
Abstract
Background
Studies from our laboratory have shown that human alveolar macrophages (AM) and bronchial epithelial cells (HBEC) exposed to ambient particles (PM10) in vitro increase their production of inflammatory mediators and that supernatants from PM10-exposed cells shorten the transit time of monocytes through the bone marrow and promote their release into the circulation.
Methods
The present study concerns co-culture of AM and HBEC exposed to PM10 (EHC-93) and the production of mediators involved in monocyte kinetics measured at both the mRNA and protein levels. The experiments were also designed to determine the role of the adhesive interaction between these cells via the intercellular adhesion molecule (ICAM)-1 in the production of these mediators.
Results
AM/HBEC co-cultures exposed to 100 μg/ml of PM10 for 2 or 24 h increased their levels of granulocyte-macrophage colony-stimulating factor (GM-CSF), M-CSF, macrophage inflammatory protein (MIP)-1β, monocyte chemotactic protein (MCP)-1, interleukin (IL)-6 and ICAM-1 mRNA, compared to exposed AM or HBEC mono-cultures, or control non-exposed co-cultures. The levels of GM-CSF, M-CSF, MIP-1β and IL-6 increased in co-cultured supernatants collected after 24 h exposure compared to control cells (p < 0.05). There was synergy between AM and HBEC in the production of GM-CSF, MIP-1β and IL-6. But neither pretreatment of HBEC with blocking antibodies against ICAM-1 nor cross-linking of ICAM-1 on HBEC blocked the PM10-induced increase in co-culture mRNA expression.
Conclusion
We conclude that an ICAM-1 independent interaction between AM and HBEC, lung cells that process inhaled particles, increases the production and release of mediators that enhance bone marrow turnover of monocytes and their recruitment into tissues. We speculate that this interaction amplifies PM10-induced lung inflammation and contributes to both the pulmonary and systemic morbidity associated with exposure to air pollution.
Similar content being viewed by others


Background
Exposure to ambient particulate matter with a diameter of less than 10 μm (PM10) is strongly associated with increased morbidity and mortality, particularly in subjects with pre-existing pulmonary and cardiovascular diseases [1, 2]. This increase in mortality induced by PM10 exposure was present even when adjusted for the other major risk factors such as cigarette smoking [1]. A recent report [3] has shown that environmentally relevant concentrations of PM2.5 induced airway inflammation even in healthy subjects with a selective influx of monocytes.
Although the biological mechanisms are still unclear, PM10 are known to stimulate the production of reactive oxygen species and inflammatory mediators by alveolar macrophages (AM) [4–7] and epithelial [7–10] and other lung cells [11]. When AM and airway epithelial cells are directly exposed to inhaled atmospheric particles these small particles are phagocytized by both cells [10, 12]. Both cell types can synthesize a variety of pro-inflammatory cytokines that induce airway inflammation and contribute to the airway lesions in asthma and chronic obstructive pulmonary diseases [9]. In vitro, AM and lung epithelial cells interact in response to PM10 and this interaction has been implicated in amplifying their mediator production [7, 13]. Studies from our laboratory have shown that the PM10(EHC-93)-induced interaction of human AM and bronchial epithelial cells (HBEC) enhances the synthesis and release of a variety of pro-inflammatory cytokines and that supernatants from these co-cultures instilled into rabbit lungs induces a systemic inflammatory response [13].
We recently showed that deposition of PM10 (EHC-93 and inert carbon particles) in the lung shortened the transit time of monocytes through the bone marrow and enhanced their release into the circulation [14, 15]. Furthermore, we also showed that monocytes are the predominant inflammatory cells that accumulate in the alveoli following repeated PM10 exposure [16]. The present study was designed to determine whether, and if so, which interactions between AM and HBEC (AM/HBEC co-cultures) amplify the response to PM10 exposure, especially the synthesis of inflammatory mediators that enhance bone marrow turnover of monocytes and their recruitment into the lung. We used primary cultures of HBEC and human AM freshly isolated from lobectomy or pneumonectomy specimens and measured the expression of inflammatory mediators relevant to monocyte kinetics. We further evaluated the potential role of the intercellular adhesion molecule (ICAM)-1 in the production of mediators by AM/HBEC co-cultures exposed to PM10.
Methods
Urban air particles (PM10)
PM10 particles were collected in an urban environment (EHC-93) and obtained from the Environmental Health Directorate, Health Canada, Ottawa, Ontario. A detailed analysis of the EHC-93 has been presented elsewhere [17]. Particles were suspended at a concentration of 1 mg/ml in hydrocortisone-free supplemented bronchial epithelial cell growth medium (BEGM; Clonetics, San Diego, CA) and sonicated 3 times for 1 min each at maximal power on a Vibra Cell VC-50 sonicator (Sonics and Materials Inc., Danbury, CT) prior to adding to the cells. The endotoxin content of the PM10 suspension of 100 μg/ml was 6.4 ± 1.8 EU/ml or less than 3.0 ng/ml [10, 13]. This dose of LPS has been shown not to activate either AM or lung epithelial cells to produce cytokines [10].
