
Abstract
Cryptosporidium spp. is a major cause of diarrheal disease worldwide, particularly in malnourished children and untreated AIDS patients in developing countries in whom it can cause severe, chronic, and debilitating disease. Unfortunately, there is no consistently effective drug for these vulnerable populations and no vaccine, partly due to a limited understanding of both the parasite and the host immune response. In this review, we will discuss our current understanding of the systemic and mucosal immune responses to Cryptosporidium infection, discuss the feasibility of developing a Cryptosporidium vaccine, and evaluate recent advances in Cryptosporidium vaccine development strategies
Similar content being viewed by others


Introduction
Cryptosporidium spp. are intestinal apicomplexan parasites that cause significant diarrheal disease in humans worldwide [1–4]. Diarrhea causes nearly 11 % of deaths in children under the age of five [5], and the recent landmark Global Enteric Multicenter Study (GEMS) case-control study identified Cryptosporidium as one of four pathogens responsible for moderate to severe diarrhea in children in this age group, as well as the second leading cause of diarrheal disease and death in infants in seven countries in South Asia and sub-Saharan Africa [6]. For those who survive, cryptosporidiosis becomes part of a vicious cycle of infection and malnutrition, leading to growth failure, cognitive delays, and physical impairment [7]. In addition, Cryptosporidium infection is a major cause of AIDS-associated diarrhea, with incidence rates reaching as high as 80 % in developing countries [8]. Though infection of immunocompetent hosts is often asymptomatic or self-limiting, Cryptosporidium infection of untreated AIDS patients can lead to unrelenting disease and increased mortality [8]. In industrialized nations, rates of cryptosporidiosis are increasing due to water-borne outbreaks, largely from recreational water sources [9, 10]. Cryptosporidium caused nearly one third of water-borne outbreaks reported in the USA in 2009–2010 [10]. Ease of transmission and dissemination has led to the inclusion of Cryptosporidium as a priority pathogen for biodefense [11]. Yet, in spite of the global impact of cryptosporidiosis, there is no consistently effective therapy for those most affected by the disease [12]. Nitazoxanide, the only FDA-approved drug for cryptosporidiosis, is ineffective in AIDS patients and has not been widely tested in malnourished children [12, 13]. The development of new interventions in these vulnerable populations is urgent, but the inability to continuously propagate or genetically manipulate the parasite in vitro is a major challenge to finding suitable drug targets. Furthermore, an incomplete understanding of the host immune response to Cryptosporidium has impeded development of an effective vaccine strategy. In this review, we will outline recent advances in our understanding of immune responses to Cryptosporidium, discuss the feasibility of developing a Cryptosporidium vaccine, and describe recent advances in vaccine development strategies.
Life Cycle
Before discussing host immune responses and potential intervention targets and strategies, it is important to first understand the life cycle of Cryptosporidium. Infection is initiated by the ingestion of environmentally resistant oocysts, which excyst in the small intestine and release sporozoites, the primary invasive stage [14]. Triggers of excystation are not fully understood but include changes in temperature and pH, as well as the presence of bile salts [15]. Excysted sporozoites then undergo a unique process known as gliding motility in which surface protein translocation is coupled to actin myosin-dependent motility, leading to migration across the surface of epithelial cells [16]. During this process, cell-binding adhesins are secreted from a region within the sporozoites known as the “apical complex,” a collection of secretory organelles—micronemes, dense granules, and a rhoptry—that release proteins in a temperature- and calcium-dependent manner [15, 17]. Cryptosporidium is unique among apicomplexans in that it relies heavily on O-glycosylated mucin-like glycoproteins for attachment to and invasion of host cells [17–21]. These include the circumsporozoite-like glycoprotein CSL [21], gp900 [18], gp40 and gp15 [19], and Muc4 [20], all of which localize to the surface and/or apical region of sporozoites and bind to intestinal epithelial cells in vitro. Following invasion, the parasite undergoes intracellular development within a parasitophorous vacuole composed of both host and parasite components [22]. Small villi-like folds of the parasite cytoplasm extend into the host cell to form a structure similar to a desmosome, called the feeder organelle [22, 23]. It is speculated that the feeder organelle acts as a route of transport between the parasite and host. Within the parasitophorous vacuole, the parasite undergoes both sexual and asexual development [17, 24]. Merozoites released during the asexual cycle can invade neighboring epithelial cells to maintain infection. Microgametes and macrogametes formed during the sexual cycle fuse to form zygotes, which mature into thin- or thick-walled oocysts. Thin-walled oocysts excyst in the small intestine leading to autoinfection, especially in AIDS patients, while thick-walled oocysts are released into the environment.
Immune Responses to Cryptosporidium
The host immune response to Cryptosporidium involves components of both the innate and adaptive immune systems. Several recent reviews have discussed these responses in detail [25–28]. Here, we will review advances made in the past few years.
