
Abstract
Background
ER stress is a strong indicator of whether or not a cell is undergoing physiological stress. C. elegans is a practical system of characterizing the effect of ER stress at the in vivo or organismal level.
Methods
This study characterized taurine’s anti-ER stress potential employing western blotting on ER stress markers and assays of motility, lifespan comparison, and fecundity measurement.
Results
When treated with tunicamycin, C. elegans showed the typical ER stress symptoms. It showed a higher expression of hsp-70 and skn-1 than the non-treated control. Survivorship significantly decreased under tunicamycin treatment, and the offspring number also decreased. During the synchronized culture under ER stress conditions, the C. elegans showed early signs of aging especially between L3 and L4 within their life span, along with lowered motility. The worms, however, showed a positive response to the taurine treatment under ER stress conditions.
Conclusions
When C. elegans were treated with taurine before or after the tunicamycin treatment, they showed a less severe level of ER stress, including an enhanced survivorship, increased motility, and augmented fecundity. Taken together, these results strongly indicate that taurine works positively to cope with ER stress from the organismal perspective.
Similar content being viewed by others


Background
Taurine is known to help cells recover from damage, and to prevent physiological stress by adjusting osmolarity. Under cellular stress conditions, taurine maintains the cellular homeostasis by achieving an osmotic balance within the cells; in detail, by controlling the functional gating of the ion channels involved in the intercellular ion trafficking [1]. No concrete mechanism, however, is available to explain the ability of taurine to counteract the harmful effects of physiological stress, such as endoplasmic reticulum (ER) stress.
ER stress symptoms serve as legitimate indicators of whether or not cells are undergoing physiological stress [2–5]. ER stress has been well characterized in terms of cellular response to various causative agents. The in vivo effect of ER stress is poorly understood, however, although a mouse model has been developed to monitor ER stress in vivo. There have been significant inconsistencies between in vitro and in vivo experiments on the ER stress response. This strongly implies that the organismal response represents the mixture of different cellular outcomes, and that the in vivo ER stress response may employ different elements along the pathway.
Despite the simple body scheme of C. elegans, it has been proven as sharing numerous vital biological pathways with mammals. It has become a valuable animal system in gaining an integrated understanding of organismal reactions to various forms of environmental and physiological stress [5–7]. Due to its short generation period, it enables researchers to study the effect of certain agents or conditions on aging and fecundity [8, 9]. Under certain circumstances, C. elegans provides a practical system of studying both taurine’s effect on various ER stress responses at the organismal level and C. elegans’ anti-ER stress capability.
To characterize the potential anti-ERS mechanism of taurine, C. elegans was treated with tunicamycin, an ER stress inducer. Under the induced ER stress conditions, the effect of taurine was studied by monitoring the difference in the expression between ER stress marker protein (hsp-70) and the factor mediating adaptive responses to cellular stress (skn-1). The skn-1 is known to improve the ER stress conditions that negatively affected the lifespan, mobility, and fecundity of the C. elegans [10]. These three categories of stress physiological marker were also used to characterize whether taurine serves as an anti-ER stress mediator.
Methods
C. elegans were normally grown at 25oC in the nematode growth medium (NGM) [11–13]. To induce ER stress conditions, worms were treated with tunicamycin at 10 μg/ml. To assay the effect of taurine, the worms were incubated with various extracellular taurine concentrations after they were treated for 12 h under ERS conditions. The ER stress conditions were verified by monitoring the ER stress marker expression. Equal homogenized samples were electrophoresed on a 10% SDS PAGE. Protein expression was quantified and standardized to the expression of actin protein. The values of the relative expression were obtained against the control treatments. The expression of skn-1 and hsp-70 was detected via western blotting using antibodies that were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The western blotting was carried out according to the standard procedure, and the antibodies were diluted at 1:1,000.
The lifespan of the C. elegans was determined according to the method that Hyun et al. used [14]. After the worms were sacrificed by bleaching, 10 eggs were placed on NGM supplemented with OP50 at 25°C up to the young adult stage. Approximately 50 worms were placed on plates that contained 0-10 μg/ml of tunicamycin for 3 h. Then they were transferred to media that contained 10 and 100 μg/ml of taurine. They were monitored until they no longer responded to gentle stimulation with a platinum wire. For all the lifespan experiments, the assays were repeated twice.