Isolation of HBEC and human AM
Bronchial tissue and broncho-alveolar lavage (BAL) fluid was obtained from a total of ten patients who underwent lobectomy or pneumonectomy for small peripheral nodules at St. Paul's Hospital, Vancouver. Informed consent was obtained from all subjects and these studies were approved by the Human Ethics Committee of the University of British Columbia. All subjects were current smokers and were asked to abstain from smoking for 6 weeks prior to the operation. Their mean age was 67.2 yr (range 56–74 yr) (6 women and 4 men). Primary HBEC were isolated from bronchial tissues according to a previously described procedure [10]. In brief, pieces of excised human bronchial tissue approximately 1 cm long were incubated at 4°C for 24 h with 0.1% protease (Type14; Sigma) solution prepared in BEGM containing Fungizone (1 μg/ml; GIBCO BRL, Gaithersburg, MD). The epithelial cells were harvested, washed with BEGM with added antibiotics(100 U/ml of penicillin and 100 μ/gml of streptomycin; Sigma)and Fungizone, and cultured in a 25-cm2 cell culture flask until 80 to 90% confluent. Then the cells were trypsinized and placed in 100-mm cell culture dishes and cultured in BEGM. Light microscopy showed that 95% of the isolated cells had features of bronchial epithelial cells, that is they formed a monolayer of ciliated cells. Also, by trypan blue exclusion, >95% of these cells were viable. Human AM were harvested from BAL fluid obtained from lung segments or lobes that were free of the tumor using a method previously described in detail [7, 13]. The BAL fluid cells were >90% viable (trypan blue exclusion method) and consisted of 90–95% AM (as assessed by Wrights-Giemsa stain) and less than 2% neutrophils. AM mono-cultures and AM/HBEC co-cultures were suspended in BEGM. BEGM used throughout this study was without hydrocortisone.
Exposure of cells to PM10
Primary HBEC from the third or fourth passage of cells from each patient were cultured to 90–100 % confluence in 100-mm cell culture dishes (approximately 2.5–3.0 × 106 cells/dish) then exposed for 2 and 24 h to fresh stock suspensions of 100 μg/ml PM10 (EHC-93) prepared in BEGM.
AM (1.0 × 107) from each patient were placed in 100-mm cell culture dishes and allowed to adhere to the plastic dish for 30 min in humidified incubator (5% CO2 at 37°C). The non-adherent cells less than 1.0 × 106) were then removed by rinsing twice with BEGM and adherent AM (>98% AM) were incubated in 10 ml of BEGM with or without 100 μg/ml of PM10 for 2 and 24 h.
In co-culture experiments, freshly prepared AM (5.0 × 106) were directly placed on the confluent HBEC monolayers which were grown in 100-mm cell culture dishes. The AM were allowed to adhere to HBEC and the non-adherent cells were removed by washing twice with BEGM. The AM/HBEC co-cultured cells were incubated in 10 ml of BEGM with or without 100 μg/ml of PM10 for 2 and 24 h. Cell viability was determined following the 24 h PM10 exposure in all experiments using the trypan blue exclusion method.
RNase protection assay (RPA)
After 2 or 24 h treatment, total RNA was isolated from the cells using a single-step phenol/chloroform extraction procedure (Trizol, Life Technologies, Inc., Grand Island, NY). The levels of inflammatory mediator mRNA were determined using the RiboQuant™ multi-probe system (PharMingen, San Diego, CA) following the instructions of the supplier. Two customized template sets were used that included mRNAs of the following inflammatory mediators: human regulated on activation, normal T-cells expressed and secreted (RANTES), macrophage inflammatory protein (MIP)-1β, granulocyte-macrophage colony-stimulating factor (GM-CSF), M-CSF, monocyte chemotactic protein (MCP)-1, interleukin (IL)-6 and leukemia inhibitory factor (LIF). Human ICAM-1 mRNA was determined using a separate template set. Internal controls included mRNAs of the ribosomal protein L32 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). In brief, 10 μg of total cellular RNA was hybridized overnight to the [α-32P] UTP-labeled riboprobes which had been synthesized from the supplied template sets. Single-stranded RNA and free probe remaining after hybridization were digested by a mixture of RNase A and T1. The protected RNA was then phenolized, precipitated, and analyzed on a 5% denaturing polyacrylamide gel. Following electrophoresis, the gel was dried under vacuum and subjected to autoradiography. The quantity of protected labeled RNA was determined using densitometry and the NIH image 1.63 software (National Institutes of Health, Bethesda, MD). Results were normalized to the expression of the internal control, GAPDH. For the densitometric analysis each RPA was repeated four to six times.
ELISA measurements
Cell culture supernatants were collected 24 h after addition of 100 μg/ml of PM10 suspension, centrifuged, filtered through a syringe filter with pore size of 0.22 μm (Corning, Cambridge, MA) to eliminate as much as possible any remaining particles and stored at -80°C until use. MIP-1β, GM-CSF, M-CSF, MCP-1 and IL-6 levels were measured by the Cytokine Core Laboratory (Baltimore, MD) using an ELISA based on a biotin-strepavidin-peroxidase detection system as previously described [10]. All measurements were done in triplicate and values corrected for the number of AM used in each experiment are reported as the means of five experiments.
