eigenmannia. (A) Sex chromosome differentiation in Eigenmannia sp.2. (B-G) Independent sex chromosome differentiation mechanisms in populations of E. virescens from: (B) Mogi-Guaçu, (C) Tiete River, (D) São Francisco River, (E) Marajo Island, (F) Middle Amazonas River, and (G) sample from present work.
Evidence supporting our proposal for the independent origin of sex chromosomes in different species of genus Eigenmannia can be found in two previous studies using sex chromosome probes produced by microdissection from two species: the Y of Eigenmannia sp.2 (E2Y) (2n = 31/32, X1X1X2X2/X1X2Y) and the X of E. virescens (EVX) (2n = 38, XX/XY) [36, 37]. Cross-species experiments showed that these probes cross-hybridized to the autosomes but not the sex chromosomes, demonstrating that the sex chromosomes are not homologous between these two species (and therefore are likely to have arisen independently). The same thing could have happened among the ZZ/ZW system-utilizing species of E. virescens. Furthermore, the previous and present studies all found that the sex chromosomes of the various species have different characteristics (Figure 5). If we hypothesize that these forms evolved sequentially, we must assume that many evolutionary steps have occurred along the way. Since each studied taxon was found to have a different KF, we must also suppose that each karyotype gave rise to next, which does not appear likely. Collectively, these lines of evidence would seem to support our hypothesis that the sex chromosomes in different species of genus Eigenmannia may have independent origins.
Conclusion
We herein report that samples of E. virescens obtained from four localities in the Eastern Amazon region had the same karyotype and possessed differentiated sex chromosomes (ZZ/ZW). Previous studies showed that Eigenmannia sp. 2 and E. virescens have differentiated sex chromosomes, and diverse sex chromosome systems have been described for E. virescens from different Brazilian rivers. A comparative analysis of our present data and the previous reports led us to suggest that the sex chromosomes of different species of Eigenmannia may have arisen independently.
References
Albert JS: Family Sternopygidae. Checklist of the freshwater fish of South and Central America. Edited by: Reis RE, Kullander SO, Ferraris CJ Jr. 2003, Edipuers, Porto Alegre, 493-497.
Alves-Gomes JA, Guillermo O, Haygood M, Heiligenberg W, Meyer A: Phylogenetic analysis of the South American electric fish (order Gymnotiformes) and the evolution of their electrogenic system: a synthesis based on morphology, electrophysiology, and mitochondrial sequence data. Molecular Biology and Evolution. 1995, 2: 298-318.
Alves-Gomes JA: The phylogenetic position of the South American electric fish genera Sternopygus and Archolaemus (Ostariophysi: Gymnotiformes) according to 12S and 16S mitocondrial DNA sequences. Phylogeny and classification of Neotropical fishes. Edited by: Malabarba LR, Reis RE, Vari RP, Lucena ZMS, Lucena CAS. 1998, Porto Alegre, Edipucrs, 447-460.
Crampton WGR, Albert JS: Evolution of electric signal diversity in the gymnotiform fish. Fish communication. Edited by: Collin SP, Kpoor BG, Ladich F, Moller P. 2005, Science Publisher Inc, New York
Kramer B: Mechanisms of signal analysis in Eigenmannia (Gymnotiformes): The jamming avoidance response and communication. Biology of tropical fishes. Edited by: Val AL, Almeida-Val VMF. 1999, INPA, Manaus, Chapter 4: 41-61.
Mago-Leccia F: Los peces de la família Sternopygidae de Venezuela. Acta Cientifica Venezolana. 1978, 29: 1-90.
Albert JS: Species diversity and phylogenetics systematics of American knifefish (Gymnotiformes, Teleostei). Misc Publi Mus Zool University of Michigan. 2001, 190: 1-129.
Bichuette ME, Trajano E: Morphology and distribution of the knifefish Eigenmannia vicentespelea Triques, 1996 (Gymnotiformes: Sternopygidae) from Central Brazil, with an expanded diagnosis and comments on subterranean evolution. Neotropical Ichtyology. 2006, 4: 99-105.
Mago-Leccia F: Electric fishes of the continental waters of America. Biblioteca de la Academia de Ciencias Fisicas, Matematicas, y Naturals, Caracas, Venezuela. 1994, 29: 1-206.
Triques ML: Filogenia dos gêneros de Gymnotiformes (Actinopterygii, Ostariophysi), com base em caracteres esqueléticos. Comunicações do Museu de Ciências PUCRS série zoológica, Porto Alegre. 1993, 6: 85-130.