To analyze the effect of taurine on the mobility of taurine, the distance of the movement of the worms that were treated in the presence of taurine and the distance of movement of the worms that were treated without taurine were compared. Initially, the worms were treated with 10 μg/ml of tunicamycin and transferred to taurine-containing media. Following the worms’ relocation to the taurine media, their total moving distance was determined from the track they had made.
The numbers of eggs that were laid were compared after the taurine treatment under the ER stress conditions. The worms were maintained on NGM plates covered with a lawn of OP-50 until they showed fresh moult. The adults were selected for uniformity and transferred to a fresh plate that contained 5 μg/ml of tunicamycin. Following their incubation for 6 h, they were transferred to taurine-containing media. Their fecundity was measured according to the combined number of their fertilized eggs and larvae for 3 days.
Results and discussion
This study evaluated the anti-ER stress effect of taurine by determining if it is capable of reducing the stress caused by tunicamycin. The expression of hsp70, which was high after the tunicamycin treatment, considerably decreased under the taurine treatment in a dose-dependent manner. The skn-1 expression decreased when the worms were treated with taurine. In terms of the organismal markers, taurine showed an ER stress relieving effect by restoring the level of survivorship, fecundity, and motility of the worms.
ER stress marker expression was reduced when worms were treated with taurine
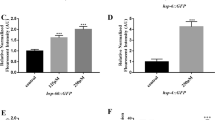
Based on the ER stress marker expression, taurine appears to have exerted an inhibitory effect on the progression of ER stress. When the cells were treated with taurine after their ER stress exposure, the level of hsp-70 significantly increased relative to the other ER stress markers. The significant upregulation of the ER stress markers suggests that taurine retards ER stress. The level of hsp70 expression showed typical dose-dependence, along with the amount of the tunicamycin that was added (Fig. 1). When the worms were incubated with various taurine concentrations, however, taurine downgraded the hsp-70 protein expression depending on the taurine concentration. The data imply that taurine lessens the intensity of ER stress caused by tunicamycin.

Heat shock protein expression under taurine treatment. C. elegans were treated with taurine for 12 h after they had been cultured with tunicamycin. The hsp-70 expression was compared among four taurine concentrations. Protein expression was quantified and standardized to actin protein expression.
The Increase of skn-1 is evident under taurine treatment
The expression of skn-1 increased after the tunicamycin treatment, albeit differently from hsp70; but its expression significantly increased under the taurine treatment in a dose-dependent manner (Fig. 2). With regard to skn-1, however, the timing of the taurine treatment affected the expression of skn-1. When taurine was applied after tunicamycin, taurine had a more instantaneous effect on the level of the skn-1 expression (right panel) than when they were pre-treated with taurine (left panel). The results indicate that taurine is capable of lowering the level of ER stress by an increase in the level of skn-1, an anti-oxidative stress protein. This observation strongly implies that an existing ER stress response accelerates the expression of skn-1 by factors from the stress or by mechanisms to be further elucidated.

Skn-1 expression Worms were cultured with taurine either before or after treatment with tunicamycin. The expression of skn-1 protein expression was detected via western blotting.
This oxidative stress response is very important as a cellular defense function and appears to be widely conserved during evolution. Oxidative stress induces skn-1 to accumulate in intestinal nuclei. Skn-1 is distantly related to the nrf (nuclear receptor factor) proteins that induce phase II detoxification gene transcription in mammals. The increase in skn-1 upon taurine treatment has a significant meaning, since ER stress could be diminished, as evidenced in Fig. 1, by the induction of detoxification gene expression, or by augmented expression of skn-1.
Tunicamycin causes an increment in the reactive oxygen species (ROS), along with ER stress. Increased skn-1 expression may not help worms avoid the toxicity of tunicamycin-causing oxidative stress [15–18]. Should taurine’s efficacy as anti-ER stress agent be validated in subsequent experiments, this result would strongly imply that taurine may help lessen ER stress via the de novo anti-oxidative stress pathway.
