
Abstract
Background
Autophagy and molecular chaperones both regulate protein homeostasis and maintain important physiological functions. Atg7 (autophagy-related gene 7) and Hsp27 (heat shock protein 27) are involved in the regulation of neurodegeneration and aging. However, the genetic connection between Atg7 and Hsp27 is not known.
Methods
The appearances of the fly eyes from the different genetic interactions with or without polyglutamine toxicity were examined by light microscopy and scanning electronic microscopy. Immunofluorescence was used to check the effect of Atg7 and Hsp27 knockdown on the formation of autophagosomes. The lifespan of altered expression of Hsp27 or Atg7 and that of the combination of the two different gene expression were measured.
Results
We used the Drosophila eye as a model system to examine the epistatic relationship between Hsp27 and Atg7. We found that both genes are involved in normal eye development, and that overexpression of Atg7 could eliminate the need for Hsp27 but Hsp27 could not rescue Atg7 deficient phenotypes. Using a polyglutamine toxicity assay (41Q) to model neurodegeneration, we showed that both Atg7 and Hsp27 can suppress weak, toxic effect by 41Q, and that overexpression of Atg7 improves the worsened mosaic eyes by the knockdown of Hsp27 under 41Q. We also showed that overexpression of Atg7 extends lifespan and the knockdown of Atg7 or Hsp27 by RNAi reduces lifespan. RNAi-knockdown of Atg7 expression can block the extended lifespan phenotype by Hsp27 overexpression, and overexpression of Atg7 can extend lifespan even under Hsp27 knockdown by RNAi.
Conclusions
We propose that Atg7 acts downstream of Hsp27 in the regulation of eye morphology, polyglutamine toxicity, and lifespan in Drosophila.
Similar content being viewed by others


Background
The aging process results from imbalanced homeostasis combined with accumulating macromolecular damage due to different intrinsic and environmental stresses [1–3]. Protein homeostasis is important in maintaining physiological function to protect against cellular degeneration [4]. Autophagy and molecular chaperones are two defensive systems utilized to uphold cellular protein quality and homeostasis [5, 6].
Macroautophagy (herein called autophagy) is a cellular, catabolic process that breaks down and recycles macromolecules and organelles under starvation conditions. Autophagy function is executed by a series of autophagy related genes (Atg) which are evolutionarily conserved from yeast to mammals [7]. Autophagy participates in many physiological functions including aging and neurodegeneration [8, 9], and mounting evidence demonstrates that autophagy participates in the regulation of lifespan in different species [10–12]. In C. elegans, loss-of-function of bec-1/Atg6 or RNA interference-mediated depletion of Atg-7 or Atg-12 inhibits the extended lifespan in daf-2 mutants [13, 14], and the knockdown of bec-1 or Atg7 by RNAi abolishes dietary restriction-mediated longevity in eat-2 mutants [15]. In addition, mutations in Atg1 Atg7 Atg18, and bec-1 reduce lifespan in C. elegans[16]. In Drosophila, Atg7-null mutants are short-lived and hypersensitive to starvation and oxidative stress [17], and the neuronal overexpression of Atg8a regulates lifespan and tolerance to oxidative stress [18]. Atg7 is an E1-like enzyme and is important for the membrane elongation of the autophagosome [7]. Atg7 deficient mice exhibit polyubiquitinated protein accumulation and neurodegeneration [19] and higher levels of polyubiquitinated proteins have been detected in the aging Atg7 mutant fly head [17]. Autophagy also protects against neurodegeneration [20] and the induction of autophagy by the reduction of TOR (target of rapamycin) activity reduces polyglutamine toxicity in both fly and mouse [21]. Suppression of basal autophagy in the central nervous system causes neurodegeneration in Atg7 conditional knockout mice [19, 22].
Molecular chaperones modulate protein re-folding and facilitate the degradation of denatured proteins. Molecular chaperones are also implicated in several physiological functions: autophagy, neurodegeneration, stress tolerance, and aging [23–25]. Heat shock protein 27 (Hsp27) is a member of the ATP-independent, small heat shock protein family. Hsp27 null mutants exhibit decreased lifespan and reduced starvation tolerance [26], while the overexpression of Hsp27 increases lifespan and enhances stress resistance in Drosophila[27, 28]. Overexpression of Hsp27 prevents cellular polyglutamine toxicity and rescues the mosaic eyes induced by mild polyglutamine toxicity [27, 29].
Both Hsp27 and Atg7 are involved in maintaining protein quality and modulating lifespan and neurodegeneration. However, the interaction between Hsp27 and Atg7 is unknown. We report here that Atg7 is downstream of Hsp27 in the regulation of eye morphology, polyglutamine toxicity, and lifespan in Drosophila. The levels of Hsp27 and Atg7 both regulate eye morphology and the polyglutamine toxicity of 41Q. The overexpression of Atg7 rescues both the rough eye phenotype resulting from knockdown of Hsp27 as well as the more severe mosaic eye phenotype induced by the knockdown of Hsp27 under 41Q toxicity. In addition, the expression of Atg7 regulates lifespan in Drosophila and the enhanced lifespan seen with the overexpression of Hsp27 requires the expression of Atg7. Together we provide several lines of genetic evidence linking Hsp27 to Atg7 in the modulation of eye morphology, polyglutamine toxicity, and lifespan regulation.