Innate Immune Responses
Upon ingestion, oocysts and excysted sporozoites first encounter innate immune components along the gastrointestinal tract. Early mediators of innate immune protection include the thick mucus layer of the small intestine, intestinal epithelial cells (IECs), and chemokines, cytokines, and antimicrobial peptides (AMPs) secreted into the intestinal lumen and/or underlying submucosa and bloodstream [29]. Previous studies have elucidated the importance of IECs and soluble mediators during Cryptosporidium infection [30]. IECs provide an initial mechanical and functional barrier [31] and also serve as the primary host cell for Cryptosporidium infection. IEC as well as biliary epithelial cells express several toll-like receptors (TLRs), including TLRs 2, 4, 5 and 9, which have been shown to be important in modulation of the host immune response and subsequent parasite clearance [32–36]. In response to infection, IECs secrete chemokines and cytokines such as IL-8, CXCL10, and CCL2 responsible for the recruitment of inflammatory cells and activation of adaptive immune cells [30, 37, 38], prostaglandins that enhance intestinal fluid secretion [39], and AMPs like β-defensins, which are capable of directly killing sporozoites in vitro [40]. Recently, it was shown that CCL20, a chemokine [41] and AMP [42•] secreted, in part, by IECs in the intestine, was downregulated during Cryptosporidium infection of neonatal mice [43]. Furthermore, oral administration of recombinant CCL20 reduced parasite burden in a manner independent of immune cell recruitment, but rather via direct cytolytic activity on extracellular infective stages of the parasite. CX3CL1 is another chemokine recently shown to be important during Cryptosporidium infection [44]. Its soluble form acts as a potent recruiter of leukocytes, while its membrane-bound form functions as an adhesion molecule for CX3CR1+ T lymphocytes, NK cells, and monocytes [45]. Chen and colleagues found that CX3CL1 expression in biliary epithelial cells was increased in a manner dependent on NF-κB during Cryptosporidium infection in vitro [44]. This effect also extended to a mouse model of Cryptosporidium biliary infection, in which recruitment of CX3CR1+ cells to the biliary tract was increased in livers of infected mice. In addition to chemokines, proinflammatory cytokines secreted by IECs and phagocytes have been shown to play a crucial role in the innate immune response to Cryptosporidium [26, 30]. Lastly, mannose-binding lectin (MBL), a soluble innate immune mediator secreted by hepatocytes, has been shown to be important in the protection against cryptosporidiosis. Several studies have shown that low serum MBL in children and AIDS patients is associated with increased susceptibility to recurrent Cryptosporidium infection [46, 47]. The mechanism by which MBL protects against infection is not fully understood but likely involves complement activation on extracellular stages of the parasite [47].
Early in infection, interferon gamma (IFN-γ) secreted by NK cells, macrophages, and dendritic cells is thought to be the major cytokine involved in orchestrating both the innate and adaptive immune responses [25, 26, 48–50], but recent evidence suggests that IL-18 is important in the control of Cryptosporidium infection as well [51•, 52–54]. IL-18 is secreted by IECs, macrophages, and dendritic cells at sites of infection [53]. Its effects are pleiotropic and include stimulation of IFN-γ and tumor necrosis factor alpha (TNF-α) production by immune cells, chemotaxis of inflammatory cells, maintenance of epithelial integrity, and stimulation of AMP secretion by IECs [51•, 52, 53, 55]. How IL-18 protects against Cryptosporidium infection is not fully understood, but it likely involves multiple effector functions. McDonald and colleagues previously found that IL-18 might reduce Cryptosporidium infection via enhancement of secretion of AMPs by IECs [52]. More recently, they found that IL-18 confers protection against Cryptosporidium parvum infection in vivo by coordinating with IL-12 to enhance IFN- γ production by macrophages [53]. These results were supported by studies done by Mead and colleagues, who found that IL-18 protected against C. parvum infection in vivo via stimulation of IFN-γ production and AMP expression [51•]. They also found that mouse dendritic cells were able to produce IL-18 upon stimulation with C. parvum antigens [56]. It is unclear whether the effects of IL-18 on IFN-γ and AMP production are mutually exclusive or whether IL-18-dependent stimulation of AMP production by IECs is due to enhanced IFN-γ secretion.
Upon their interaction with Cryptosporidium, IECs relay signals to innate immune effector cells [25, 26, 38, 57, 58]. These include dendritic cells, NK cells, macrophages, neutrophils mast cells, and eosinophils. Dendritic cells migrate toward areas of C. parvum infection in an IFN- γ-dependent manner [38], but until recently, their role in clearance of the parasite was unclear. In vitro studies found that bone marrow-derived dendritic cells challenged with C. parvum sporozoites or antigens secreted a number of cytokines, including type I IFN, TNF-α, IL-6, IL-1β, IL-12, and IL-18 [36, 56, 59]. More recent studies have elucidated the importance of dendritic cells in vivo. Mead and colleagues found that depletion of dendritic cells in adult mice significantly increased oocyst shedding and intestinal pathology [60]. Furthermore, parasite burden was reduced upon adoptive transfer of dendritic cells, with longer protection observed in mice receiving dendritic cells stimulated with live parasite in vitro prior to transfer. Laurent and colleagues extended this finding by evaluating the role of dendritic cells during C. parvum infection in neonatal mice [61••], which are more susceptible to infection compared to adults. They found that neonatal mice had significantly lower numbers of intestinal CD103+ dendritic cells during the first weeks of life, when the mice were most susceptible to infection. Stimulation of dendritic cell production during this period led to increased resistance to infection through a mechanism dependent on IL-12 and IFN-γ and independent of adaptive responses. Recently, dendritic cells were shown to transport Cryptosporidium parasites and antigens to mesenteric lymph nodes in mice [36], possibly to activate adaptive immune cells, though it is not clear whether antigens are able to cross the parasitophorous vacuolar membrane for antigen processing and presentation. Together, these studies suggest that dendritic cells not only play a role in the clearance of Cryptosporidium infection, but may also be necessary during the activation of the adaptive immune response. Furthermore, their absence early in development may contribute to the increased susceptibility to mucosal infections found in neonates.
