
Abstract
Classical scrapie is a naturally transmitted prion disease of sheep and goats. Contaminated environments may contribute to the spread of disease and evidence from animal models has implicated urine, blood, saliva, placenta and faeces as possible sources of the infection. Here we sought to determine whether sheep naturally infected with classical scrapie shed prions in their faeces. We used serial protein misfolding cyclic amplification (sPMCA) along with two extraction methods to examine faeces from sheep during both the clinical and preclinical phases of the disease and showed amplification of PrPSc in 7 of 15 and 14 of 14 sheep respectively. However PrPSc was not amplified from the faeces of 25 sheep not exposed to scrapie. These data represent the first demonstration of prion shedding in faeces from a naturally infected host and thus a likely source of prion contamination in the environment.
Similar content being viewed by others
Introduction
Prion diseases, or transmissible spongiform encephalopathies (TSEs), are fatal, neurological disorders that affect a range of mammalian species. They include scrapie in sheep, chronic wasting disease (CWD) in deer, bovine spongiform encephalopathy (BSE) in cattle and Creutzfeldt-Jakob disease (CJD) in humans. The "protein-only" hypothesis is the widely accepted paradigm for disease propagation and is based on the conversion of an apparently benign host prion protein (PrPC) into a pathological isoform, PrPSc, which constitutes the infectious agent [1]. To date PrPSc is the only validated biochemical marker for prion diseases. These disorders are characterised by prolonged incubation periods before the onset of clinical signs, and the accumulation of PrPSc particularly within the CNS. The sites of PrPSc accumulation vary depending on the combination of host and prion strain under investigation, for instance classical ovine scrapie, cervid CWD and human vCJD display widespread distribution of PrPSc within the CNS and lymphoreticular tissues (LRS) [2–4] whereas for cattle BSE, atypical scrapie and human CJD the accumulation of PrPSc within LRS is much more restricted [5, 6].
To date, studies investigating the shedding of the prion agent have focussed on scrapie and CWD. There is accumulating evidence for the excretion of prions within milk [7, 8], urine [9, 10], saliva [10–12] and faeces [13]. Such reports utilise methodologies that are exquisitely sensitive in their detection of prion infectivity or PrPSc, namely bioassay or amplification of PrPSc by serial protein misfolding cyclic amplification (sPMCA). PMCA was pioneered by Soto et al. [14] in a rodent model of scrapie and this is an in vitro technique that amplifies trace amounts of PrPSc within a test sample during iterative rounds of sonication and incubation at 37°C. Importantly, this technique has been adapted for the high level amplification of PrPSc from natural sources of prion infection including ovine scrapie [15], bovine BSE [16], human CJD [17] and cervid CWD [18].
There is evidence from experimentally infected animals that prions are excreted in faeces. Safar et al. [13] used a rodent scrapie model to demonstrate the presence of prions in faeces when measured by conformation-dependent immunoassay (CDI) and transgenic mouse bioassay. Prions were present in faeces from hamsters throughout disease incubation. An analogous study in hamsters employed the sPMCA amplification of prions in faecal extracts and demonstrated the presence of prions during clinical disease [19]. In addition, experimentally infected mule deer excreted prion infectivity in faeces during preclinical and clinical stages of CWD [20].
Here, we apply sPMCA to investigate the excretion of prions within the faeces of naturally infected sheep, both in the preclinical and clinical stages of disease progression.
Materials and methods
Samples
Faecal samples were removed directly from the gut at post mortem or from the rectum of live animals while avoiding contamination with blood. Visible contamination with blood was not observed in any of the samples. This method of collection also minimised any contamination from the environment. Samples were stored at -20°C until assayed. Faeces were collected from sheep homozygous for VRQ (codons 136, 154 and 171 respectively of the PRNP gene) at 9 to 10 months of age and at clinical end-point (mean survival time of 22 months of age). As negative controls, faeces were collected from VRQ/VRQ unexposed sheep from a VLA New Zealand derived flock. No consistent differences in the consistency of the faeces were observed between exposed and negative control sheep.