Immunocytochemistry
To demonstrate cell surface ICAM-1 (CD54) expression on HBEC and CD11b on AM, cells were placed or grown on coverslips in 6-well plates and incubated for 2 or 24 h with 100 μg/ml of PM10. Cells were fixed with 1% paraformaldehyde for 10 min and immunocytochemistry was performed by the alkaline phosphatase anti-alkaline phosphatase method using mouse anti-human CD54 monoclonal antibody (Immunotech, Marseille, France) and mouse anti-human CD11b monoclonal antibody (DAKO, Copenhagen, Denmark) to identify cell surface expression of ICAM-1 and CD11b.
Cell adhesion blockers and ICAM-1 cross-linking
In experiments testing whether anti-CD54 and anti-CD11b block mediator production by co-cultured AM/HBEC, HBEC and AM were preincubated for 1 h before PM10 exposure with control IgG F(ab')2 fragments (2 μg/ml; Jackson ImmunoResearch Laboratories, PA), mouse anti-human monoclonal CD54 F(ab')2 fragments, and/or monoclonal CD11b F(ab')2 fragments (1 μg/ml, respectively; Caltag Laboratories, CA). Cells were then co-cultured and exposed to PM10 for 24 h in the presence of the blocking antibodies before analysis by RPA. To determine whether ligand binding to CD54 on HBEC in of itself contributes to the enhanced mediator response of these cells to PM10 stimulation cross-linking antibodies to CD54 were used to simulate this possibility. We used previously reported methods of cross-linking CD54 which resulted in intracellular signaling [18, 19]. After 2 h of exposure to PM10, HBEC were incubated for 1 h with 1 μg/ml mouse anti-human CD54 or 1 μg/ml control mouse non-specific IgG (DAKO). Cells were washed and then incubated for 4 h with 10 μg/ml rabbit anti-mouse IgG (DAKO) to cross-link the bound anti-CD54 and mRNA mediator expression was assessed as above.
Statistical Analysis
Data are expressed as mean values ± SE. The minimum number of replicates for all measurement was at least three. For RPA and ELISA, differences between matched pairs (control versus PM10 treated) were compared by Wilcoxon signed ranked test. To compare mediator production by co-cultures to that by AM plus HBEC mono-cultures, we used the Mann-Whitney U test. Differences between multiple groups were compared by one-way analysis of variance (ANOVA). The post hoc test for multiple comparisons was the Dunnett's test. Significance was assumed at p < 0.05.
Results
AM/HBEC co-cultures and PM10
We previously showed that the majority of AM and HBEC were in contact with each other in our co-culture system [13]. Both cells internalized PM10 particles with many cells containing more than one particle. The 100 μg/ml concentration of PM10 used throughout this experiment was not toxic to either AM or HBEC and >90% of cells were viable after 24 h exposure as assessed by the trypan blue exclusion method.
Expression of mRNA induced by PM10
Representative autoradiographs of mRNA expression by AM or HBEC mono-cultures and AM/HBEC co-cultures after 2 and 24 h incubation in medium alone (control) or a 100 μg/ml of PM10 suspension (PM10) are shown in Figure 1. Because the RPA kit is not provided with an internal control to account for variation between autoradiographs of different pairs of control versus PM10 treated cells such as those shown in Figure 1 and due to that fact that cells from a single but different patient are represented in each different pair, as documented by the differences in intensity of the control L32 band(s) compared to that of the corresponding GAPDH bands as well as differences in the L32 banding pattern (Fig. 1), expected large variations in densitometric data between corresponding pairs of autoradiographs were found. Despite these variations the compiled densitometric analyses of these autoradiographs yielded statistically significant results. However, because of the unavoidable variations, the compiled densitometric results for a few mediators differed from that depicted in the representative autoradiographs. In Figure 1 mRNA expression of the inflammatory mediators of interest, RANTES, MIP-1β, GM-CSF, M-CSF, MCP-1, IL-6 and LIF, was not altered after 2 h of PM10 exposure of neither AM nor HBEC mono-cultures and this result was confirmed after densitometric analysis (n = 4, data not shown). Only the expression of ICAM-1 mRNA by HBEC at this time-point appears to be marginally increased (Fig. 1) but after densitometric analysis this change was not found to be significant (n = 4, data not shown). In contrast to the results from the mono-cultures, PM10 exposure for 2 h of co-cultured AM/HBEC increased MIP-1β, GM-CSF, M-CSF, IL-6, LIF and ICAM-1 mRNA expression (Fig. 1) and, of these, densitometric analysis of six such RPA experiments confirmed that increases in MIP-1β, GM-CSF, IL-6, and ICAM-1 were significant (Fig. 2A), as well as that of MCP-1 (Fig. 2A) which was not detected in the representative autoradiograph (Fig. 1).