NK cells serve as a major source of IFN-γ early in infection (reviewed in [26]). They are activated by IL-15 secreted by IECs and have been shown to lyse C. parvum-infected IECs in response to IL-15 in vitro [62]. Previous studies found that mice lacking functional NK cells were more susceptible to Cryptosporidium infection [63, 64], while treatment of immunocompetent and immunodeficient mice with IL-12, a potent NK cell activator, enhanced protection [65]. Both studies found that NK cell-dependent protection was primarily mediated through IFN-γ. Drouet and colleagues recently found an increase in early recruitment of activated, perforin + NK cells to areas of infection in neonatal lambs infected with C. parvum [66•]. These studies suggest that the role of NK cells during Cryptosporidium infection may involve both IFN-γ-mediated protection and direct cytolysis.
In addition to dendritic and NK cells, mast cells may be necessary for parasite clearance [58], but their role during Cryptosporidium infection remains poorly understood. Mast cells play a pivotal role in bacterial and parasitic infections [67, 68]. They secrete inflammatory mediators (histamine, cytokines, prostagalandins, leukotrienes) locally and systemically, mobilize and recruit innate and adaptive immune components, and generally influence an overall Th2-type immune response [67, 68]. In mice infected with Cryptosporidium muris, accumulation of gastric mucosal mast cells was shown to correlate with oocyst shedding, suggesting a role in parasite clearance [69]. Similarly, Zhang and colleagues found an association between intestinal mucosal mast cell accumulation and C. parvum infection in calves [70], with increased recruitment associated with enhanced clearance of the parasite. Mast cells may also contribute to jejunal hypersensitivity in rats infected with C. parvum [71]. Still, most of these studies have focused on evaluating a correlation between mast cell recruitment and parasite clearance and pathology. Future studies should aim to elucidate whether Cryptosporidium can trigger mast cell degranulation and whether mast cell depletion or inhibition of degranulation affects parasite clearance. It would also be interesting to evaluate whether mast cells influence the Th1/Th2 balance during infection, as Cryptosporidium generally induces a Th1-type response [25, 26, 72].
Advances in genomic research have uncovered another crucial component of the innate immune response against Cryptosporidium—noncoding RNAs (ncRNAs) [73]. In particular, recent studies have found significant alterations in TLR4- and NF-κB-dependent microRNA (miRNA) regulation in epithelial cells infected with C. parvum [74]. These studies also found that inhibition of select miRNAs led to enhanced infection in vitro, suggesting a direct link between miRNA regulation and host cell protection against Cryptosporidium. Relevant functional targets include KH-type splicing regulatory protein (KSRP) [75], SNAP23 [76•], CX3CL1 [44], CCL20 [43], and ICAM-1 [77]. KSRP is an RNA-binding protein that regulates messenger RNA (mRNA) decay of several important immune mediators, including inducible nitric oxide synthase (iNOS) [78]. Recently, Chen and colleagues discovered that the miRNA miR-27b directly downregulated KSRP translation during C. parvum infection in vitro, which led to increased iNOS mRNA stability and NO production [75]. Inhibition of miR-27b led to an increase in infection due to upregulation of KSRP translation and enhanced iNOS decay. Another study by this group found that downregulation of let-7 miRNAs during C. parvum infection led to increased SNAP23 expression and a subsequent increase in exosome release from biliary epithelial cells [76•]. These exosomes, which carry immune effector molecules like AMPs, were able to directly interact with C. parvum sporozoites to reduce their viability and infectivity. This group also found that another miRNA, miR-221, regulated ICAM-1 expression during C. parvum infection, possibly as a means to enhance lymphocyte homing to sites of infection [77]. Lastly, two chemokines mentioned previously, CX3CL1 and CCL20, were also found to be regulated by miRNAs during C. parvum infection [43, 44]. CX3CL1 is negatively regulated by miR-424 and miR-503, whereas CCL20 is negatively regulated by miR-21. Interestingly, miR-21 was upregulated during C. parvum infection, which led to downregulation of CCL20 and an increase in parasite burden. This suggests that Cryptosporidium has developed ways of dampening the innate immune response via the regulation of miRNAs.
Adaptive Immune Responses
The importance of the adaptive immune response during Cryptosporidium infection is highlighted by the susceptibility of AIDS patients to cryptosporidiosis, as well as the resolution of infection observed following CD4+ T cell reconstitution in patients given antiretroviral therapy [8, 25]. Though disease severity in humans is often viewed as being inversely proportional to absolute CD4+ T cell numbers, Tzipori and colleagues recently found that persistent cryptosporidiosis in macaques was more dependent on SIV load, viral damage to gut lymphoid tissue, and rapid depletion of mucosal CD4+ T cells during the acute phase of viral infection than on declining circulating CD4+ T cell levels during chronic SIV infection [79], suggesting that depletion of local CD4+ T cells may be more predictive of disease severity than absolute CD4+ T cell numbers.
Much of our understanding of the cell-mediated immune response to Cryptosporidium infection is limited to CD4+ T cells; the role and importance of CD8+ T cells are less clear [50]. Previous studies found that CD8+ T cell numbers increased during Cryptosporidium infection of macaques [79], and both CD4+ and CD8+ T cells isolated from humans with previous Cryptosporidium infection could produce IFN-γ in response to stimulation by Cryptosporidium hominis antigens [80]. Furthermore, in vitro studies found that CD8+ T cells isolated from donors with prior exposure to Cryptosporidium were able to lyse C. parvum-infected IECs in a manner dependent on the release of cytotoxic granules [81]. The importance of CD8+ T cells in vivo was recently studied by Salát and colleagues [82], who found that reconstitution of immunocompromised mice with activated CD8+ T cells significantly reduced the length and severity of C. muris infection, albeit to a lesser extent than reconstitution with CD4+ T cells. Regardless, these studies suggest that CD8+ T cells contribute to the cell-mediated immune response to Cryptosporidium, likely via direct cytolysis of infected IECs and through IFN-γ-mediated protection and clearance.