Methods
Fly strains and maintenance
The RNAi lines were obtained from Vienna Drosophila RNAi Center (VDRC), UAS-hsp27RNAi (#40530), UAS-hsp22RNAi (#43632), UAS-atg1RNAi (#16133), UAS-atg4RNAi (#107317), UAS-atg5RNAi (#104461), UAS-atg7RNAi (#45560), UAS-atg8aRNAi (#43096), UAS-atg8aRNAi (#43097), UAS-atg9RNAi (#10045), UAS-atg12RNAi (#102362), UAS-atg18RNAi (#105366). GMR-Gal4; UAS-41Q and GMR-Gal4/Cyo; UAS-63Q were provided by Dr. Parsa Kazemi-Esfarjani. To generate UAS-Atg7 transgenic flies, the EST clone RE27292 containing the full-length Atg7 was used to amplify the coding sequence by the primers (forward: 5’-GTACTCGAG AAGCAA AACATGAGCACGG-3’ and reverse: 5’-CATAGATCT ATCCTCGTCGCT ATCGGA-3’) and subcloned into the XhoI and BglII sites of the transgenic vector, pINDY6[28]. The resultant construct was verified by DNA sequencing to confirm that no mutations derived from PCR amplification were made, and injected into w1118 eggs for the generation of UAS-Atg7 transgenic flies. All flies were maintained on standard fly food as described in Liu et al.[30] and incubated at 25°C, 65% humidity, in a 12 h/12 h light/dark-cycle fly incubator.
Fly eye image
Two-day-old flies of the different types were anaesthetized by carbon dioxide on a porous platform and the eye images were taken by light microscopy (SMZ1500, Nikon). For the scanning electron micrograph, the fly was fixed on a copper stage and the fly eye image was acquired by scanning electron microscopy (TM-1000, Hitachi). For each fly line, a total of more than 86 eye images from at least three independent crosses were examined.
RT-PCR and real-time PCR
Total RNA was prepared from about 20 flies of each specific allele and homogenized in 1 ml Trizol solution. Equal amounts (1 μg) of each DNase I-treated RNA were reverse-transcribed to cDNA with MMLV reverse transcriptase (Promega). The cDNAs were used as templates for RT-PCR or real-time PCR as described in Liu et al.[30]. The information of the primers is available upon request.
Lifespan and starvation assays
For the lifespan assay, all the fly lines have been outcrossed with w1118 as described previously [31]. The newly eclosed flies of each allele were collected by sex with 30 flies per vial, maintained at 25°C, 65% humidity in a 12 h/12 h light/dark-cycle fly incubator and transferred to a new vial every 3 or 4 days until all were dead. The statistical significance was calculated by log rank test. At least three independent measurements were performed for each experiment.
For the starvation assay, newly eclosed flies of each type were collected by sex with 20 flies per vial and recovered overnight. Next day the flies were transferred to the vials with 1% agar and transferred to new agar vials daily. The numbers of the dead flies were recorded every 4 hours until all were dead. The statistical significance was calculated by student’s t test.
Immunofluorescence
GFP-NLS-marked Atg7 or Hsp27 RNAi knockdown clones in the larval fat body were generated by heat shock-independent FLP/FRT induction as described previously [32, 33]. FLP/FRT method allows to examine the mitotic GFP-NLS-marked RNAi knockdown clones surrounded by the control cells that do not incorporate the RNAi knockdown in the same tissue under the same condition [33]. Fat bodies from early third instar larva cultured in standard fly food with yeast paste (fed condition) or in dishes containing 20% sucrose only (starvation condition) for 4 hr were dissected and fixed with 4% paraformaldehyde and then examined by confocal laser scanning microscope (LSM510; Carl Zeiss Inc.) equipped with a 63x Plan-Apochromat (NA1.4) objective lens.
Results
Autophagy-related gene 7 is downstream of heat shock protein 27 in the regulation of Drosophila eye phenotype
Protein homeostasis plays an important role in lifespan and stress response [1, 2]. Heat shock protein 27 (Hsp27) has been shown to regulate lifespan and response to different stresses [26–28]. Autophagy-related gene 7 (Atg7) is required for normal lifespan and tolerance to starvation and oxidation [17]. However, the genetic interaction between Hsp27 and Atg7 is unknown. We examined the effects of altering Hsp27 and Atg7 expression in the Drosophila eye using the GMR-Gal4 driver followed by the analyses of eye morphology utilizing scanning electron microscopy and light microscopy. Overexpression of Hsp27 or Atg7 results in a normal eye phenotype and regular ommatidia shape as seen in the GMR-Gal4 control flies (Figure 1, A-A”, B-B”, D-D”). Interestingly, knockdown expression of either Hsp27 or Atg7 by expression of interfering RNAs using GMR-Gal4 results in similar rough eye phenotypes with fused and enlarged ommatidia (Figure 1, C-C”, E-E”). Overexpression of Atg7 in the Hsp27 knockdown background fully rescues the rough eye phenotype of the Hsp27 knockdown (Figure 1, F-F”). However, overexpression of Hsp27 in the Atg7 knockdown background fails to rescue the rough eye phenotype of the Atg7 knockdown (Figure 1, G-G”). These results suggest that Atg7 is located downstream of Hsp27 in the regulation of Drosophila eye morphology. To further confirm that Hsp27 and Atg7 function in the same pathway controlling eye phenotype, we examined whether there is any additive effect on fly eye morphology by either the co-overexpression or co-knockdown of Hsp27 and Atg7. The overexpression of both Hsp27 and Atg7 in combination produces a normal eye phenotype, similar to the overexpression of Hsp27 or Atg7 alone (Figure 1, B-B”, D-D”, H-H”). The simultaneous knockdown of Hsp27 and Atg7 does not further deteriorate the rough eye phenotype when compared to the effects of either gene alone (Figure 1, C-C”, E-E”, I-I”), implying that Hsp27 and Atg7 operate in the same pathway. These data provide the first evidence that Atg7 is downstream of Hsp27 in the regulation of Drosophila eye morphology.