RNase protection assay of mRNA expression by AM and HBEC. Representative autoradiographs of RNase protection assays (RPAs) showing mediator expression by AM/HBEC co-cultures, AM mono-cultures and HBEC mono-cultures after 2 and 24 h incubation in medium alone (control) or a 100 μg/ml of PM10 suspension (PM10). After 24 h exposure AM showed increased expression of LIF and ICAM-1 mRNA. Expression of GM-CSF, LIF and ICAM-1 mRNA by HBEC was increased by 24 h PM10 stimulation compared to their respective controls. MIP-1β, GM-CSF, M-CSF, MCP-1, IL-6, LIF and ICAM-1 mRNA expression by AM/HBEC co-cultures was increased 2 and/or 24 h after incubation with PM10 compared to control. L32 and GAPDH were used as controls for lane loading.

Densitometric analysis of bands on RPAs. (A): the density of the bands representing the mediator mRNAs in AM/HBEC co-cultures on autoradiographs such as that shown in Figure 1A was compared to that of the GAPDH mRNA band in the same lane and the resulting ratio (PM10; black bars) is shown as the percentage change from control values (white bars). The mean densitometric value confirmed that the mRNA levels of MIP-1β, GM-CSF, MCP-1, IL-6 and ICAM-1 at 2 h and those of MIP-1β, GM-CSF, M-CSF and ICAM-1 after 24 h exposure were significantly higher than control values. Values are means ± SE of six experiments representing the AM/HBEC co-culture group. (B): the mean densitometric value confirmed that the mRNA levels of GM-CSF, LIF and ICAM-1 at 24 h exposure were significantly higher than control values. Values are means ± SE of four experiments representing the HBEC mono-culture group. *p < 0.05 compared with control.
After 24 h exposure to PM10 the representative autoradiographs showed that increases in mRNA expression by AM mono-cultures were restricted to that of LIF and ICAM-1 (Fig. 1) but this was not confirmed after statistical analysis of the densitometric results (n = 4, data not shown). In contrast, the increases in GM-CSF, LIF and ICAM-1 by HBEC mono-cultures (Fig. 1) were found to be statistically significant (p < 0.05 and n = 4, respectively)(Fig. 2B). Co-cultures exposed to PM10 at this time-point showed strong increases in MIP-1β, GM-CSF, M-CSF, IL-6 and ICAM-1 mRNA and minor increases in those of MCP-1 and LIF (Fig. 1). Except for IL-6, the strong increases were confirmed by the densitometric analysis of six RPA experiments (Fig. 2A).
Mediator production induced by PM10
Figure 3 shows the GM-CSF, IL-6, MIP-1β, MCP-1 and M-CSF protein levels in supernatants of AM/HBEC co-cultures, AM mono-cultures and HBEC mono-cultures incubated for 24 h with medium alone (control) or with 100 μg/ml of PM10. GM-CSF, IL-6, MIP-1β and M-CSF production by AM/HBEC co-cultures stimulated by PM10 were significantly increased compared to control levels. GM-CSF and IL-6 production by AM mono-cultures stimulated with PM10 suspension increased significantly compared to controls. MIP-1β production by HBEC mono-culture stimulated by PM10 were significantly increased over control levels.

Mediator protein levels in supernatants of AM and HBEC. GM-CSF, IL-6, MIP-1β, MCP-1, and M-CSF protein levels in supernatants of AM mono-cultures, HBEC mono-cultures and AM/HBEC co-cultures incubated for 24 h with medium alone (control; white bars) or 100 μg/ml of PM10 (black bars). GM-CSF and IL-6 production by AM mono-cultures and AM/HBEC co-cultures stimulated by PM10 increased significantly compared to controls. Exposure to PM10 also increased MIP-1β production by HBEC mono-cultures and AM/HBEC co-cultures and M-CSF production by AM/HBEC co-cultures. The GM-CSF, IL-6 and MIP-1β produced by exposed AM/HBEC co-cultures significantly exceeded the sum of those produced by AM and HBEC mono-cultures exposed separately. Values are means ± SE of five experiments. * p < 0.05 compared with control. † p < 0.05 for exposed AM/HBEC co-cultures compared to the sum of the exposed HBEC and AM mono-cultures.
The GM-CSF, IL-6 and MIP-1β produced by AM/HBEC co-cultures in response to PM10 stimulation were more than the sum of the respective mediator produced by PM10 exposed AM and HBEC mono-cultures alone suggesting a synergistic effect in production of these cytokines (p < 0.05). This synergistic effect was not seen in the production of M-CSF. MCP-1 production was not significantly increased by PM10 in either mono-cultures or AM/HBEC co-cultures but its expression by AM tended to be decreased by co-culturing (Fig. 3).
Expression of ICAM-1 induced by PM10
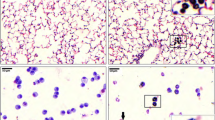
Figure 4 shows immunocytochemically stained CD54 on HBEC (to identify ICAM-1) and CD11b on AM. In the absence of PM10, HBEC express low levels of ICAM-1 on their cell surface (Fig. 4A). After stimulation with 100 μg/ml of PM10 for 24 h many more cells stained positively for ICAM-1 and their intensity of staining was increased (Fig. 4B). Most AM expressed surface CD11b and this expression was unaffected by 2 and 24 h stimulation with PM10 (Fig. 4C, D and data not shown).