The role of humoral immunity during Cryptosporidium infection is incompletely understood. Passive immunization studies in animal models have shown a correlation between anti-Cryptosporidium antibody administration and reductions in oocyst shedding and disease severity (reviewed in [50, 83]). The use of hyperimmune bovine colostrum for passive immunotherapy of cryptosporidiosis in humans has also been evaluated, with variable results [50, 83]. Numerous studies in humans have found an association between levels of anti-Cryptosporidium antibodies and history of infection [25, 84–87, 88••, 89]. Our studies found significantly increased IgG, IgM, and IgA responses to C. parvum p23 and increased IgG and IgA responses to C. parvum and C. hominis gp15 in Bangladeshi children infected primarily with C. hominis [86, 87], suggesting a cross-reactive humoral response to gp15. We found a similar phenomenon in anti-gp15 antibody responses in South Indian children infected with Cryptosporidium [84]. Interestingly, several studies found that acute and asymptomatic cryptosporidiosis were associated with higher IgG, IgM, and/or IgA responses to Cryptosporidium spp. antigens compared to persistent cryptosporidiosis [86, 87, 88••], suggesting that humoral immunity may play a role in limiting the length and severity of infection. However, it is also likely that the association between anti-Cryptosporidium antibody responses and disease may reflect underlying cell-mediated immune responses. We found that not only did HIV-infected patients with asymptomatic cryptosporidiosis have higher circulating IgG and fecal IgA levels to Cryptosporidium antigens compared to patients with diarrhea, but these patients, on average, also had significantly higher CD4+ T cell counts [88••], which is known to be more predictive of disease severity. Future studies should aim to distinguish between the correlative and causative effects of the humoral response during Cryptosporidium infection.
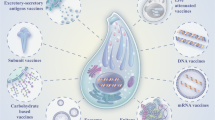
Vaccine Development
Considerations and Concerns
Before discussing potential vaccine candidates, it is important to first consider the feasibility of developing an effective vaccine against Cryptosporidium. The ideal vaccine should provide rapid life-long immunity in all vaccinated individuals, be broadly protective against the most common species and subtypes of the organism, prevent disease transmission, and be readily accessible, stable, and cheap (reviewed in [28, 90]). Multiple Cryptosporidium species and subtypes can infect vulnerable populations, primarily malnourished children and immunocompromised individuals in developing countries. An effective Cryptosporidium vaccine should elicit a strong cross-protective mucosal immune response to subtypes of both C. parvum and C. hominis, the two most common species infecting humans. Although, all subtypes of C. hominis are anthroponotic, C. parvum displays anthroponotic as well as zoonotic subtypes. C. parvum has a broad host range, and host specificity varies by species subtype [91]. A recent study showed that infection of gnotobiotic pigs with C. hominis led to complete protection against subsequent infection with C. hominis, but incomplete protection against infection with C. parvum [92], suggesting that targeting a single species alone is not sufficient to provide cross-protection against different species. Similarly, we showed that immune responses to the polymorphic gp40 antigen are, in part, subtype-specific in infected children (Ajjampur et al. 2011). Therefore, vaccine development should focus on targeting those subtypes most common in human infections. The vaccine should also be effective in children and immunocompromised individuals and cheap and accessible to populations in developing countries. To date, there are no FDA-approved parasite vaccines, likely due to the greater complexity of these organisms compared to bacteria and viruses. Though several studies suggest that humoral cross-reactivity between species exists [84, 86, 87], whether this is associated with cross-protection is unknown. Use of a live, attenuated form of Cryptosporidium may be required to elicit a strong mucosal immune response (reviewed in [28]). Though this is generally not an issue in immunocompetent individuals, live, attenuated organisms may still cause disease or transmit infection in patients with underlying immune disorders. Lastly, populations most affected by cryptosporidiosis may not have the capacity to develop robust, sustained immune-mediated protection in response to vaccination. Therefore, the use of adjuvants may be necessary to enhance immune responses in these populations. TLR ligands have been used as adjuvants in numerous vaccine trials, with many providing enhanced protection compared to vaccination with antigen alone [93]. In addition, several studies have shown that administration of TLR agonists in mice infected with Cryptosporidium leads to a more robust immune response and accelerated parasite clearance [33, 35], supporting their feasibility as effective vaccine adjuvants.
Cryptosporidium Antigens and Putative Vaccines
In order to elicit a protective immune response, vaccines must contain antigens exposed to the host immune system; this may involve use of a single protein, multiple antigens, or the entire organism. As discussed previously, use of a live, attenuated Cryptosporidium vaccine may still cause disease in immunocompromised populations. Its development is also challenging due to an inability to continuously propagate or genetically manipulate the parasite in vitro. Therefore, most studies have focused on the immunogenic potential of specific proteins involved in attachment to and invasion of host cells, such as CSL, gp900, gp40, gp15/17, Cp15/60, Cp23/27, Cp12, and Muc4 (reviewed in [28, 83]). Antibodies targeting these proteins can inhibit Cryptosporidium infection in vitro [17, 20, 94], and humoral responses to several of these antigens are elevated in patients with cryptosporidiosis [25, 84–87, 88••, 89]. A few of these antigens such as gp900, gp40, gp15/17, and Cp23/27 have been shown to induce cellular immune responses in animals or humans [56, 80, 95]. However, it is not known whether these responses are protective. This is critical, since protective immunity to Cryptosporidium is mostly cell-mediated.