Faeces suspension preparation
Method 1 [Silicon dioxide (SiO2)]: After thawing, faecal pellets were transferred to 50 mL disposable tubes for homogenisation and diluted to three times the original mass using phosphate buffered saline (PBS, 15 mM KH2PO4, 81 mM Na2HPO4, 137 mM NaCl, and 3 mM KCl) containing 2× miniprotease inhibitors (without EDTA, Roche, Welwyn Garden City, UK). Each faecal suspension was vortexed for 5 to 10 min until homogeneous and 0.5 mL aliquots of this 33% faecal homogenate were transferred to a 15 mL falcon tube and stored at -80°C until required.
Method 2 [Sodium Phosphotungstic acid (NaPTa)]: After thawing the faecal samples were transferred to disposable tubes for homogenisation and diluted 1:9 in water containing protease inhibitors (Roche). The samples were then homogenised using the Prionics PrioGENISER™ (Prionics, Zurich, Switzerland) twice for 40 s at maximum speed. Prions were then extracted from faecal homogenates using NaPTa.
Extraction of prions
Method 1 [SiO2]: Faecal homogenates (0.5 mL of a 33% w/v faecal homogenate) were diluted 1:1 in 2× phosphate buffer (pH 7) containing 1% deoxycholate (DOC) and 1% NP40, then further diluted to 5 mL with 1× PBS containing 0.5% DOC and 0.5% NP40. The faecal homogenates were then incubated for 1 h at room temperature (RT). Following centrifugation at 1500 g for 3 min the supernatant was removed and the pellet discarded. 80 μL of a 20% (w/v) SiO2 suspension were added to the 5 mL faecal supernatant and the sample rotated gently for 2 h at RT. The samples were centrifuged at 700 g for 4 min and the supernatant discarded. The SiO2 pellets were washed by gentle pipetting using 1 mL 0.1% SDS.
Method 2 [NaPTa]: Faecal homogenates (0.5 mL of a 10% w/v homogenate in deionised water containing protease inhibitors (mini-pill complete, Roche)) were transferred to homogenisation tubes and sodium dodecyl sulphate (SDS) added to a final concentration of 1%. The samples were then ribolysed (BioRad, Hemel Hempstead, UK) three times for 45 s each and cooled between each cycle. The samples were incubated at RT for 1 h rotating gently. Homogenates were then centrifuged for 20 min at 4°C at 15 000 g and the supernatants transferred to a clean tube. The pellets were discarded. The supernatant was diluted 1:1 with PBS containing 4% N-Lauroylsarcosine sodium salt (Sigma, Poole, UK) and treated with Benzonaze (1 unit/μL, Sigma) for 30 min at 50°C. Following treatment, NaPTa was added to a final concentration of 0.57% and the sample incubated at 37°C rotating gently for 30 min. Samples were centrifuged at 15 000 g for 15 min at 10°C and the supernatant discarded. Pellets were frozen at -80°C. Upon analyses, pellets were resuspended in 50 μL of PMCA buffer (PBS, additional 150 mM NaCl, 4 mM EDTA, pH8.0, 1.0% (v/v) Triton X-100 and miniprotease inhibitor, Roche).
Amplification of prions
Method 1 [SiO2]: Faecal extracts derived from SiO2 precipitation were centrifuged at 16 000 g for 3 min. 10 μL of each faecal extract supernatant (from the equivalent of 8.3 mg wet weight of faeces) were mixed 1:9 with normal ovine brain homogenate. The samples were then subjected to 40 s cycles of sonication every 30 min incubating at 37°C for 24 h (1 round), after which the amplified samples were diluted 1:2 with PMCA substrate in a final volume of 100 μL and the sample subjected to further rounds of PMCA. Ten rounds of amplification were carried out. The amplification product was stored at -20°C. Samples from sheep incubating scrapie as well as from sheep not exposed to the scrapie agent were analyzed concurrently within the same run on the same sonicator. Each SiO2 extract was amplified within two separate experiments. After 10 rounds of amplification in the absence of PolyA no de novo synthesis was observed. Amplification was not extended beyond 10 rounds.