Surface expression of ICAM-1 on HBEC and CD11b on AM. Photomicrographs of primary cultured HBEC and human AM on coverslips. Immunocytochemistry was performed using mouse anti-human CD54 monoclonal antibody on HBEC and mouse anti-human CD11b monoclonal antibody on AM. In the absence of PM10 stimulation HBEC rarely expressed CD54 (A). After stimulation with 100 μg/ml of PM10 for 24 h the majority of cells stained positively (arrows, pink cells) for CD54 (B). Expression of surface CD11b on AM (C) was unaffected by 2 h stimulation with PM10 (D). The scale bars represent 20 μm.
ICAM-1 and PM10-induced mediator production by AM/HBEC co-cultures
To determine the role of β2-integrin/ICAM-1 interaction in mediator production by AM/HBEC co-cultures, AM and HBEC were incubated with inhibitors of these adhesion molecules before PM10 exposure. Representative autoradiographs of mRNA expression by such AM/HBEC co-cultures after 24 h incubation in a 100 μg/ml of PM10 suspension are shown in Figure 5A. They include pretreatment of neither cell before co-culture, of only AM with control IgG or anti-CD11b antibody, of only HBEC with control IgG or anti-CD54 antibody, and of both cell types with both antibodies. The increased mRNA expression in PM10-stimulated AM/HBEC co-cultures was not affected by any of the pretreatments with these antibodies. In addition, as shown in Figure 5B, CD54 cross-linking itself in the absence of AM did not induce mediator expression in HBEC exposed to PM10.

AM/HBEC co-culture responses after pretreatment with cell adhesion blockers. (A): autoradiographs from RNase protection assay of mediator mRNA expression by AM/HBEC co-cultures pretreated before 24 h incubation in a 100 μg/ml of PM10 suspension including no pretreatment (no treatment) before co-culture, AM pretreated with control IgG (AM-IgG), HBEC with control IgG (HBEC-IgG), AM with anti-CD11b antibody (AM-CD11b), HBEC with anti-CD54 antibody (HBEC-CD54) and both cell types with respective antibodies (both antibodies). The mRNA expression in PM10-exposed AM/HBEC co-cultures was not affected by any pretreatments with these antibodies. (B): in the absence of AM, pretreatment of HBEC to cross-link CD54 with antibody followed by 2 h exposure to PM10 (PM10-CD54) did not alter mediator expression compared with HBEC pretreated with control IgG (PM10-IgG) and non-pretreated HBEC (control).
Discussion
AM and lung epithelial cells play a key role in processing inhaled particulate matter. In the present study we confirmed that exposing co-cultures of human AM and HBEC atmospheric particles to for 2 hr increased mRNA expression of GM-CSF, MCP-1 and IL-6 [13]. The current addition of mRNAs, that of M-CSF and MIP-1β, to this list of these mediators involved in the marrow production, mobilization and recruitment of monocytes that are increased in response to PM10 exposure reinforces the hypothesis that exposure of the lung to environmental pollutants can stimulate a systemic inflammatory response [4]. Besides these bone marrow oriented mediators, mRNA expression of ICAM-1, an adhesion molecule potentially involved in an interaction between AM and HBEC to amplify marrow-related mediator expression, was increased. Another important hitherto unreported finding, that of sustained increased expression of many of these mediator mRNAs, MIP-1β, GM-CSF, M-CSF, and ICAM-1, over 24 h of exposure, supports the robust increase in the expression of the corresponding mediator proteins that we observed. These included MIP-1β, M-CSF, ICAM-1 as well as the previously reported GM-CSF and IL-6 [13]. Furthermore, the synergistic increases in GM-CSF, IL-6 and MIP-1β secretion by the co-cultures compared to the sum of the mono-cultures in response to PM10 exposure indicate an interaction between these cells with ICAM-1 possibly contributing to this interaction.
IL-6, the hematopoietic growth factors GM-CSF and M-CSF, and the C-C chemokine MIP-1 are important mediators in the production and mobilization of monocytes from the bone marrow [20–22]. IL-6 is considered an important multifunctional cytokine involved in the regulation of a variety of cellular responses, including being a permissive factor for monocytic colony formation by human hematopoietic progenitor cells in combination with GM-CSF [23]. Monocytes recruited into the lung play a critical important role in clearing foreign material such as particles from the lung which underscores the importance of mediators such as GM-CSF as both a pro-inflammatory but also an anti-inflammatory mediator. This anti-inflammatory role is supported by studies that showed that GM-CSF has a protective role against pulmonary fibrosis [24] or hyperoxic lung injury [25] in animal models. Both IL-6 and GM-CSF stimulate the marrow to produce and release monocytes while the acute response cytokines, IL-1 and TNF-α, secreted in response to PM10 stimulation by AM [7, 13] induce the production of monocytic chemoattractants such as MCP-1 [20, 21, 26–29]. MIP-1β is a chemotactic factor for human monocytes similar to MIP-1α [22]. Because PM10 did not induce MIP-1β production in human AM [4] or its mRNA in HBEC in the current study, increased MIP-1β expression in the co-cultures most likely relies on an interaction between these two cells. The significance of such an interaction is reinforced by our finding that the production of this chemokine in response to PM10, along with that of GM-CSF and IL-6, is synergistically increased, as noted above, when AM and HBEC are co-cultured. Such a synergistic increase in mediator production could augment the release of both monocytes and polymorphonuclear leukocytes from the bone marrow observed after stimulation by mediators produced by AM incubated alone with EHC-93 ex vivo [6] and thus contribute to a similar response to in vivo exposure to the ambient particles [6, 15].