The type of vaccine is another important consideration. Broadly, Cryptosporidium vaccines can be divided into three categories: protein antigens, DNA-based vaccines, and antigens expressed in bacterial and parasite vectors (reviewed in [28]). DNA-based methods involve administration of a plasmid encoding a particular antigen. The plasmid is taken up by host cells, and the antigen is expressed and exposed to the immune system. Several recent studies have evaluated DNA-based vaccines for Cryptosporidium P2, Cp12, Cp15/60, Cp23, Cp21, and COWP [96–99, 100•]. He and colleagues recently compared the efficacy of univalent and divalent DNA-based vaccines containing Cp15 and/or Cp23 [97]. Infected mice immunized with the divalent construct developed stronger immune responses and shed fewer oocysts than mice given the univalent vaccine. Similarly, Huang and colleagues found that a divalent DNA-based vaccine containing Cp15 and Cp23 elicited more protection in infected mice than immunization with a single antigen or crude protein [100•], suggesting that immunization with multiple antigens enhances protection. Still, DNA-based vaccines have several disadvantages, including poor immunogenicity, improper processing of foreign antigens, and the potential of host cell transformation [101]. Bacterial vectors offer the advantage of inducing strong systemic and mucosal immune responses but must be attenuated to prevent potential damage to the host [28]. Attenuated Salmonella vaccines have been used for several Cryptosporidium antigens [28]. Mead and colleagues showed that oral administration of a Salmonella serovar Typhi vector encoding Cp23 and Cp40 led to specific immune responses against these antigens [102]. More recently, Galen, Buck, Guerrant, and colleagues developed a Salmonella serovar Typhi vector containing Cp15 and evaluated its efficacy via intranasal administration in well-nourished and malnourished mice infected with Cryptosporidium [103•, 104, 105••]. Unfortunately, disease course was not affected by immunization. Both well-nourished and malnourished immunized mice developed strong systemic immune responses, but only well-nourished mice developed a robust local immune response, suggesting that malnutrition negatively impacts the mucosal immune response during infection or immunization—an important finding considering the significant burden of Cryptosporidium in malnourished children. Another study found that mice immunized with a Lactobacillus casei bacterial vector containing C. parvum p23/27 developed systemic and mucosal immune responses against Cryptosporidium [106]. Together, these studies support the use of bacterial vectors for the production of robust systemic and local immune responses to Cryptosporidium antigens. Parasite vectors expressing Cryptosporidium antigens offer the additional advantage of post-translational modifications and presenting of antigens similar to native Cryptosporidium proteins. Toxoplasma gondii, an apicomplexan parasite similar to Cryptosporidium, is commonly used as a heterologous expression system for other apicomplexans [107] and could therefore serve as an effective vector for Cryptosporidium antigens. C. parvum gp40, gp15, and p23 have been successfully expressed in T. gondii and display similar post-translational modifications as the native proteins [108, 109]. Unfortunately, though it has shown promise as a Cryptosporidium vaccine in animal models [109], the disease risks associated with infecting humans with T. gondii are too great [28]. Use of attenuated or avirulent strains of T. gondii or immunization with purified recombinant proteins expressed in T. gondii may be explored in the future.
Though most vaccine studies have focused on a few Cryptosporidium antigens, other promising candidates likely exist. In order to identify new potential vaccine targets, researchers should focus on ways to screen for proteins that are necessary for infection and can induce protective cell-mediated immune responses. It is important to note that many of the proteins implicated in Cryptosporidium attachment and invasion undergo extensive post-translational modification, particularly glycosylation [17]. These modifications may be necessary for the function and immunogenic potential of the protein and, therefore, are important to consider when designing a screen for Cryptosporidium antigens. A phage-display C. parvum complementary DNA (cDNA) library was used to identify the surface antigen Cp12 and the immunodominant acidic ribosome protein P2 [110, 111]. Zhang and colleagues recently utilized a cell-free ribosome display platform to screen a C. parvum cDNA library for proteins that could adhere to IECs [112]. They discovered a novel protein, Cp20, which localized to the surface of both C. parvum sporozoites and oocysts. Mice immunized with a DNA vaccine containing the Cp20 sequence developed strong cell-mediated and humoral immune responses against Cryptosporidium and shed fewer oocysts than unvaccinated mice, suggesting that Cp20 may serve as an effective vaccine target. Another promising screening technique is the use of “reverse vaccinology,” in which the Cryptosporidium proteome is mined in silico in order to identify potential surface proteins exposed to the immune system during infection. Buck and colleagues recently utilized this technique to identify several well-known and novel Cryptosporidium vaccine candidates, including Cp15, profilin, and calcium apyrase [105••]. Mice vaccinated with a live Salmonella vector expressing these proteins developed strong, specific cell-mediated and humoral responses, suggesting that these antigens may elicit a protective immune response against Cryptosporidium and thus serve as effective vaccine targets. Additional studies should aim to determine whether immunization with these antigens also provides protection from Cryptosporidium infection.
Conclusions
The host immune response to Cryptosporidium infection involves components of both the innate and adaptive immune systems. IECs provide a first line of defense, followed by the recruitment of innate immune cells such as NK cells, dendritic cells, macrophages, and mast cells. Together, they secrete AMPs, inflammatory mediators, chemokines, and cytokines and activate and mobilize cells of the adaptive immune system. CD4+ T cells and IFN-γ are essential to the clearance of Cryptosporidium infection, but CD8+ T cells and humoral responses may play a role as well. Still, our understanding of the immune response to Cryptosporidium is limited, which provides a major obstacle to the development of immune-based interventions. Numerous Cryptosporidium proteins mediate infection and are immunogenic in humans, but few have shown promise as effective vaccine candidates, partly because we do not fully understand which immune responses are necessary for protection. Another obstacle is our inability to propagate or genetically manipulate the parasite in vitro, which hinders our search for and validation of other potential vaccine candidates. Clearly, the development of an effective vaccine against Cryptosporidium is a challenge—one that will require significant advancements in our understanding of both the parasite and the host immune response.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Striepen B. Parasitic infections: time to tackle cryptosporidiosis. Nature. 2013;503(7475):189–91.