Almeida-Toledo LF, Stocker AJ, Foresti F, Toledo-Filho AS: Fluorescence in situ hibridization with rDNA probes on chromosomes of two nucleolus organizer region phenotypes of a species of Eigenmannia (Pisces, Gymnotoidei, Sternpygidae). Chromosome Research. 1996, 4: 301-305. 10.1007/BF02263681.
Almeida-Toledo LF, Foresti F, De Almeida-Toledo Filho S: Spontaneous triploidy and NOR activity in Eigenmannia sp. (Pisces, Sternopygidae) from Amazon basin. Genetica. 1985, 66: 85-88. 10.1007/BF00139713.
Almeida-Toledo LF, Foresti F, Péquignot EV, Daniel-Silva MFZ: XX:XY sex chromosome system with X heterocromatinization: an early stage of sex chromosome differentiation um the Neotropic electric eel Eigenmannia virescens. Cytogenetics and Cell Genetics. 2001, 95: 73-78. 10.1159/000057020.
Almeida-Toledo LF, Daniel-Silva MFZ, Moysés CB, Fonteles SBA, Lopes CE, Akama A, Foresti F: Chromosome evolution in fish: sex chromosome variability in Eigenmannia virescens (Gymnotiformes, Sternopygidae). Cytogenetic and Genoma Research. 2002, 99: 164-169. 10.1159/000071589.
Almeida-Toledo LF, Foresti F, Daniel MFZ, Toledo-Filho S: Sex chromosome evolution in fish: the formation of the neo-Y chromosome in Eigenmannia (Gymnotiformes). Chromosoma. 2000, 109: 197-200. 10.1007/s004120050428.
Bertollo LAC, Takashi CS, Moreira-Filho O: Cytotaxonomic considerations on Hoplias lacerdae (Pisces, Erythrinidae). Brazilian Journal of Genetics. 1978, 2: 103-120.
Sumner AT: A simple technique for demonstrating centromeric heterochromatin. Experimental Cell Research. 1972, 75: 304-306. 10.1016/0014-4827(72)90558-7.
Howell WM, Black DA: Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia. 1980, 36: 1014-1015. 10.1007/BF01953855.
Schweizer D: Simultaneous fluorescent staining of R bands and specific heterochromatic regions (DA/DAPI Bands) in human chromosomes. Cytogenetics and Cell Genetics. 1980, 27: 190-193. 10.1159/000131482.
Pieczarka JC, Nagamachi CY, Souza ACP, Milhomem SSR, Castro RR, Nascimento AL: An adaptation to DAPI-Banding to fishes chromosomes. Caryologia. 2006, 59: 43-46.
Daniels LM, Delany ME: Molecular and cytogenetic organization of the 5S ribosomal DNA array in chicken (Gallus gallus). Chromosome Research. 2003, 11: 305-317. 10.1023/A:1024008522122.
Levan A, Fredga K, Sandberg HA: Nomenclature for centromeric position on chromosomes. Hereditas. 1964, 52: 201-220. 10.1111/j.1601-5223.1964.tb01953.x.
Almeida-Toledo LF, Viegas-Péquignot E, Foresti F, Toledo Filho AS, Dutrillaux B: BrdU replication patterns demonstrating chromosome homoeologies in two fish species, genus Eigenmannia. Cytogenetics and Cell Genetics. 1988, 48: 117-120. 10.1159/000132603.
Almeida-Toledo LF, Foresti H, Almeida-Toledo Filho S: Complex sex chromosome system in Eigenmannia sp. (Pisces, Gymnotiformes). Genetica. 1984, 64: 165-169. 10.1007/BF00115340.
Moysés CB, Mockford S, Almeida-Toledo LF, Wright JM: Nine polymorphic microsatellite loci in the Neotropical electric eel Eigenmannia (Teleostei: Gymnotiformes). Molecular Ecology Notes. 2005, 5: 7-9. 10.1111/j.1471-8286.2004.00803.x.
Galetti PM, Martins C: Contribuição da hibridização in situ para o conhecimento dos cromossomos de peixes. FISH: Conceitos e aplicações na citogenética. Edited by: Guerra M. 2004, Ribeirão Preto: Sociedade Brasileira de Genética, 61-88.
Li WH: Fundamentals of Molecular Evolution. 1991, Sinaur, Sunderland, 1: 216-219. 1
Milhomem SSR, Pieczarka JC, Crampton WGR, Souza ACP, Carvalho JR, Nagamachi CY: Differences in karyotype between two sympatric of Gymnotus (Gymnotiformes: Gymnotidae) from the eastern amazon of Brazil. Zootaxa. 2007, 1397: 55-62.