MCP-1 was the other C-C chemokine that we studied. Along with additional support from results from our laboratory [15], Rosseau and colleagues [30] have shown that the induction of MCP-1 in AM is a major contributor to the recruitment of peripheral blood monocytes into the alveolar compartment. In the present study we showed that production of MCP-1 by AM was just marginally increased by PM10 exposure (p = 0.07). Interestingly, the production and release of MCP-1 by AM/HBEC co-cultures tended (not significant) to be lower than by AM alone (Fig. 3). In AM/HBEC co-cultures, expression of MCP-1 mRNA was significantly increased by PM10 after 2 h but not 24 h exposure suggesting suppression of MCP-1 expression following prolonged exposure of lung cells to particles. This suggests a translational or post-translational control of MCP-1 production and could be an important immunomodulatory pathway by which the local inflammatory reaction in the lung is controlled after PM10 exposure. Together, our findings suggest that both colony stimulating factors and chemokines are released from lung cells following the inhalation of atmospheric particles and that these mediators are critically important in the production and the release of monocytes from the marrow as well as their recruitment into the lung.
The close proximity of AM and epithelial cells in the lung suggests that interaction between these cells is critically important in generating inflammatory mediators in response to noxious stimuli. Previous studies from our laboratory [13] support this concept showing that AM and epithelial cells in co-culture interact to amplify their pro-inflammatory mediator mRNA generation in response to PM10 exposure compared to exposure of mono-cultures of these cells. That soluble factors contribute to this interaction was shown when conditioned media from PM10-stimulated AM induced increases in mRNA expression of many of these mediators in HBEC [13]. On the other hand, that cellular contact between different lung cells (e.g., epithelial, endothelial cells, and fibroblast) is necessary for cell activation and cytokine production [11, 31] has also been demonstrated. Along these lines, we recently showed increased expression of ICAM-1 mRNA after incubation of HBEC with conditioned media from PM10-stimulated AM and this response was blocked by neutralizing antibodies to TNF-α and IL-1β [7]. While TNF-α and IL-1β appear to be major players in the interaction between AM and HBEC in response to PM10, our results suggest that ICAM-1 may play an important role in facilitating the AM-HBEC interaction via these soluble factors.
We postulated that adhesive interactions between CD11/CD18 on AM with ICAM-1 on HBEC contribute to the amplified production of cytokines from AM/HBEC co-cultures observed in the current study. Previous studies have demonstrated that cross-linking CD11b/CD18 on the surface of phagocytes using a combination of either its ligand ICAM-1 or anti-ICAM-1 antibodies primes phagocytes for increased respiratory burst and release of reactive oxygen intermediates [32–34]. In the present study we showed increased epithelial cell surface expression of ICAM-1 induced by PM10 exposure, while CD11b was constitutively expressed on the surface of AM. Cross-linking ICAM-1 on HBEC did not change their PM10-induced mRNA expression. Furthermore, blocking CD11b/CD18 and one of its ligands, ICAM-1 (CD54), did not block or decrease the PM10-induced mRNA expression in AM/HBEC co-cultures. These results are consistent with those of Tao and co-workers [35] who demonstrated that TNF-α and MIP-2 responses to urban air particles in rat AM and RLE (rat alveolar type II epithelial cell line) co-cultures were not blocked with anti-CD18 (β2-integrins)/CD54, arginine-glycine-aspartate peptide (against β1/β3-integrins) and heparin (non-specific anti-inflammatory agent). Paine and colleagues [36] demonstrated that blocking ICAM-1 (anti-CD54 F(ab')2 fragments) decreased rat AM phagocytosis of beads and their planar chemotaxis over the surface of rat alveolar type I epithelial cells. This suggests that ICAM-1 is important for the efficient phagocytosis of particles by AM and promotes mobility of AM on airway epithelial cell surface in the alveolus. Together these studies showed that the β2-integrin/ICAM-1 interaction between AM and lung epithelial cells are important in the chemotaxis of AM in the lung and their phagocytosis of inhaled particles but that this adhesive interaction may not contribute to the mediator production and release by AM and lung epithelial cells. These findings do not exclude the possibility that other adhesive interactions or simultaneous adhesive interactions of more that one adhesion molecule are involved in the particle-induced AM-bronchial epithelial cell mediator response.
Conclusion
Exposure of AM/HBEC co-cultures to ambient particles increased the expression and release of a variety of inflammatory mediators including GM-CSF, M-CSF, IL-6 and MIP-1β that enhance bone marrow production of monocytes and their recruitment into the lung. In addition this type of exposure resulted in synergistic production of GM-CSF and IL-6 in the co cultured cells. The adhesive interaction between ICAM-1 on epithelial cells with the β2-integrin CD11b on AM did not contribute to this synergistic mediator production. We speculate that the interaction between AM and lung epithelial cells amplifies PM10-induced lung inflammation and contributes to the pulmonary morbidity associated with exposure to particulate matter air pollution. This enhanced lung inflammation may also contribute to the systemic inflammatory response as well as the cardiovascular morbidity and mortality induced by air pollution [1, 2, 37].