Leav BA, Mackay M, Ward HD. Cryptosporidium species: new insights and old challenges. Clin Infect Dis. 2003;36(7):903–8.
Collinet-Adler S, Ward HD. Cryptosporidiosis: environmental, therapeutic, and preventive challenges. Eur J Clin Microbiol Infect Dis. 2010;29(8):927–35.
Checkley W, White Jr AC, Jaganath D, Arrowood MJ, Chalmers RM, Chen X, et al. A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for cryptosporidium. Lancet Infect Dis. 2015;15(1):85–94.
Liu L, Oza S, Hogan D, Perin J, Rudan I, Lawn JE, et al. Global, regional, and national causes of child mortality in 2000–13, with projections to inform post-2015 priorities: an updated systematic analysis. Lancet. 2015;385(9966):430–40.
Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case–control study. Lancet. 2013;382(9888):209–22.
Shirley DA, Moonah SN, Kotloff KL. Burden of disease from cryptosporidiosis. Curr Opin Infect Dis. 2012;25(5):555–63.
O’Connor RM, Shaffie R, Kang G, Ward HD. Cryptosporidiosis in patients with HIV/AIDS. AIDS. 2011;25(5):549–60.
Hlavsa MC, Roberts VA, Anderson AR, Hill VR, Kahler AM, Orr M, et al. Surveillance for waterborne disease outbreaks and other health events associated with recreational water—United States, 2007–2008. MMWR Surveill Summ. 2011;60(12):1–32.
Hlavsa MC, Roberts VA, Kahler AM, Hilborn ED, Wade TJ, Backer LC, et al. Recreational water-associated disease outbreaks—United States, 2009–2010. MMWR Morb Mortal Wkly Rep. 2014;63(1):6–10.
Rotz LD, Khan AS, Lillibridge SR, Ostroff SM, Hughes JM. Public health assessment of potential biological terrorism agents. Emerg Infect Dis. 2002;8(2):225–30.
Cabada MM, White Jr AC. Treatment of cryptosporidiosis: do we know what we think we know? Curr Opin Infect Dis. 2010;23(5):494–9.
Abubakar I, Aliyu SH, Arumugam C, Hunter PR, Usman NK. Prevention and treatment of cryptosporidiosis in immunocompromised patients. Cochrane Database Syst Rev. 2007;1:CD004932.
Tzipori S, Ward H. Cryptosporidiosis: biology, pathogenesis and disease. Microbes Infect. 2002;4(10):1047–58.
Smith HV, Nichols RA, Grimason AM. Cryptosporidium excystation and invasion: getting to the guts of the matter. Trends Parasitol. 2005;21(3):133–42.
Wetzel DM, Schmidt J, Kuhlenschmidt MS, Dubey JP, Sibley LD. Gliding motility leads to active cellular invasion by Cryptosporidium parvum sporozoites. Infect Immun. 2005;73(9):5379–87.
Wanyiri J, Ward H. Molecular basis of Cryptosporidium-host cell interactions: recent advances and future prospects. Future Microbiol. 2006;1:201–8.
Barnes DA, Bonnin A, Huang JX, Gousset L, Wu J, Gut J, et al. A novel multi-domain mucin-like glycoprotein of Cryptosporidium parvum mediates invasion. Mol Biochem Parasitol. 1998;96(1–2):93–110.
Cevallos AM, Zhang X, Waldor MK, Jaison S, Zhou X, Tzipori S, et al. Molecular cloning and expression of a gene encoding Cryptosporidium parvum glycoproteins gp40 and gp15. Infect Immun. 2000;68(7):4108–16.
O’Connor RM, Burns PB, Ha-Ngoc T, Scarpato K, Khan W, Kang G, et al. Polymorphic mucin antigens CpMuc4 and CpMuc5 are integral to Cryptosporidium parvum infection in vitro. Eukaryot Cell. 2009;8(4):461–9.
Riggs MW, Stone AL, Yount PA, Langer RC, Arrowood MJ, Bentley DL. Protective monoclonal antibody defines a circumsporozoite-like glycoprotein exoantigen of Cryptosporidium parvum sporozoites and merozoites. J Immunol. 1997;158(4):1787–95.
Tzipori S, Griffiths JK. Natural history and biology of Cryptosporidium parvum. Adv Parasitol. 1998;40:5–36.
Perkins ME, Riojas YA, Wu TW, Le Blancq SM. CpABC, a Cryptosporidium parvum ATP-binding cassette protein at the host-parasite boundary in intracellular stages. Proc Natl Acad Sci U S A. 1999;96(10):5734–9.
O’Hara SP, Chen XM. The cell biology of cryptosporidium infection. Microbes Infect. 2011.
Borad A, Ward H. Human immune responses in cryptosporidiosis. Future Microbiol. 2010;5:507–19.
McDonald V, Korbel DS, Barakat FM, Choudhry N, Petry F. Innate immune responses against Cryptosporidium parvum infection. Parasite Immunol. 2013;35(2):55–64.
Wilhelm CL, Yarovinsky F. Apicomplexan infections in the gut. Parasite Immunol. 2014;36(9):409–20.