The expression of hsp-70 decreased under taurine treatment, but that of skn-1 increased. The increase in the skn-1 expression with taurine treatment is confusing, since its higher expression may refer to an increasing level of ER stress under taurine treatment [19–21]. Unlike hsp-70, skn-1 may play a more functional role in lessening the severity of ER stress than other ER stress markers.
Taurine prolongs the lifespan of ER stressed worms
The worms’ lifespan was negatively affected by their treatment with tunicamycin. When the treatment began one day after the initiation of the culture, the worms responded sensitively to tunicamycin, and many of them died (Fig. 3). After they were treated with tunicamycin alone, half of them died within 20 days after the start of the treatment. The taurine treatment, however, greatly improved the worms’ survival rate. No difference was seen between the tunicamycin and the tunicamycin-free treatment groups. Depending on the concentration of the taurine treatment, however, the worms recovered their lifespan up to the level where no tunicamycin was applied.

Life span assay Worms were grown on taurine-containing media (1 or 10 mg/ml) following the treatment with tunicamycin (10 μg/ml). Survivorship was recorded daily following taurine treatment of the worms which were subjected to ER stress. The percentage of live worms greatly increased in a dose dependent fashion.
Taurine helps restore the worm’s fecundity
Taurine’s effect on fecundity was measured following its application to tunicamycin-treated worms. After the tunicamycin treatment, the worms laid much fewer eggs. When the worms were cultured on taurine media, however, their egg-laying ability normalized (Fig. 4). This increased level of fecundity could be attributed to taurine’s reduction of the level of ER stress [22–24].

Fecundity assay Under taurine treatment, tunicamycin-induced stress appeared to decrease in terms of fecundity. Each value represents the mean of three experiments. Taurine appeared more effective when it is applied before the addition of tunicamycin than the “after” treatment (10 μg/ml).
Taurine helped C. elegans recover from tunicamycin-reduced motility
When the C. elegans were treated with tunicamycin, their mobility more significantly decreased than with the non-tunicamycin control. They substantially recovered their movement ability, however, within one hour after the transfer. The values of their total moving distance for different concentrations of media-applied taurine were compared. Fig. 5 shows the different effects of the taurine and the taurine-free treatment with different doses. This result strongly implies that taurine helped the worms recover from the negative effect of ER stress on their muscular activity [25–28].

Motility comparison The extent of worms’ movement was visually compared among various taurine treatments. The upper panel shows the different degrees of mobility between taurine (left) and taurine-free treatment (right). Although tunicamycin retarded worm’s movement, its negative effect was compensated by taurine in the media (lower panel or table).
Conclusions
Upon treatment with tunicamycin, C. elegans showed typical ER stress symptoms, such as elevated expression of heat shock proteins (Hsp-70). Skn-1 expression increased and induced an anti-oxidative stress pathway. The tunicamycin treatment also caused many physiological stress symptoms: decreased survivorship, retarded movement, low fecundity, and early aging. When the worms were treated with taurine, however, they showed positive responses against ER stress conditions. They showed less ER stress, longer survivorship, and improved mobility and fecundity. These results strongly indicate that taurine fights ER stress from the in vivo physiological perspective.
Abbreviations
- ER:
-
endoplasmic reticulum
- nrf:
-
nuclear receptor factor
- hsp:
-
heat shock protein
- NGM:
-
nematode growth media
References
Malhotra JD, Kaufman RJ: The endoplasmic reticulum and the unfolded protein response. Semin Cell Dev Biol. 2007, 18: 716-731. 10.1016/j.semcdb.2007.09.003.
Shen X, Ellis RE, Sakaki K, Kaufman RJ: Genetic interactions due to constitutive and inducible gene regulation mediated by the unfolded protein response in C. elegans. PLoS Genet. 2005, 1: e37-10.1371/journal.pgen.0010037.
Marciniak SJ, Yun CY, Oyadomari S, Novoa I, Zhang Y, Jungreis R, Nagata K, Harding HP, Ron D: CHOP induces death by promoting protein synthesis and oxidation in the stressed endoplasmic reticulum. Genes Dev. 2004, 18 (24): 3066-3077. 10.1101/gad.1250704.