Atg7 Is Downstream of Hsp27 in the Regulation of Drosophila Eye Phenotype. (A-A”) The GMR-Gal4/+ control fly has a normal eye phenotype and normal shape of individual ommatidia. (B-B’, D-D”) Overexpression of Hsp27 or Atg7 also results in a normal eye phenotype and regular ommatidia. (C-C”, E-E”) Knockdown of Hsp27 or Atg7 displays similar rough eye phenotype and enlarged and fused ommatidia. (F-F”) Overexpression of Atg7 rescues the rough eye and irregular shape of ommatidia by knockdown of Hsp27. (G-G”) Overexpression of Hsp27 cannot revert the rough eye and abnormal shape of ommatidia resulting from the knockdown of Atg7. (H-H”) Co-overexpression of Hsp27 and Atg7 still leads to normal eye phenotype and ommaditia. (I-I”) Co-knockdown of Hsp27 and Atg7 causes a similar phenotype: rough eyes and irregular shape of ommatidia like that of the individual knockdowns of Hsp27 or Atg7. Optical micrograph (A-I) and scanning electron micrograph (A’-I’: 300X; A”-I”:1500X). Genotypes: GMR-Gal4 in trans to the alleles indicated.
Knockdown of other autophagy-related genes and heat shock protein 22 does not result in a rough eye phenotype in Drosophila
To determine whether the rough eye phenotype is specific to Atg7, or whether it represents a general effect of altering autophagy, the effects of knockdown of additional autophagy-related genes were examined by using GMR-Gal4 and none of these displayed the rough eye phenotype (Figure 2, Figure 1, E-E”). Knockdown of Atg1 shows a normal eye phenotype (Figure 2, A-A”, Figure 1, A-A”), while knockdown of other autophagy-related genes: Atg 4, 5, 8, 9, 12, 18 displayed subtle eye color phenotypes but had no effect on the ommatidia structure (Figure 2, B-G, B’-G’, B”-G”). These data suggests that the rough eye phenotype resulting from Atg7 knockdown is Atg7-specific and not involved in the alteration of other autophagy-related genes. Similarly, to examine whether the rough eye phenotype is specific to Hsp27 knockdown, we tested the effects of knockdown of Hsp22, another known lifespan modulation gene [34], by GMR-Gal4 and did not observe any effects on the eye like that of Hsp27 knockdown (Figure 1, C-C”). Q-PCR analysis confirms that there is reduced expression of Atg and Hsp22 genes in the RNAi knockdown experiments (data not shown). Thus the rough eye phenotype is specific to the knockdown of either Atg7 or Hsp27.

Knockdown of the Other Autophagy-related Genes and Heat Shock Protein 22 Do Not Result in Any Rough Eye Phenotype in Drosophila . RNAi knockdown of different autophagy-related genes by GMR-Gal4 show normal eye morphology and regular ommatidia (like the control in Figure 1, A-A”. (A-A”) Atg1, (B-B”) Atg4, (C-C”) Atg5, (D-D”) Atg8a, (E-E”) Atg9, (F-F”) Atg12, (G-G”) Atg18, and (H-H”) Hsp22. Optical micrograph (A-H) and SEM (A’-H’:300X; A”-H”:1500X). Genotypes: GMR-Gal4 in trans to the alleles indicated.
Knockdown of Atg7 but not Hsp27 blocks starvation-induced autophagosome formation
To verify that the knockdown of Atg7 by UAS-Atg7RNAi from VDRC can affect starvation-induced autophagy, we generated UAS-Atg7RNAi clones in the fat-body by using the FLP/FRT method [32, 33] and examined the distribution of mcherry-Atg8a puncta. The distribution of mcherry-Atg8a is in a uniformly diffuse structure under optimal feeding conditions (Figure 3, B, J) and becomes localized to punctate structure under starvation conditions (Figure 3, F, N). Under starvation conditions, the GFP-NLS clones with the Atg7 knockdown, where the cells are circled by dotted line, display a reduced number of mcherry-Atg8a puncta than the surrounding control clones without Atg7 knockdown which have no GFP-NLS signal (Figure 3, E, F). These results demonstrate that knockdown of Atg7 by UAS-Atg7RNAi is able to block mcherry-Atg8a mediated autophagosome formation under starvation. To examine whether knockdown of Hsp27 can alter autophagosome formation, we also generated UAS-Hsp27RNAi clones in the fat body and inspected the distribution of mcherry-Atg8a puncta. Under starvation, the autophagosome formation indicated by mcherry-Atg8a puncta is not altered by comparing the GFP-NLS marked Hsp27 RNAi knockdown clones, which are circled by dotted line, to the surrounding control clones without GFP-NLS signal and no Hsp27 RNAi knockdown (Figure 3, M, N). The data indicate that Hsp27 knockdown does not block the mcherry-Atg8a mediated autophagosome formation under starvation. The notion is consistent with the previous data since knockdown of Atg8 does not result in the rough eye as the knockdown of Hsp27, suggesting that Hsp27 and Atg8 do not function in the same genetic pathway.

Starvation-induced Autophagosome Formation is Inhibited by RNAi-mediated Depletion of Atg7 but not Hsp27. (A, E, I, M) GFP-NLS labeled fat body cells circled by dotted line indicate the presence of UAS-Atg7RNAior UAS-Hsp27RNAigenerated by the FLP/FRT method. The cells outside of the circled dotted line are used as the control cells without UAS-Atg7RNAior UAS-Hsp27RNAi. (B, F, J, N) The distribution patterns of mcherry-Atg8a are shown under either fully-fed or starvation conditions. (C, G, K, O) The fat body cells are stained with DAPI. (D, L) The picture D is merged from panels A, B, C and L is merged from panels I, J, K under nutrient-rich conditions. (H, P) Picture H is merged from panels E, F, G and P is merged from M, N, O under starvation conditions. The distribution of mcherry-Atg8a puncta is dramatically altered in starved fat body cells (F, N) compared to those under nutrient-rich conditions (B, J). GFP-labeled cells expressing Atg7-RNAi markedly suppress mCherry-Atg8a puncta formation (F), but not in that of Hsp27 knockdown (N).