References
Dockery DW, Pope CA, Xu X, Spengler JD, Ware JH, Fay ME, Ferris BG, Speizer FE: An association between air pollution and mortality in six U.S. cities. N Engl J Med 1993,329(1):753–759.
Pope CA 3rd: Epidemiology of fine particulate air pollution and human health: biologic mechanisms and who's at risk? Environ Health Perspect 2000, 108:713–723.
Schaumann F, Borm PJ, Herbrich A, Knoch J, Pitz M, Schins RP, Luettig B, Hohlfeld JM, Heinrich J, Krug N: Metal-rich Ambient Particles (Particulate Matter2.5) Cause Airway Inflammation in Healthy Subjects. Am J Respir Crit Care Med 2004, 170:898–903.
van Eeden SF, Tan WC, Suwa T, Mukae H, Terashima T, Fujii T, Qui D, Vincent R, Hogg JC: Cytokines involved in the systemic inflammatory response induced by exposure to particulate matter air pollutants (PM(10)). Am J Respir Crit Care Med 2001, 164:826–830.
Monn C, Becker S: Cytotoxicity and induction of proinflammatory cytokines from human monocytes exposed to fine (PM2.5) and coarse particles (PM10–2.5) in outdoor and indoor air. Toxicol Appl Pharmacol 1999, 155:245–252.
Mukae H, Hogg JC, English D, Vincent R, van Eeden SF: Phagocytosis of particulate air pollutants by human alveolar macrophages stimulates the bone marrow. Am J Physiol Lung Cell Mol Physiol 2000, 279:L924–931.
Ishii H, Fujii T, Hogg JC, Hayashi S, Mukae H, Vincent R, van Eeden SF: Contribution of IL-1 beta and TNF-alpha to the initiation of the peripheral lung response to atmospheric particulates (PM10). Am J Physiol Lung Cell Mol Physiol 2004, 287:L176–183.
Carter JD, Ghio AJ, Samet JM, Devlin RB: Cytokine production by human airway epithelial cells after exposure to an air pollution particle is metal-dependent. Toxicol Appl Pharmacol 1997, 146:180–188.
Mills PR, Davies RJ, Devalia JL: Airway epithelial cells, cytokines, and pollutants. Am J Respir Crit Care Med 1999, 160:S38–43.
Fujii T, Hayashi S, Hogg JC, Vincent R, Van Eeden SF: Particulate matter induces cytokine expression in human bronchial epithelial cells. Am J Respir Cell Mol Biol 2001, 25:265–271.
Lang DS, Schocker H, Hockertz S: Effects of crocidolite asbestos on human bronchoepithelial-dependent fibroblast stimulation in coculture: the role of IL-6 and GM-CSF. Toxicology 2001, 159:81–98.
Mukae H, Vincent R, Quinlan K, English D, Hards J, Hogg JC, van Eeden SF: The effect of repeated exposure to particulate air pollution (PM10) on the bone marrow. Am J Respir Crit Care Med 2001, 163:201–209.
Fujii T, Hayashi S, Hogg JC, Mukae H, Suwa T, Goto Y, Vincent R, van Eeden SF: Interaction of alveolar macrophages and airway epithelial cells following exposure to particulate matter produces mediators that stimulate the bone marrow. Am J Respir Cell Mol Biol 2002, 27:34–41.
Goto Y, Hogg JC, Shih CH, Ishii H, Vincent R, van Eeden SF: Exposure to ambient particles accelerates monocyte release from bone marrow in atherosclerotic rabbits. Am J Physiol Lung Cell Mol Physiol 2004, 287:L79–85.
Goto Y, Ishii H, Hogg JC, Shih CH, Yatera K, Vincent R, van Eeden SF: Particulate Matter Air Pollution Stimulates Monocyte Release from the Bone Marrow. Am J Respir Crit Care Med 2004, 170:891–897.
Goto Y, Hogg JC, Suwa T, Quinlan KB, van Eeden SF: A novel method to quantify the turnover and release of monocytes from the bone marrow using the thymidine analog 5'-bromo-2'-deoxyuridine. Am J Physiol Cell Physiol 2003, 285:C253–259.
Vincent R, Bjarnason SG, Adamson IY, Hedgecock C, Kumarathasan P, Guenette J, Potvin M, Goegan P, Bouthillier L: Acute pulmonary toxicity of urban particulate matter and ozone. Am J Pathol 1997, 151:1563–1570.
Koyama Y, Tanaka Y, Saito K, Abe M, Nakatsuka K, Morimoto I, Auron PE, Eto S: Cross-linking of intercellular adhesion molecule 1 (CD54) induces AP-1 activation and IL-1beta transcription. J Immunol 1996, 157:5097–5103.
Wang Q, Doerschuk CM: Neutrophil-induced changes in the biomechanical properties of endothelial cells: roles of ICAM-1 and reactive oxygen species. J Immunol 2000, 164:6487–6494.