Method 2 [NaPTA]: Faecal extracts derived from NaPTa precipitation (8 μL; from the equivalent of 8 mg wet weight of faeces) were mixed 1:9 with normal ovine brain homogenate (obtained at post mortem from a VRQ/VRQ sheep from the New Zealand-derived VLA scrapie-free flock; a 10% (w/v) homogenate in PMCA buffer) supplemented with synthetic polyA RNA to a final concentration 100 μg/mL. The samples were then subjected to 40 s cycles of sonication every 30 min incubating at 37°C for 24 h (1 round). The sonicated product was then replenished 1:4 with fresh normal brain homogenate and sonication repeated for a further 24 h. This was repeated until four rounds were completed. Using this PMCA protocol rounds were restricted to 4 because de novo synthesis was observed at 6 rounds or more under PrPSc free conditions (data not shown). No de novo synthesis was observed in the presence of PolyA at round 4. The amplification products were stored after all PMCA rounds at -20°C until tested for PrPSc content.
Analysis of PrPSc
PMCA products were tested using the IDEXX HerdChek antigen test for scrapie as directed by the manufacturers. Positives were scored using the manufacturer's cut-off values. Alternatively, PMCA samples were digested with 50 μg/mL proteinase K, 0.045% (w/v) SDS for 1 h at 37°C before analysis by Western blotting using 12% (w/v) NuPAGE pre-cast Bis-Tris gels as previously described [21].
Statistical analysis
When comparing the percentage of positive sPMCA reactions for different cohorts of samples, data were set up as 2 × 2 contingency tables and Fisher's exact test (one-tailed) was applied to derive p-values.
Results
Faeces were sampled directly from the transverse colon of clinically-affected sheep at post mortem or from the rectum of live sheep with preclinical scrapie. Prions were extracted from faeces employing a mild detergent extraction (0.5% w/v DOC and NP40) followed by SiO2 precipitation. Prions extracted using SiO2 were amplified over 10 rounds of sPMCA before attempted detection of PrPSc by Western blot. This method was previously shown to amplify prions from the saliva of sheep incubating scrapie [11]. Here, when applied to the faeces from scrapie infected animals, PrPSc was amplified from 6 of 29 animals: four animals with clinical disease and two animals which were clinically normal at approximately the mid-point of disease incubation (10 positive sPMCA reactions from 58 analyses; Figure 1, Table 1). No prions were amplified from faeces collected from sheep which had not been exposed to the scrapie agent (10 animals, 60 analyses, Figure 1, Table 1). These data demonstrate that prions were secreted in faeces from sheep incubating natural scrapie (p = 0.001).

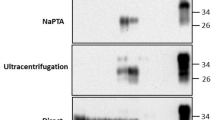
Prion excretion in faeces of healthy and clinical scrapie infected sheep. Prions extracted from 8.3 mg of faeces were used as seed for Method 1 [10 rounds of sPMCA]. After 4 to 10 rounds of sPMCA (as indicated), products were digested with proteinase K and 10 μL applied to Western blots. PrP was detected with monoclonal antibodies SHA31 and P4; molecular weight markers are shown (M1, 20 and 30 kDa; M2, 19 and 28 kDa). Sheep were either in the preclinical (lanes 1 and 2 for animals 1236/09 and 1305/09 respectively) or clinical stages of disease progression (lanes 3-6 for animals 0320/09, 0324/09, 0470/09 and 0572/09 respectively) or were unexposed to the scrapie agent (lanes 7-16 for animals R185, R223, R233, R234, R297, R303, R328, R382, R413 and R421 respectively). Two analyses were carried out for each sample and representative analyses are shown. Cerebellum (2.5 μL of a 10% w/v homogenate) from a terminally affected sheep was analysed on each blot as a PrPSc control (C).