Atg7 and Hsp27 attenuate the mild polyglutamine toxicity of 41Q but cannot rescue longer polyglutamine tract toxicity by 63Q
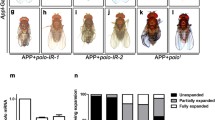
Overexpression of Hsp27 can rescue the mosaic eye phenotype resulting from mild polyglutamine (41Q)-induced toxicity but not the rough eye phenotype resulting from severe polyglutamine (127Q) toxicity [27]. Since Atg7 acts downstream of Hsp27 in the eye, we were interested in determining whether the overexpression of Atg7 would also only rescue mild polygutamine phenotypes. As with Hsp27, the overexpression of Atg7 rescues the mosaic eye phenotype caused by 41Q (Figure 4, A, B, D) but cannot rescue the more severe, rough eye phenotypes resulting from the longer polyglutamine tract of 63Q (Figure 4, G, H, J). The knockdown of either Hsp27 or Atg7 enhances the pigmentation phenotype observed in the eye of flies expressing 41Q. Interestingly only the knockdown of Atg7, but not that of Hsp27, enhances the eye morphology phenotype (rough eye) in combination with 41Q overexpression (Figure 4, C, E). The knockdown of Hsp27 or Atg7 does not exacerbate the rough eye phenotypes of 63Q (Figure 4, I). Interestingly, the overexpression of Atg7 partially rescues the more dramatic mosaic eye phenotype induced by Hsp27 knockdown in the 41Q background (Figure 4, C, F), supporting the idea that Atg7 is downstream of Hsp27 in the alleviation of 41Q toxicity. However, the combination of the overexpression of Atg7 and knockdown of Hsp27 do not change the rough eye phenotype of 63Q (Figure 4, L).

Atg7 is Downstream of Hsp27 in the Attenuation of the Mild Polyglutamine Toxicity by 41Q, but the Overexpression of Both Genes Cannot Rescue the Longer Polyglutamine Tract Toxicity by 63Q. (A) Expression of UAS-41Q by GMR-GAL4 results in mosaic eyes. (B, D) Both the overexpression of Hsp27 and Atg7 rescue the mosaic eye by 41Q. (C, E) Under 41Q background, both the knockdown of Hsp27 and Atg7 generate comparable worsened mosaic eyes, whereas knockdown of Atg7 leads to a rough eye surface. (F) Overexpression of Atg7 as well as knockdown of Hsp27 improves the mosaic eye by 41Q. (G, M) The expressions of UAS-63Q by GMR-Gal4 produce similar rough eye phenotype. (H, J) Overexpression of Hsp27 or Atg7 cannot rescue the rough eye induced by 63Q. (I, K) Knockdown of Hsp27 or Atg7 in conjunction with 63Q does not cause further deterioration of the eyes. (L) Overexpression of Atg7 together with knockdown of Hsp27 does not alter the rough eye phenotype by 63Q. Genotypes: (A-F) GMR-Gal4; UAS-41Q in trans to the alleles indicated. (G-L) GMR-Gal4/Cyo; UAS-63Q in trans to the alleles indicated.
Atg7 regulates lifespan and is required for Hsp27-mediated extended lifespan in Drosophila
Hsp27 levels are likely to regulate Drosophila lifespan since Hsp27 overexpression extends Drosophila lifespan [27, 28] while the knockout Hsp27 mutant is short-lived [26]. The knockdown of Hsp27 by either hs-Gal4, or da-Gal4 exhibits reduced Hsp27 levels and displays a 20% (P < 0.001), and 27% (P < 0.001) decrease in mean lifespan, respectively (Figure 5, A - D; Additional file 1: Table S1). Since Atg7 is downstream of Hsp27 in the regulation of eye morphology and mild polyglutamine toxicity, and Atg7 null mutants display shortened lifespan [17], we tested whether Hsp27-mediated enhanced lifespan requires Atg7. Atg7 overexpression by hs-Gal4 shows a robust increase in Atg7 transcripts relative to control flies and increases the mean lifespan by about 11% (P < 0.01) relative to the control flies (Figure 5, E and G; Additional file 2: Table S2). Conversely, knockdown of Atg7 by hs-Gal4 exhibits reduced levels of Atg7 transcripts and decreases mean lifespan by about 10% (P < 0.01) when compared to the control flies (Figures 5F and H; Additional file 2: Table S2). These results indicate that like Hsp27 Atg7 levels also regulate Drosophila lifespan.

Atg7 Is Downstream of Hsp27 in the Regulation of Drosophila Lifespan. (A, B, E, F) RT-PCR verifies that the transcript levels of Hsp27 and Atg7 are altered upon Gal4 induction. (C, D) RNAi knockdown of Hsp27 by hs-Gal4 and da-Gal4 both reduce Drosophila lifespan. (G) Overexpression of Atg7 by hs-Gal4 increases Drosophila lifespan. (H) Knockdown of Atg7 by hs-Gal4 decreases Drosophila lifespan. (I) Neuronal overexpression of Atg7 by appl-Gal4 enhances Drosophila lifespan. (J) Overexpression of Atg7 along with knockdown of Hsp27 by appl-Gal4 displays extended lifespan. On the other hand, simultaneous knockdown of Atg7 and overexpression of Hsp27 exhibits reduced lifespan.