Milhomem SSR, Pieczarka JC, Crampton WGR, Silva DS, Souza ACP, Carvalho JR, Nagamachi CY: Chromosomal evidence for a cryptic species in the Gymnotus carapo species-complex (Gymnotiformes, Gymnotidae). BMC Genetics. 2008, 9: 75-10.1186/1471-2156-9-75. doi:10.1186/1471-2156-9-75
Nascimento AL, Souza ACP, Feldberg E, Carvalho JR, Barros RMS, Pieczarka JC, Nagamachi CY: Cytogenetic analysis on Pterophyllum scalare (Perciformes, Cichlidae) from Jari River, Pará state. Caryologia. 2006, 59: 138-143.
Silva DS, Milhomem SSR, Souza ACP, Pieczarka JC, Nagamachi CY: A conserved karyotype of Sternopygus macrurus (Sternopygidae, Gymnotyformes) in the Amazon region: Differences from other hydrographic basins suggest cryptic speciation. Micron. 2008, 39: 1251-1254. 10.1016/j.micron.2008.04.001.
Foresti F, Almeida-Toledo LF, Toledo-Filho SA: Polymorphic nature of nucleolus organizer regions in fishes. Cytogenetics and Cell Genetics. 1981, 31: 137-144. 10.1159/000131639.
Oliveira C, Nirchio M, Granado A, Levy S: Karyotypic characterization of Prochilodus mariae, Semaprochilodus kneri and S. laticeps (Teleostei: Prochilodontidae) from Caicara del Orenoco, Venezuela. Neotropical Ichthyology. 2003, 1: 47-52. 10.1590/S1679-62252003000100005.
Abuín M, Martínez P, Sánchez L: Localization of the repetitive telomeric sequence (TTAGGG)n in four salmonid species. Genome. 1996, 39: 1035-1038. 10.1139/g96-129.
Oliveira C, Foresti F, Hulsdorf AWS: Genetics of Neotropical fish: from chromosomes to populations. Fish Physiology and Biochemistry. 2009, 35: 81-100. 10.1007/s10695-008-9250-1. Doi: 10.1007/s10695-008-9250-1
Henning F, Trifonov V, Almeida-Toledo LF: Use of chromosome microdissection in fish molecular cytogenetics. Genetics and Molecular Biology. 2008, 31: 279-283. 10.1590/S1415-47572008000200022.
Henning F, Trifonov V, Ferguson-Smith MA, Almeida-Toledo LF: Non-homologous sex chromosomes in two species of the genus Eigenmannia (Teleostei: Gymnotiformes). Cytogenetics and Genome Research. 2008, 121: 55-58. 10.1159/000124382.
Acknowledgements
This research was financially supported by CNPq, CAPES, FAPESPA and UFPA. JCP and CYN are CNPq Researchers, level 1. DSS received a CAPES Masters Scholarship on Genetics and Molecular Biology. SSRM received a CNPq Doctors Scholarship on Genetics and Molecular Biology. We thank the Programa de Pós-Graduação em Genética e Biologia Molecular for supporting this study; researchers Ms. Jaime Ribeiro Jr. and Ms. Augusto Cesar Paes de Souza for their help with field work and species identification; and geologist João Marinho Milhomem Neto for help in mapping where the sample collections took place. The field collections were authorized by the IBAMA (Instituto Brasileiro de Meio Ambiente). Finally, we are grateful to Dr. Wolmar Benjamin Wosiacki (Curator of Fishes, Museu Paraense Emilio Goeldi) for vouchering the specimens and adding them to the museum's collection.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
DSS collected the samples, collaborated on all cytogenetic procedures, undertook the bibliographic review, and coordinated the writing of this paper. JCP helped conceive the study and participated in developing the laboratory techniques, cytogenetic analyses and writing. SSRM participated in collecting the specimens and developing the laboratory techniques. CYN coordinated the study, helped develop the laboratory techniques and cytogenetic analyses, and reviewed the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Silva, D.S., Milhomem, S.S., Pieczarka, J.C. et al. Cytogenetic studies in Eigenmannia virescens (Sternopygidae, Gymnotiformes) and new inferences on the origin of sex chromosomes in the Eigenmannia genus. BMC Genet 10, 74 (2009). https://doi.org/10.1186/1471-2156-10-74
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2156-10-74