In order to estimate the levels of scrapie prions present within faeces, the limit of detection of the methodology was determined for the earliest sPMCA round that amplified PrPSc from both preclinical and clinical faecal samples. Using SiO2 extraction coupled with sPMCA amplification of prions in the absence of polyA RNA, seven rounds of sPMCA amplification yielded detectable PrPSc for 4 of the samples, 3 taken from clinical animals (0320/09, 0324/09, 0572/09) and one taken from an animal in the preclinical stage of disease (1305/09). A dilution series of a brain homogenate from an animal with terminal scrapie was spiked into a faecal homogenate and then extracted and amplified. Extraction followed by amplification for 7 rounds of sPMCA detected PrPSc down to 0.64 ng of brain material spiked into 167 mg wet weight of faeces (data not shown).
To investigate further the shedding of prions in faeces, a second method of extraction and amplification was developed. This method used potentially more stringent extraction conditions and enhanced sPMCA amplification. Briefly, prions were eluted from faeces with 1% SDS followed by NaPTa precipitation of prions and sPMCA amplification employing polyA;-the presence of polyA having been shown to increase the sensitivity of the sPMCA amplification [15, 22]. The presence of PrPSc was demonstrated in the amplification product using the IDEXX HerdChek antigen assay for scrapie (Table 1). PrPSc was detected in the faeces of all 14 sheep which were in the preclinical stages of scrapie and sampled at 9 to 10 months of age (19 positive sPMCA reactions from 22 analyses; Table 1). Of the sheep at clinical end-stage, PrPSc was amplified from the faeces of 6 out of 15 sheep (8 out of 27 analyses were positive). Analyses of animals not exposed to the scrapie agent again demonstrated that no prions could be detected (25 animals, 45 analyses; Table 1 and data not shown). These data indicate that scrapie prions are excreted in faeces in both preclinical (p < 0.001) and clinical (p < 0.001) stages of the disease. PrPSc was detected at the earliest following round 4 using this method except in 1 sample (324/09) where PrPSc was detected following round 3.
These data report for the first time the excretion of prions in the faeces of naturally infected animals and further support the role of the faecal-oral route as a means of horizontal transmission in the field.
Discussion
The reported study demonstrates the presence of prions in the faeces of sheep naturally infected with scrapie. Additionally, PrPSc was present in the faeces of sheep both in the terminal stages of disease and when in the early preclinical phase, suggesting that prions are likely to be shed into the environment throughout pathogenesis. These data concur with a study of mule deer experimentally infected with CWD [20] where infectivity was observed in faecal samples collected from the deer from nine months after inoculation and maintained during the clinical phase several months later. Together these data are fully consistent with faeces being a source of environmental prion contamination that contributes to transmission for both scrapie and CWD.
Within the present study, two distinct methods were applied to detect faecal PrPSc. Prions were detected less frequently when employing mild detergent extraction, SiO2 precipitation and a standard sPMCA amplification, compared to when using SDS extraction, NaPTa precipitation and sPMCA incorporating polyA. This may indicate that the latter method has an improved sensitivity. The method based on mild extraction and non-enhanced sPMCA detected prions more frequently in clinical samples and in contrast the method based on SDS extraction and polyA-enhanced amplification detected prions more frequently in preclinical samples. This suggests that preclinical animals may be more likely to secrete prions compared to animals with clinical disease, but that the levels of prion present within faeces is higher in the clinical animals. With the mild extraction conditions amplification of prions was observed at earlier rounds in samples from clinical compared to preclinical animals, supporting the proposition that prions are at higher levels in faeces during clinical disease.