It has been shown that neuronal overexpression of Atg8a by appl-Gal4 extends Drosophila lifespan and increases resistance to starvation [18]. To test whether neuronal overexpression of Atg7 enhances lifespan and starvation resistance, Atg7 was overexpressed in neurons using appl-Gal4, resulting in increases of 12% (P < 0.001) in mean lifespan and 18% (P < 0.01) in starvation resistance (Figure 5, I; Additional file 2: Table S2 and Additional file 3:Table S4). In addition, the simultaneous overexpression of Atg7 and knockdown of Hsp27 results in flies that exhibit a 21% (P < 0.001) extension in mean lifespan (Figure 5, J). Conversely, the flies possessing both knockdown of Atg7 and overexpressing Hsp27 display a reduction of 27% (P < 0.001) in mean lifespan relative to the control flies (Figure 5, J; Additional file 4 : Table S3). To further demonstrate that Atg7 functions downstream of Hsp27, we carried out the locomotion assay to measure the climbing activity of the flies with the different combination of overexpression and knockdown of Atg7 and Hsp27 along with the control flies under paraquat-induced oxidative stress. Similar to the lifespan result, the flies with simultaneous overexpression of Atg7 and knockdown of Hsp27 displayed significantly better climbing activity (42%, P ≤ 0.001) than that of the control flies (22%), and the flies with simultaneous knockdown of Atg7 and overexpression of Hsp27 exhibited a significantly lowered locomotion activity (15%, P ≤ 0.01) than that of the control flies (Additional file 5 : Figure S1). The climbing activity data in accordance with the lifespan data supports our hypothesis that Atg7 acts downstream of Hsp27. Taken together, these results indicate that as seen with Drosophila eye morphology and polyglutamine toxicity, Atg7 also acts downstream of Hsp27 in regulating lifespan.
Discussion
Hsp27 and Atg7 are both implicated in the processes of aging and neurodegeneration. In this report, we provide several lines of evidence to show that Atg7 is downstream of Hsp27 in the regulation of eye morphology, polyglutamine toxicity, and lifespan. Autophagy-related genes are conserved among different species [7, 35]. Each of the identified Atgs has a role in autophagy, but their roles in other processes remains largely unclear.
In the examination of eye phenotype, we observed that the knockdown of either Hsp27 or Atg7 exhibited similar rough eye phenotypes. These effects appear to be specific to these particular molecules since the knockdown of other Atgs (Atg1 Atg4 Atg5 Atg8a Atg9 Atg12 and Atg18) or Hsp22 does not produce a similar, rough eye phenotype. The ability of Atg7 to rescue the phenotype induced by Hsp27 knockdown also suggests that a unique interaction exists between Hsp27 and Atg7. A recent study indicates that knockdown of Atg7 by GMR-Gal4 on X chromosome causes retinal degeneration [36]. In addition, the rhabdomeres were shown degenerated in the aged atg7d77 mutant flies [37]. Both support our finding that RNAi knockdown of Atg7 results in rough eye in Drosophila.
Autophagy serves to protect against neurodegenerative diseases [20] and aberrations in autophagy have been implicated in neurodegeneration [38]. In both fly and mouse models, induction of autophagy by inhibiting mTOR ameliorates polyglutamine toxicity [21]. And in humans, a polymorphism study of more than 900 European Huntington’s disease patients revealed that one variant of Atg7 (Atg7V471A) is statistically correlated with early onset of Huntington’s disease [39]. These findings suggest that a specific function of Atg7 is to attenuate polyglutamine toxicity and support our findings that Atg7 rescues polyglutamine toxicity by 41Q in Drosophila. Hsp27 has also been shown to reduce cellular polyglutamine toxicity [29] and the overexpression of Hsp27 in Drosophila rescues the pigmentation defects induced by 41Q [27]. Several lines of evidence suggest that heat shock proteins may rely upon autophagy to reduce polyglutamine toxicity. For example, the anti-polyglutamine-aggregation activity of HspB7, one of the human small heat shock proteins, was substantially diminished in Atg5-deficient cells [40]. In addition, it is possible that the small heat shock protein HspB8-Bag3 complex enhance Htt43Q degradation via autophagy since the treatment of the Htt43Q transfected HEK-293T and COS1 cells with an autophagy inhibitor significantly reduced HspB8-Bag3-mediated Htt43Q degradation [41]. Furthermore, it was recently suggested that the small heat shock protein HspB7 assists in the loading of misfolded proteins or aggregates in autophagosomes [42]. Together, these findings indicate that autophagy is downstream of small heat shock proteins and support our results that Atg7 is downstream of Hsp27.
The inhibition of autophagy results in decreased lifespan. Atg7 activity is essential for the longevity resulting from either reduced insulin signaling or caloric restriction in which depletion of Atg7 was found to block the longevity phenotypes of both daf-2 and eat-2 mutants [13, 15]. Our data showed that RNAi knockdown of Atg7 by hs-Gal4, starting from embryonic to adulthood stage, results in a shortened lifespan similar to that of the Drosophila Atg7 null mutant [17]. Loss-of-function mutations in Atg7 as well as Atg1 Atg18, and Beclin-1 shorten lifespan in C. elegans[16]. Several autophagy mutants including Atg7 were identified chronologically short-lived in a yeast genetic screen [43]. However, it should be noted that not all autophagy genes are linked to aging and Atg7 is one of the conserved Atg genes that is involved in the regulation of aging in most species [9]. Conversely, the induction of autophagy increases lifespan. The induction of autophagy by caloric restriction or reducing target of rapamycin (TOR) activity enhances lifespan [9] and the neuronal overexpression of Atg8a increases Drosophila lifespan [18]. We have found that the overexpression of Atg7 extends lifespan in Drosophila and that the neuronal overexpression of Atg7 is sufficient to reverse Hsp27-knockdown-mediated, shortened lifespan. Knockdown of Atg7 blocks Hsp27-mediated extended lifespan, again supporting the model that Atg7 acts downstream of Hsp27 in the regulation of lifespan. It has been reported that in adult flies, RNAi knockdown of Atg7 by Geneswitch-Actin-Gal4 did not show reduced lifespan [44]. This discrepancy may be due to the different Gal4 drivers used and that the knockdown of Atg7 occurring only during adulthood is insufficient to cause shortened lifespan since autophagy activity is known to be tightly regulated during development.