Mead JR. Prospects for immunotherapy and vaccines against Cryptosporidium. Hum Vaccine Immunotherapeut. 2014;10(6):1505–13.
Jr. RBJ. An overview of the innate immune system. 2015.
Pantenburg B, Dann SM, Wang HC, Robinson P, Castellanos-Gonzalez A, Lewis DE, et al. Intestinal immune response to human Cryptosporidium sp. infection. Infect Immun. 2008;76(1):23–9.
Peterson LW, Artis D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol. 2014;14(3):141–53.
O’Hara S, Tietz Bogert PS, Trussoni CE, Chen X, Larusso NF. Tlr4 Promotes Cryptosporidium parvum clearance in a mouse model of biliary cryptosporidiosis. J Parasitol. 2011.
Barrier M, Lacroix-Lamande S, Mancassola R, Auray G, Bernardet N, Chausse AM, et al. Oral and intraperitoneal administration of phosphorothioate oligodeoxynucleotides leads to control of cryptosporidium parvum infection in neonatal mice. J Infect Dis. 2006;193(10):1400–7.
Costa LB, Johnbull EA, Reeves JT, Sevilleja JE, Freire RS, Hoffman PS, et al. Cryptosporidium- malnutrition interactions: mucosal disruption, cytokines and TLR signaling in a weaned murine model. J Parasitol. 2011.
Lantier L, Drouet F, Guesdon W, Mancassola R, Metton C, Lo-Man R, et al. Poly(I:C)-induced protection of neonatal mice against intestinal Cryptosporidium parvum infection requires an additional TLR5 signal provided by the gut flora. J Infect Dis. 2014;209(3):457–67.
Perez-Cordon G, Yang G, Zhou B, Nie W, Li S, Shi L, et al. Interaction of Cryptosporidium parvum with mouse dendritic cells leads to their activation and parasite transportation to mesenteric lymph nodes. Pathogens Dis. 2014;70(1):17–27.
Laurent F, Eckmann L, Savidge TC, Morgan G, Theodos C, Naciri M, et al. Cryptosporidium parvum infection of human intestinal epithelial cells induces the polarized secretion of C-X-C chemokines. Infect Immun. 1997;65(12):5067–73.
Auray G, Lacroix-Lamande S, Mancassola R, Dimier-Poisson I, Laurent F. Involvement of intestinal epithelial cells in dendritic cell recruitment during C. parvum infection. Microbes Infect. 2007;9(5):574–82.
Laurent F, Kagnoff MF, Savidge TC, Naciri M, Eckmann L. Human intestinal epithelial cells respond to Cryptosporidium parvum infection with increased prostaglandin H synthase 2 expression and prostaglandin E2 and F2alpha production. Infect Immun. 1998;66(4):1787–90.
Carryn S, Schaefer DA, Imboden M, Homan EJ, Bremel RD, Riggs MW. Phospholipases and cationic peptides inhibit cryptosporidium parvum sporozoite infectivity by parasiticidal and non-parasiticidal mechanisms. J Parasitol. 2012;98(1):199–204.
Schutyser E, Struyf S, Van Damme J. The CC chemokine CCL20 and its receptor CCR6. Cytokine Growth Factor Rev. 2003;14(5):409–26.
Yang D, Chen Q, Hoover DM, Staley P, Tucker KD, Lubkowski J, et al. Many chemokines including CCL20/MIP-3alpha display antimicrobial activity. J Leukoc Biol. 2003;74(3):448–55. This study identified a novel AMP able to target Cryptosporidium directly. Furthermore, their results suggest that Cryptosporidium is able to counteract CCL20 via miRNA-mediated downregulation.
Guesdon W, Auray G, Pezier T, Bussiere FI, Drouet F, Le Vern Y, et al. CCL20 displays antimicrobial activity against Cryptosporidium parvum but its expression is reduced during infection in the intestine of neonatal mice. J Infect Dis. 2015.
Zhou R, Gong AY, Chen D, Miller RE, Eischeid AN, Chen XM. Histone deacetylases and NF-kB signaling coordinate expression of CX3CL1 in epithelial cells in response to microbial challenge by suppressing miR-424 and miR-503. PLoS One. 2013;8(5):e65153.
Stievano L, Piovan E, Amadori A. C and CX3C chemokines: cell sources and physiopathological implications. Crit Rev Immunol. 2004;24(3):205–28.
Carmolli M, Duggal P, Haque R, Lindow J, Mondal D, Petri Jr WA, et al. Deficient serum mannose-binding lectin levels and MBL2 polymorphisms increase the risk of single and recurrent Cryptosporidium infections in young children. J Infect Dis. 2009;200(10):1540–7.
Kelly P, Jack DL, Naeem A, Mandanda B, Pollok RC, Klein NJ, et al. Mannose-binding lectin is a component of innate mucosal defense against Cryptosporidium parvum in AIDS. Gastroenterology. 2000;119(5):1236–42.
Lean IS, McDonald V, Pollok RC. The role of cytokines in the pathogenesis of Cryptosporidium infection. Curr Opin Infect Dis. 2002;15(3):229–34.
Theodos CM. Innate and cell-mediated immune responses to Cryptosporidium parvum. Adv Parasitol. 1998;40:87–119.
Riggs MW. Recent advances in cryptosporidiosis: the immune response. Microbes Infect. 2002;4(10):1067–80.
Bedi B, McNair NN, Forster I, Mead JR. IL-18 cytokine levels modulate innate immune responses and cryptosporidiosis in mice. J Eukaryot Microbiol. 2015;62(1):44–50. This study evaluated the protective effect of IL-18 in vivo and discusses potential mechanisms.