Several sources of prions in faeces could be postulated including environmental ingestion and swallowing of infected saliva [11]. However, the most likely source is shedding from the gut-associated lymphoid tissue. Early and rapid accumulation of PrPSc in the Peyer's patches is consistent with the detection of prions in faeces during the preclinical phase. In sheep with a VRQ/VRQ PRNP genotype this accumulation commences at around 2-3 months of age. Ruminants have specialised Peyer's patches that are continuous throughout the length of the ileum amounting to approximately 100 000 follicles and all of these could potentially be infected and shedding prions into the lumen. However the number of follicles diminishes at around 12-15 months of age thus reducing the potential for shedding at this site in older sheep such as those with clinical disease in the present study. Additionally, the recto-anal mucosa associated lymphoid tissue may also play a role in contributing to the levels of PrPSc found in the faeces; in the present study this route would not contribute to prion in the samples from clinical sheep due to direct sampling from the gut. These factors would be expected to result in lower prion levels and/or a lower frequency of prion excretion within the clinical faecal samples. However, this is not supported by the presented data indicating that levels of prion excretion is not solely influenced by excretion from RAMALT or the ileal Peyer's patches.
Not all prion diseases result in infection of the gut-associated lymphoid tissue. The low levels of prion infection in the gut of cattle with natural BSE [23] could explain the apparent, fortuitous failure of lateral spread of BSE within herds.
Here, with the estimated levels of prion shed within the faeces combined with an estimated ovine faecal output of 67 kg/month [24] it could be speculated that between 9 months and 22 months of age sheep could excrete prions equivalent to the potentially infectious load within approximately 3.3 mg of clinically infected brain. Tamguney et al. [20] used bioassay to estimate the levels of infectivity in faeces from experimentally-infected CWD deer; over a 10 month period these levels were estimated to approach that present within a clinically infected whole brain. These levels are much higher than those found in the present study with scrapie infected sheep; however, this could reflect differences between the prion strains and host species under investigation or be due to differences in the prion detection methodologies employed.
Previous studies have indicated that risk factors for horizontal transmission in the field involve contact during the postnatal period [25–27]. However, exposure to pasture that has been grazed by infected sheep is sufficient to transmit disease even in the absence of lambing [25, 28]. The findings of the present study would suggest that prions in faeces will contribute to this contamination and support the role of the faecal-oral route as a means of horizontal transmission. Strategies for the control of scrapie will need to take this into account particularly for goats where successful elimination is still reliant on depopulation, disinfection and restocking.
In North America, the presence of prions in secreta and excreta of deer and the resultant contamination of the environment is likely to have contributed to the spread of CWD and this is likely to progress unabated given the estimation of the amount of prions contained within cervine faeces [20]. Therefore environmental contamination of pasture remains a serious impediment to future eradication of prion disease in both Europe and in North America.
References
Prusiner SB: Prions. Proc Natl Acad Sci USA. 1998, 95 (23): 13363-13383. 10.1073/pnas.95.23.13363.
Langeveld JP, Jacobs JG, Erkens JH, Bossers A, van Zijderveld FG, van Keulen LJ: Rapid and discriminatory diagnosis of scrapie and BSE in retro-pharyngeal lymph nodes of sheep. BMC Vet Res. 2006, 2: 19-10.1186/1746-6148-2-19.
Wadsworth JDF, Joiner S, Hill AF, Campbell TA, Desbruslais M, Luthert PJ, Collinge J: Tissue distribution of protease resistant prion protein in variant Creutzfeldt-Jakob disease using a highly sensitive immunoblotting assay. Lancet. 2001, 358 (9277): 171-180. 10.1016/S0140-6736(01)05403-4.
Fox KA, Jewell JE, Williams ES, Miller MW: Patterns of PrPCWD accumulation during the course of chronic wasting disease infection in orally inoculated mule deer (Odocoileus hemionus). J Gen Virol. 2006, 87 (Pt 11): 3451-3461.