Yet we cannot exclude that chaperone-mediated autophagy (CMA) is involved in the connection between Hsp27 and Atg7. CMA is a specific cargo delivery process to the lumen of the lysosome, mediated by Hsc70, Hsp90, and the lysosome-associated membrane protein type 2A (LAMP-2A) [45, 46]. However, a recent study in Drosophila shows that the co-chaperone Starvin assists in the coordination of Hsc70 and HspB8 through chaperone-assisted selective autophagy, which is distinct from CMA, to depose damaged filamin for muscle maintenance [47]. It is possible that Hsp27 may function through chaperone-assisted selective autophagy linking to Atg7.
Conclusion
In summary, our finding sheds new insight in the linkage of Hsp27 to Atg7 in the regulation of eye morphology, polyglutamine toxicity, and lifespan. The information provides a new aspect in the understanding how Hsp27 may connect to Atg7 to modulate certain physiological functions.
Abbreviations
- Atg:
-
autophagy-related gene
- Hsp:
-
heat shock protein.
References
Haigis MC, Yankner BA: The aging stress response. Mol Cell. 2010, 40 (2): 333-344. 10.1016/j.molcel.2010.10.002.
Koga H, Kaushik S, Cuervo AM: Protein homeostasis and aging: The importance of exquisite quality control. Ageing Res Rev. 2011, 10 (2): 205-215. 10.1016/j.arr.2010.02.001.
Taylor RC, Dillin A: Aging as an event of proteostasis collapse. Cold Spring Harb Perspect Biol. 2011, 3 (5): a004440-10.1101/cshperspect.a004440.
Douglas PM, Dillin A: Protein homeostasis and aging in neurodegeneration. J Cell Biol. 2010, 190 (5): 719-729. 10.1083/jcb.201005144. 2010/09/08 ed
Tyedmers J, Mogk A, Bukau B: Cellular strategies for controlling protein aggregation. Nat Rev Mol Cell Biol. 2010, 11 (11): 777-788. 10.1038/nrm2993.
Wong E, Cuervo AM: Integration of clearance mechanisms: the proteasome and autophagy. Cold Spring Harb Perspect Biol. 2011, 2 (12): a006734-
He C, Klionsky DJ: Regulation mechanisms and signaling pathways of autophagy. Annu Rev Genet. 2009, 43: 67-93. 10.1146/annurev-genet-102808-114910.
Kroemer G, Marino G, Levine B: Autophagy and the integrated stress response. Mol Cell. 2010, 40 (2): 280-293. 10.1016/j.molcel.2010.09.023.
Rubinsztein DC, Marino G, Kroemer G: Autophagy and aging. Cell. 2011, 146 (5): 682-695. 10.1016/j.cell.2011.07.030.
Chang YY, Neufeld TP: Autophagy takes flight in Drosophila. FEBS Lett. 2010, 584 (7): 1342-1349. 10.1016/j.febslet.2010.01.006.
Cuervo AM, Bergamini E, Brunk UT, Droge W, Ffrench M, Terman A: Autophagy and aging: the importance of maintaining "clean" cells. Autophagy. 2005, 1 (3): 131-140. 10.4161/auto.1.3.2017.
Madeo F, Tavernarakis N, Kroemer G: Can autophagy promote longevity?. Nat Cell Biol. 2010, 12 (9): 842-846. 10.1038/ncb0910-842.
Hars ES, Qi H, Ryazanov AG, Jin S, Cai L, Hu C, Liux LF: Autophagy regulates ageing in C. elegans. Autophagy. 2007, 3 (2): 93-95.
Melendez A, Talloczy Z, Seaman M, Eskelinen EL, Hall DH, Levine B: Autophagy genes are essential for dauer development and life-span extension in C. elegans. Science. 2003, 301 (5638): 1387-1391. 10.1126/science.1087782.
Jia K, Levine B: Autophagy is required for dietary restriction-mediated life span extension in C. elegans. Autophagy. 2007, 3 (6): 597-599.
Toth ML, Sigmond T, Borsos E, Barna J, Erdelyi P, Takacs-Vellai K, Orosz L, Kovacs AL, Csikos G, Sass M, Vellai T: Longevity pathways converge on autophagy genes to regulate life span in Caenorhabditis elegans. Autophagy. 2008, 4 (3): 330-338.
Juhasz G, Erdi B, Sass M, Neufeld TP: Atg7-dependent autophagy promotes neuronal health, stress tolerance, and longevity but is dispensable for metamorphosis in Drosophila. Genes Dev. 2007, 21 (23): 3061-3066. 10.1101/gad.1600707.
Simonsen A, Cumming RC, Brech A, Isakson P, Schubert DR, Finley KD: Promoting basal levels of autophagy in the nervous system enhances longevity and oxidant resistance in adult Drosophila. Autophagy. 2008, 4 (2): 176-184.