Oppenheim J: Human chemokines: an update. Annu Rev Immunol 1998, 15:675–705.
Shanley TP, Warner RL, Ward PA: The role of cytokines and adhesion molecules in the development of inflammatory injury. Mol Med Today 1995, 1:40–45.
Wang JM, Sherry B, Fivash MJ, Kelvin DJ, Oppenheim JJ: Human recombinant macrophage inflammatory protein-1 alpha and -beta and monocyte chemotactic and activating factor utilize common and unique receptors on human monocytes. J Immunol 1993, 150:3022–3029.
Jansen JH, Kluin-Nelemans JC, Van Damme J, Wientjens GJ, Willemze R, Fibbe WE: Interleukin 6 is a permissive factor for monocytic colony formation by human hematopoietic progenitor cells. J Exp Med 1992, 175:1151–1154.
Moore BB, Coffey MJ, Christensen P, Sitterding S, Ngan R, Wilke CA, McDonald R, Phare SM, Peters-Golden M, Paine R 3rd, Toews GB: GM-CSF regulates bleomycin-induced pulmonary fibrosis via a prostaglandin-dependent mechanism. J Immunol 2000, 165:4032–4039.
Paine R 3rd, Wilcoxen SE, Morris SB, Sartori C, Baleeiro CE, Matthay MA, Christensen PJ: Transgenic overexpression of granulocyte macrophage-colony stimulating factor in the lung prevents hyperoxic lung injury. Am J Pathol 2003, 163:2397–2406.
Le J, Vilcek J: Tumor necrosis factor and interleukin 1: cytokines with multiple overlapping biological activities. Lab Invest 1987, 56:234–248.
Dinarello CA: Interleukin-1 and its biologically related cytokines. Adv Immunol 1989, 44:153–205.
Keatings VM, Collins PD, Scott DM, Barnes PJ: Differences in interleukin-8 and tumor necrosis factor-alpha in induced sputum from patients with chronic obstructive pulmonary disease or asthma. Am J Respir Crit Care Med 1996, 153:530–534.
Jiang Y, Beller DI, Frendl G, Graves DT: Monocyte chemoattractant protein-1 regulates adhesion molecule expression and cytokine production in human monocytes. J Immunol 1992, 148:2423–2428.
Rosseau S, Hammerl P, Maus U, Walmrath HD, Schutte H, Grimminger F, Seeger W, Lohmeyer J: Phenotypic characterization of alveolar monocyte recruitment in acute respiratory distress syndrome. Am J Physiol Lung Cell Mol Physiol 2000, 279:L25–35.
Lee YC, Rannels DE: Alveolar macrophages modulate the epithelial cell response to coal dust in vitro. Am J Physiol 1996, 270:L123–132.
Liles WC, Ledbetter JA, Waltersdorph AW, Klebanoff SJ: Cross-linking of CD18 primes human neutrophils for activation of the respiratory burst in response to specific stimuli: implications for adhesion-dependent physiological responses in neutrophils. J Leukoc Biol 1995, 58:690–697.
Rothlein R, Kishimoto TK, Mainolfi E: Cross-linking of ICAM-1 induces co-signaling of an oxidative burst from mononuclear leukocytes. J Immunol 1994, 152:2488–2495.
Shappell SB, Toman C, Anderson DC, Taylor AA, Entman ML, Smith CW: Mac-1 (CD11b/CD18) mediates adherence-dependent hydrogen peroxide production by human and canine neutrophils. J Immunol 1990, 144:2702–2711.
Tao F, Kobzik L: Lung macrophage-epithelial cell interactions amplify particle-mediated cytokine release. Am J Respir Cell Mol Biol 2002, 26:499–505.
Paine R 3rd, Morris SB, Jin H, Baleeiro CE, Wilcoxen SE: ICAM-1 facilitates alveolar macrophage phagocytic activity through effects on migration over the AEC surface. Am J Physiol Lung Cell Mol Physiol 2002, 283:L180–187.
Suwa T, Hogg JC, Quinlan KB, Ohgami A, Vincent R, van Eeden SF: Particulate air pollution induces progression of atherosclerosis. J Am Coll Cardiol 2002, 39:935–942.
Acknowledgements
The authors thank Dr. W. Mark Elliott for technical support and Health Canada for providing the EHC-93. The work was supported by grants from the National Institutes of Health (HL407201), BC Lung Association and the Wolfe & Gita Churg Foundation. SF van Eeden is the recipient of a Career Investigators Award from the American Lung Association and the William Thurlbeck Distinguished Researcher Award.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Authors' contributions
HI carried out all through the experiments and drafted the manuscript. TF, YG, NS and HM participated in the design of the study. SH and JCH participated in its design and helped to draft the manuscript. SFVE conceived of the study, participated in its design and coordination and helped to draft the manuscript. RV provided EHC-93. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Ishii, H., Hayashi, S., Hogg, J.C. et al. Alveolar macrophage-epithelial cell interaction following exposure to atmospheric particles induces the release of mediators involved in monocyte mobilization and recruitment. Respir Res 6, 87 (2005). https://doi.org/10.1186/1465-9921-6-87
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1465-9921-6-87