Wells GAH, Hawkins SAC, Green RB, Austin AR, Dexter I, Spencer YI, Chaplin MJ, Stack MJ, Dawson M: Preliminary observations on the pathogenesis of experimental BSE: an update. Vet Rec. 1998, 142: 103-106. 10.1136/vr.142.5.103.
Hill AF, Butterworth RJ, Joiner S, Jackson G, Rossor MN, Thomas DJ, Frosh A, Tolley N, Bell JE, Spencer M, King A, Al-Sarraj S, Ironside JW, Lantos PL, Collinge J: Investigation of variant Creutzfeldt-Jakob disease and other human prion diseases with tonsil biopsy samples. Lancet. 1999, 353 (9148): 183-189. 10.1016/S0140-6736(98)12075-5.
Konold T, Moore SJ, Bellworthy SJ, Simmons HA: Evidence of scrapie transmission via milk. BMC Vet Res. 2008, 4 (1): 14-10.1186/1746-6148-4-14.
Maddison BC, Baker CA, Rees HC, Terry LA, Thorne L, Bellworthy SJ, Whitelam GC, Gough KC: Prions are secreted in milk from clinically normal scrapie-exposed sheep. J Virol. 2009, 83 (16): 8293-8296. 10.1128/JVI.00051-09.
Gonzalez-Romero D, Barria MA, Leon P, Morales R, Soto C: Detection of infectious prions in urine. FEBS Lett. 2008, 582 (21-22): 3161-3166. 10.1016/j.febslet.2008.08.003.
Haley NJ, Seelig DM, Zabel MD, Telling GC, Hoover EA: Detection of CWD prions in urine and saliva of deer by transgenic mouse bioassay. PLoS One. 2009, 4 (3): e4848-10.1371/journal.pone.0004848.
Maddison BC, Rees HC, Baker CA, Taema M, Bellworthy SJ, Thorne L, Terry LA, Gough KC: Prions are secreted into the oral cavity in sheep with preclinical scrapie. J Infect Dis. 2010, 1672-1676. 11
Mathiason CK, Powers JG, Dahmes SJ, Osborn DA, Miller KV, Warren RJ, Mason GL, Hays SA, Hayes-Klug J, Seelig DM, Wild MA, Wolfe LL, Spraker TR, Miller MW, Sigurdson CJ, Telling GC, Hoover EA: Infectious prions in the saliva and blood of deer with chronic wasting disease. Science. 2006, 314 (5796): 133-136. 10.1126/science.1132661.
Safar JG, Lessard P, Tamguney G, Freyman Y, Deering C, Letessier F, Dearmond SJ, Prusiner SB: Transmission and detection of prions in feces. J Infect Dis. 2008, 198 (1): 81-89. 10.1086/588193.
Saborio GP, Permanne B, Soto C: Sensitive detection of pathological prion protein by cyclic amplification of protein misfolding. Nature. 2001, 411 (6839): 810-813. 10.1038/35081095.
Thorne L, Terry LA: In vitro amplification of PrPSc derived from the brain and blood of sheep infected with scrapie. J Gen Virol. 2008, 89 (Pt 12): 3177-3184.
Gough KC, Baker CA, Taema M, Maddison BC: In vitro amplification of prions from milk in the detection of subclinical infections. Prion. 2009, 3 (4): 236-239. 10.4161/pri.3.4.10425.
Jones M, Peden AH, Prowse CV, Groner A, Manson JC, Turner ML, Ironside JW, MacGregor IR, Head MW: In vitro amplification and detection of variant Creutzfeldt-Jakob disease PrPSc. J Pathol. 2007, 213 (1): 21-26. 10.1002/path.2204.
Kurt TD, Perrott MR, Wilusz CJ, Wilusz J, Supattapone S, Telling GC, Zabel MD, Hoover EA: Efficient in vitro amplification of chronic wasting disease PrPRES. J Virol. 2007, 81 (17): 9605-9608. 10.1128/JVI.00635-07.