Komatsu M, Waguri S, Chiba T, Murata S, Iwata J, Tanida I, Ueno T, Koike M, Uchiyama Y, Kominami E, Tanaka K: Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature. 2006, 441 (7095): 880-884. 10.1038/nature04723.
Garcia-Arencibia M, Hochfeld WE, Toh PP, Rubinsztein DC: Autophagy, a guardian against neurodegeneration. Semin Cell Dev Biol. 2010, 21 (7): 691-698. 10.1016/j.semcdb.2010.02.008.
Ravikumar B, Vacher C, Berger Z, Davies JE, Luo S, Oroz LG, Scaravilli F, Easton DF, Duden R, O'Kane CJ, Rubinsztein DC: Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of Huntington disease. Nat Genet. 2004, 36 (6): 585-595. 10.1038/ng1362.
Hara T, Nakamura K, Matsui M, Yamamoto A, Nakahara Y, Suzuki-Migishima R, Yokoyama M, Mishima K, Saito I, Okano H, Mizushima N: Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature. 2006, 441 (7095): 885-889. 10.1038/nature04724.
Lanneau D, Brunet M, Frisan E, Solary E, Fontenay M, Garrido C: Heat shock proteins: essential proteins for apoptosis regulation. J Cell Mol Med. 2008, 12 (3): 743-761. 10.1111/j.1582-4934.2008.00273.x.
Tower J: Heat shock proteins and Drosophila aging. Exp Gerontol. 2011, 46 (5): 355-362. 10.1016/j.exger.2010.09.002.
Voisine C, Pedersen JS, Morimoto RI: Chaperone networks: tipping the balance in protein folding diseases. Neurobiol Dis. 2010, 40 (1): 12-20. 10.1016/j.nbd.2010.05.007.
Hao X, Zhang S, Timakov B, Zhang P: The Hsp27 gene is not required for Drosophila development but its activity is associated with starvation resistance. Cell Stress Chaperones. 2007, 12 (4): 364-372. 10.1379/CSC-308.1.
Liao PC, Lin HY, Yuh CH, Yu LK, Wang HD: The effect of neuronal expression of heat shock proteins 26 and 27 on lifespan, neurodegeneration, and apoptosis in Drosophila. Biochem Biophys Res Commun. 2008, 376 (4): 637-641. 10.1016/j.bbrc.2008.08.161.
Wang HD, Kazemi-Esfarjani P, Benzer S: Multiple-stress analysis for isolation of Drosophila longevity genes. Proc Natl Acad Sci U S A. 2004, 101 (34): 12610-12615. 10.1073/pnas.0404648101.
Wyttenbach A, Sauvageot O, Carmichael J, Diaz-Latoud C, Arrigo AP, Rubinsztein DC: Heat shock protein 27 prevents cellular polyglutamine toxicity and suppresses the increase of reactive oxygen species caused by huntingtin. Hum Mol Genet. 2002, 11 (9): 1137-1151. 10.1093/hmg/11.9.1137.
Liu YL, Lu WC, Brummel TJ, Yuh CH, Lin PT, Kao TY, Li FY, Liao PC, Benzer S, Wang HD: Reduced expression of alpha-1,2-mannosidase I extends lifespan in Drosophila melanogaster and Caenorhabditis elegans. Aging Cell. 2009, 8 (4): 370-379. 10.1111/j.1474-9726.2009.00471.x.
Wang CT, Chen YC, Wang YY, Huang MH, Yen TL, Li H, Liang CJ, Sang TK, Ciou SC, Yuh CH, Wang CY, Brummel TJ, Wang HD: Reduced neuronal expression of ribose-5-phosphate isomerase enhances tolerance to oxidative stress, extends lifespan, and attenuates polyglutamine toxicity in Drosophila. Aging Cell. 2012, 11 (1): 93-103. 10.1111/j.1474-9726.2011.00762.x.
Tang HW, Wang YB, Wang SL, Wu MH, Lin SY, Chen GC: Atg1-mediated myosin II activation regulates autophagosome formation during starvation-induced autophagy. EMBO J. 2011, 30 (4): 636-651. 10.1038/emboj.2010.338.
Theodosiou NA, Xu T: Use of FLP/FRT system to study Drosophila development. Methods. 1998, 14 (4): 355-365. 10.1006/meth.1998.0591.
Morrow G, Samson M, Michaud S, Tanguay RM: Overexpression of the small mitochondrial Hsp22 extends Drosophila life span and increases resistance to oxidative stress. FASEB J. 2004, 18 (3): 598-599.
Kourtis N, Tavernarakis N: Autophagy and cell death in model organisms. Cell Death Differ. 2009, 16 (1): 21-30. 10.1038/cdd.2008.120.
Midorikawa R, Yamamoto-Hino M, Awano W, Hinohara Y, Suzuki E, Ueda R, Goto S: Autophagy-dependent rhodopsin degradation prevents retinal degeneration in Drosophila. J Neurosci. 2010, 30 (32): 10703-10719. 10.1523/JNEUROSCI.2061-10.2010.
Wang T, Lao U, Edgar BA: TOR-mediated autophagy regulates cell death in Drosophila neurodegenerative disease. J Cell Biol. 2009, 186 (5): 703-711. 10.1083/jcb.200904090.
Marino G, Madeo F, Kroemer G: Autophagy for tissue homeostasis and neuroprotection. Curr Opin Cell Biol. 2011, 23 (2): 198-206. 10.1016/j.ceb.2010.10.001.
Metzger S, Saukko M, Van Che H, Tong L, Puder Y, Riess O, Nguyen HP: Age at onset in Huntington's disease is modified by the autophagy pathway: implication of the V471A polymorphism in Atg7. Hum Genet. 2010, 128 (4): 453-459. 10.1007/s00439-010-0873-9.