Lisse TS, Thiele F, Fuchs H, Hans W, Przemeck GKH, Abe K, Rathkolb B, Quintanilla-Martinez L, Hoelzlwimmer G, Helfrich M, Wolf E, Ralston SH, de Angelis MH: ER Stress-mediated apoptosis in a new mouse model of osteogenesis imperfecta. PLoS Genet. 2008, 4 (2): e7-10.1371/journal.pgen.0040007.
Ayyadevara S, Dandapat A, Singh SP, Siegel ER, Shmookler Reis RJ, Zimniak L, Zimniak P: Life span and stress resistance of Caenorhabditis elegans are differentially affected by glutathione transferases metabolizing 4-hydroxynon-2-enal. Mech Ageing Dev. 2007, 128 (2): 196-205. 10.1016/j.mad.2006.11.025.
Boyd WA, McBride S, Freedman JH: Effects of genetic mutations and chemical exposures on Caenorhabditis elegans feeding: Evaluation of a novel, high-throughput screening assay. PLoS ONE. 2007, 2 (12): e1259-10.1371/journal.pone.0001259.
Girard LR, Fiedler TJ, Harris TW, Carvalho F, Antoshechkin I, Han M, Sternberg PW, Stein LD, Chalfie M: WormBook: the online review of Caenorhabditis elegans biology. Nucleic Acids Res. 2007, 35: D472-D475. 10.1093/nar/gkl894.
Link EM, Hardiman G, Sluder AE, Johnson CD, Liu LX: Therapeutic target discovery using Caenorhabditis elegans. Pharmacogenomics. 2000, 1: 203-217. 10.1517/14622416.1.2.203.
Silverman GA, Luke CJ, Bhatia SR, Long OS, Vetica AC, Perlmutter DH, Pak SC: Modeling molecular and cellular aspects of human disease using the nematode Caenorhabditis elegans. Pediatr Res. 2009, 65: 10-18. 10.1203/PDR.0b013e31819009b0.
Oliveira RP, Abate JP, Dilks K, Landis J, Ashraf J, Murphy CT, Blackwell TK: Condition-adapted stress and longevity gene regulation by Caenorhabditis elegans SKN-1/Nrf. Aging Cell. 2009, 8: 524-41. 10.1111/j.1474-9726.2009.00501.x.
Stiernagle T: Maintenance of C. elegans. WormBook. The C. elegans Research Community. WormBook. 2006, doi/10.1895/wormbook.1.101.1, [http://www.wormbook.org]
Strange K, Yan X, Lorin-Nebel C, Xing J: Physiological roles of STIM1 and Orai1 homologs and CRAC channels in the genetic model organism Caenorhabditis elegans. Cell Calcium. 2007, 42: 193-203. 10.1016/j.ceca.2007.02.007.
Szewczyk NJ, Kozak E, Conley CA: Chemically defined medium and Caenorhabditis elegans. BMC Biotechnol. 2003, 3: 19-10.1186/1472-6750-3-19.
Hyun MJ, Lee JH, Lee KJ, May A, Bohr VA, Ahn BC: Longevity and resistance to stress correlate with DNA repair capacity in Caenorhabditis elegans. Nucleic Acids Res. 2008, 36: 1380-1389. 10.1093/nar/gkm1161.
Estes S, Ajie BC, Lynch M, Phillips PC: Spontaneous mutational correlations for life-history, morphological and behavioral characters in Caenorhabditis elegans. Genetics. 2005, 170: 645-653. 10.1534/genetics.104.040022.
van Anken E, Braakman I: Endoplasmic reticulum stress and the making of a professional secretory cell. Crit Rev Biochem Mol Biol. 2005, 40: 269-283. 10.1080/10409230500315352.
Ron D, Walter P: Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol. 2007, 8: 519-529. 10.1038/nrm2199.
Sasagawa Y, Yamanaka K, Ogura T: ER E3 ubiquitin ligase HRD-1 and its specific partner chaperone BiP play important roles in ERAD and developmental growth in Caenorhabditis elegans. Genes Cells. 2007, 12: 1063-1073. 10.1111/j.1365-2443.2007.01108.x.