Kruger D, Thomzig A, Lenz G, Kampf K, McBride P, Beekes M: Faecal shedding, alimentary clearance and intestinal spread of prions in hamsters fed with scrapie. Vet Res. 2009, 40 (1): 4-10.1051/vetres:2008042.
Tamguney G, Miller MW, Wolfe LL, Sirochman TM, Glidden DV, Palmer C, Lemus A, DeArmond SJ, Prusiner SB: Asymptomatic deer excrete infectious prions in faeces. Nature. 2009, 461 (7263): 529-532. 10.1038/nature08289.
Rees HC, Maddison BC, Owen JP, Whitelam GC, Gough KC: Concentration of disease-associated prion protein with silicon dioxide. Mol Biotechnol. 2009, 41 (3): 254-262. 10.1007/s12033-008-9129-5.
Deleault NR, Lucassen RW, Supattapone S: RNA molecules stimulate prion protein conversion. Nature. 2003, 425 (6959): 717-720. 10.1038/nature01979.
Terry LA, Marsh S, Ryder SJ, Hawkins SA, Wells GA, Spencer YI: Detection of disease-specific PrP in the distal ileum of cattle exposed orally to the agent of bovine spongiform encephalopathy. Vet Rec. 2003, 152 (13): 387-392. 10.1136/vr.152.13.387.
Smith KA, Frost JP: Nitrogen excretion by farm livestock with respect to land spreading requirements and controlling nitrogen losses to ground and surface waters. Part 1: cattle and sheep. Biosresource Technol. 2000, 71: 173-181. 10.1016/S0960-8524(99)00061-9.
Dexter G, Tongue SC, Heasman L, Bellworthy SJ, Davis A, Moore SJ, Simmons MM, Sayers AR, Simmons HA, Matthews D: The evaluation of exposure risks for natural transmission of scrapie within an infected flock. BMC Vet Res. 2009, 5: 38-10.1186/1746-6148-5-38.
Touzeau S, Chase-Topping ME, Matthews L, Lajous D, Eychenne F, Hunter N, Foster JD, Simm G, Elsen JM, Woolhouse ME: Modelling the spread of scrapie in a sheep flock: evidence for increased transmission during lambing seasons. Arch Virol. 2006, 151 (4): 735-751. 10.1007/s00705-005-0666-y.
Woolhouse ME, Stringer SM, Matthews L, Hunter N, Anderson RM: Epidemiology and control of scrapie within a sheep flock. Proc Biol Sci. 1998, 265 (1402): 1205-1210. 10.1098/rspb.1998.0421.
Ryder S, Dexter G, Bellworthy S, Tongue S: Demonstration of lateral transmission of scrapie between sheep kept under natural conditions using lymphoid tissue biopsy. Res Vet Sci. 2004, 76 (3): 211-217. 10.1016/j.rvsc.2003.11.007.
Acknowledgements
We thank the TSE Archive at the VLA (Addlestone, Surrey, UK) for the provision of ruminant brain material, all the staff in the animal services unit and pathology for their contribution in the collection of samples and maintenance of the flock and to Susan Bellworthy for her input at the initiation of this project. This work was funded by the Department for Environment, Food and Rural Affairs, UK.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
LT contribute to the design and coordination of the study and prepared the manuscript. LH carried out all experimental work for method 2. KB carried out all experimental work for method 1. CB carried out method development work for method 1. SE contributed to the coordination of the study and analyses of results from method 2. LTh initiated and designed the PMCA studies. BM contributed to the coordination of the study and analyses of results from method 1. KG contributed to the coordination of the study and analyses of results from method 1 and assisted in manuscript preparation. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Terry, L.A., Howells, L., Bishop, K. et al. Detection of prions in the faeces of sheep naturally infected with classical scrapie. Vet Res 42, 65 (2011). https://doi.org/10.1186/1297-9716-42-65
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1297-9716-42-65