Vos MJ, Zijlstra MP, Kanon B, van Waarde-Verhagen MA, Brunt ER, Oosterveld-Hut HM, Carra S, Sibon OC, Kampinga HH: HSPB7 is the most potent polyQ aggregation suppressor within the HSPB family of molecular chaperones. Hum Mol Genet. 2010, 19 (23): 4677-4693. 10.1093/hmg/ddq398.
Carra S, Seguin SJ, Lambert H, Landry J: HspB8 chaperone activity toward poly(Q)-containing proteins depends on its association with Bag3, a stimulator of macroautophagy. J Biol Chem. 2008, 283 (3): 1437-1444.
Vos MJ, Zijlstra MP, Carra S, Sibon OC, Kampinga HH: Small heat shock proteins, protein degradation and protein aggregation diseases. Autophagy. 2011, 7 (1): 101-103. 10.4161/auto.7.1.13935.
Matecic M, Smith DL, Pan X, Maqani N, Bekiranov S, Boeke JD, Smith JS: A microarray-based genetic screen for yeast chronological aging factors. PLoS Genet. 2010, 6 (4): e1000921-10.1371/journal.pgen.1000921.
Ren C, Finkel SE, Tower J: Conditional inhibition of autophagy genes in adult Drosophila impairs immunity without compromising longevity. Exp Gerontol. 2009, 44 (3): 228-235. 10.1016/j.exger.2008.10.002.
Arias E, Cuervo AM: Chaperone-mediated autophagy in protein quality control. Curr Opin Cell Biol. 2011, 23 (2): 184-189. 10.1016/j.ceb.2010.10.009.
Dice JF: Chaperone-mediated autophagy. Autophagy. 2007, 3 (4): 295-299.
Arndt V, Dick N, Tawo R, Dreiseidler M, Wenzel D, Hesse M, Furst DO, Saftig P, Saint R, Fleischmann BK, Hoch M, Hohfeld J: Chaperone-assisted selective autophagy is essential for muscle maintenance. Curr Biol. 2010, 20 (2): 143-148. 10.1016/j.cub.2009.11.022.
Acknowledgements
We thank Drs. Theodore Brummel, Micheline Laurent, William Ja, Pankaj Kapahi, and Chiou-Hwa Yuh for the critical reading and suggestions for the manuscript. We thank the fly lines from VDRC and Bloomington stock center, GMR-Gal4; UAS-41Q and GMR-Gal4/Cyo; UAS-63Q from Dr. Parsa Kazemi-Esfarjani. We thank the funding support from National Tsing Hua University/Chang Gung Memorial Hospital collaboration grant (101N2754E1), NTHU/McKay Hospital collaboration grant (99N2903E1), and National Science Council (100-2311-B-007-006-) to Dr. Horng-Dar Wang, National Science Council grant (100-2311-B-004-001-MY3) to Dr. Pei-Yu Wang, and Academia Sinica (AS-99-TP-B09) to Dr. Guang-Chao Chen. We are indebted for the funding support from Brain Research Center (101N2060E1) by Dr. Ann-Shyn Chiang at NTHU, the fly import assistance from Fly Core Taiwan by Dr. Chau-Ti Ting, and the transgenic fly support by Dr. Y. Henry Sun at Academia Sinica, Taipei, Taiwan.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing Interests
The authors declare that they have no competing interests.
Authors’ contributions
S-F Chen, M-L Kang, Y-C Chen, H-W Tang, C-W Huang, W-H Li, and C-P Lin carried out the experiments and analyzed the data; P-Y Wang, G-C Chen and H-D Wang designed the experiments, analyzed the data, and together with C-Y Wang discussed the data and wrote the manuscript. All authors read and approved the final manuscript.
Shih-Fen Chen, Ming-Lun Kang, Yi-Chun Chen, Hong-Wen Tang contributed equally to this work.
Electronic supplementary material
12929_2012_404_MOESM2_ESM.docx
Additional file 2: Table S2. A summary of lifespan by the overexpression or knockdown of Atg7 in Drosophila.(DOCX 21 KB)
12929_2012_404_MOESM3_ESM.docx
Additional file 3: Table S4. A summary of starvation stress response by overexpression of Atg7 in Drosophila.(DOCX 20 KB)
12929_2012_404_MOESM4_ESM.docx
Additional file 4: Table S3. A summary of lifespan resulting from simultaneous overexpression and knockdown of different combinations of Atg7 and Hsp27 in Drosophila.(DOCX 19 KB)
12929_2012_404_MOESM5_ESM.docx
Additional file 5: Figure S1. The flies with simultaneous overexpression of Atg7 and knockdown of Hsp27 display better climbing activity than those with overexpression of Hsp27 and knockdown of Atg7 under paraquat-induced oxidative stress. The climbing index for each strain: appl-Gal4/+(the control fly): 21.8 ± 0.02% (n = 195); UAS-hsp27/+; appl-Gal4/UAS-atg7RNAi: 14.7 ± 0.02% (n = 123); UAS-atg7/+; appl-Gal4/UAS-hsp27RNAi: 42.4 ± 0.01% (n = 175). (n is the total fly number from the four independent assays.). (**p < 0.01, ***p < 0.001). (DOCX 102 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Chen, SF., Kang, ML., Chen, YC. et al. Autophagy-related gene 7 is downstream of heat shock protein 27 in the regulation of eye morphology, polyglutamine toxicity, and lifespan in Drosophila. J Biomed Sci 19, 52 (2012). https://doi.org/10.1186/1423-0127-19-52
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1423-0127-19-52