Scheuner D, Kaufman RJ: The unfolded protein response: A pathway that links insulin demand with beta-cell failure and diabetes. Endocr Rev. 2008, 29: 317-333. 10.1210/er.2007-0039.
Balklava Z, Pant S, Fares H, Grant BD: Genome-wide analysis identifies a general requirement for polarity proteins in endocytic traffic. Nat Cell Biol. 2007, 9: 1066-1073. 10.1038/ncb1627.
Nass R, Hall DH, Miller DM, Blakely RD: Neurotoxin-induced degeneration of dopamine neurons in Caenorgabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 2002, 99: 3264-3269. 10.1073/pnas.042497999.
Yoshiuchi K, Kaneto H, Matsuoka TA, Kohno K, Wawaki T, Nakatani Y, Yamasaki Y, Hori M, Matsuhisa M: Direct monitoring of in vivo ER stress during the development of insulin resistance with ER stress-activated indicator transgenic mice. Biochem Biophys Res Comm. 2008, 366: 545-550. 10.1016/j.bbrc.2007.11.182.
Walter MF, Biessmann MR, Benitez C, Török T, Mason JM, Biessmann H: Effects of telomere length in Drosophila melanogaster on life span, fecundity and fertility. Chromosoma. 2007, 116: 41-51. 10.1007/s00412-006-0081-5.
Ratcliff WC, Hawthorne P, Travisano M, Denison RF: When Stress Predicts a Shrinking Gene Pool, Trading Early Reproduction for Longevity Can Increase Fitness, Even with Lower Fecundity. PLoS ONE. 2009, 4: e6055-10.1371/journal.pone.0006055.
Huang GJ, Kim Y, Dehoff M, Mizuno Y, Kamm KE, Worley PF, Muallem S, Zeng W: Ca2+ signaling in microdomains: Homer1 mediates the interaction between RyR2 and Cav1.2 to regulate excitation-contraction coupling. J Biol Chem. 2007, 282: 14283-14290. 10.1074/jbc.M611529200.
Nass R, Blakely RD: The Caenorhabditis elegans dopaminergic system: Opportunities for insights into dopamine transport and neurodegeneration. Annu Rev Pharmacol Toxicol. 2003, 43: 521-544. 10.1146/annurev.pharmtox.43.100901.135934.
Huang G, Yao J, Zeng W, Mizuno Y, Kamm KE, Stull JT, Harding HP, Ron D, Muallem S: ER stress disrupts Ca2+-signaling complexes and Ca2+ regulation in secretory and muscle cells from PERK-knockout mice. J Cell Sci. 2006, 119: 153-161. 10.1242/jcs.02731.
Tcherpakov M, Broday L, Delaunay A, Kadoya T, Khurana A, Erdjument-Bromage H, Tempst P, Qiu XB, DeMartino GN, Ronai Z: JAMP Optimizes ERAD to Protect Cells from Unfolded Proteins. Mol Biol Cell. 2008, 19: 5019-5028. 10.1091/mbc.E08-08-0839.
Acknowledgements
This article has been published as part of Journal of Biomedical Science Volume 17 Supplement 1, 2010: Proceedings of the 17th International Meeting of Taurine. The full contents of the supplement are available online at http://www.jbiomedsci.com/supplements/17/S1.
This work was supported by the University of Seoul 2009 Research Fund to DHL. We acknowledge the UOS Molecular Biology Lab students for their invaluable contributions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
HMK carried out the maintenance of C. elegans and participated in the taurine treatment and data collection. CHD participated in the design of the study and performed the statistical analysis. DHL participated in obtaining worm strains and coordination of the experiments and helped to draft the manuscript. All authors read and approved the final manuscript.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Kim, H.M., Do, CH. & Lee, D.H. Taurine reduces ER stress in C. elegans. J Biomed Sci 17 (Suppl 1), S26 (2010). https://doi.org/10.1186/1423-0127-17-S1-S26
Published:
DOI: https://doi.org/10.1186/1423-0127-17-S1-S26