McDonald V, Pollok RC, Dhaliwal W, Naik S, Farthing MJ, Bajaj-Elliott M. A potential role for interleukin-18 in inhibition of the development of Cryptosporidium parvum. Clin Exp Immunol. 2006;145(3):555–62.
Choudhry N, Petry F, van Rooijen N, McDonald V. A protective role for interleukin 18 in interferon gamma-mediated innate immunity to Cryptosporidium parvum that is independent of natural killer cells. J Infect Dis. 2012;206(1):117–24.
Ehigiator HN, McNair N, Mead JR. Cryptosporidium parvum: the contribution of Th1-inducing pathways to the resolution of infection in mice. Exp Parasitol. 2007;115(2):107–13.
Biet F, Locht C, Kremer L. Immunoregulatory functions of interleukin 18 and its role in defense against bacterial pathogens. J Mol Med. 2002;80(3):147–62.
Bedi B, Mead JR. Cryptosporidium parvum antigens induce mouse and human dendritic cells to generate Th1-enhancing cytokines. Parasite Immunol. 2012;34(10):473–85.
Codices V, Martins C, Novo C, Pinho M, de Sousa B, Lopes A, et al. Cell phenotypic change due to Cryptosporidium parvum infection in immunocompetent mice. Acta Parasitol / Witold Stefanski Inst Parasitol, Warszawa, Poland. 2013;58(1):70–9.
Harp JA, Moon HW. Susceptibility of mast cell-deficient W/Wv mice to Cryptosporidium parvum. Infect Immun. 1991;59(2):718–20.
Barakat FM, McDonald V, Foster GR, Tovey MG, Korbel DS. Cryptosporidium parvum infection rapidly induces a protective innate immune response involving type I interferon. J Infect Dis. 2009;200(10):1548–55.
Bedi B, McNair NN, Mead JR. Dendritic cells play a role in host susceptibility to Cryptosporidium parvum infection. Immunol Lett. 2014;158(1–2):42–51.
Lantier L, Lacroix-Lamande S, Potiron L, Metton C, Drouet F, Guesdon W, et al. Intestinal CD103+ dendritic cells are key players in the innate immune control of Cryptosporidium parvum infection in neonatal mice. PLoS Pathog. 2013;9(12):e1003801. This study suggests that dendritic cells are essential to clearance of Cryptosporidium infection in neonates, an important consideration for vaccine development.
Dann SM, Wang HC, Gambarin KJ, Actor JK, Robinson P, Lewis DE, et al. Interleukin-15 activates human natural killer cells to clear the intestinal protozoan cryptosporidium. J Infect Dis. 2005;192(7):1294–302.
Barakat FM, McDonald V, Di Santo JP, Korbel DS. Roles for NK cells and an NK cell-independent source of intestinal gamma interferon for innate immunity to Cryptosporidium parvum infection. Infect Immun. 2009;77(11):5044–9.
Korbel DS, Barakat FM, Di Santo JP, McDonald V. CD4+ T cells are not essential for control of early acute Cryptosporidium parvum infection in neonatal mice. Infect Immun. 2011;79(4):1647–53.
Urban Jr JF, Fayer R, Chen SJ, Gause WC, Gately MK, Finkelman FD. IL-12 protects immunocompetent and immunodeficient neonatal mice against infection with Cryptosporidium parvum. J Immunol. 1996;156(1):263–8.
Olsen L, Akesson CP, Storset AK, Lacroix-Lamande S, Boysen P, Metton C, et al. The early intestinal immune response in experimental neonatal ovine cryptosporidiosis is characterized by an increased frequency of perforin expressing NCR1 NK cells and by NCR1 CD8 cell recruitment. Vet Res. 2015;46(1):28. This study suggests that NK cells may contribute to mucosal protection via direct cytolysis.
Krishnaswamy G, Kelley J, Johnson D, Youngberg G, Stone W, Huang SK, et al. The human mast cell: functions in physiology and disease. FronT Biosci: J Virt Lib. 2001;6:D1109–27.
Abraham SN, St John AL. Mast cell-orchestrated immunity to pathogens. Nat Rev Immunol. 2010;10(6):440–52.
Rhee JK, Surl CG, Kim HC. Effects of Cryptosporidium muris (strain MCR) infection on gastric mucosal mast cells in mice. Korean J Parasitol. 1997;35(4):245–9.
Li S, Li W, Yang Z, Song S, Yang J, Gong P, et al. Infection of cattle With Cryptosporidium parvum: mast cell accumulation in small intestine mucosa. Vet Pathol. 2013.
Bai J, Khaldi S, Gargala G, Baishanbo A, Francois A, Ballet JJ, et al. Effects of octreotide on jejunal hypersensitivity triggered by Cryptosporidium parvum intestinal infection in an immunocompetent suckling rat model. Neurogastroenterol Motil. 2011.
Codices V, Martins C, Novo C, de Sousa B, Lopes A, Borrego M, et al. Dynamics of cytokines and immunoglobulins serum profiles in primary and secondary Cryptosporidium parvum infection: usefulness of Luminex(R) xMAP technology. Exp Parasitol. 2013;133(1):106–13.
Zhou R, Feng Y, Chen XM. Non-coding RNAs in epithelial immunity to Cryptosporidium infection. Parasitology. 2014;141(10):1233–43.
Zhou R, Hu G, Liu J, Gong AY, Drescher KM, Chen XM. NF-kappaB p65-dependent transactivation of miRNA genes following Cryptosporidium parvum infection stimulates epithelial cell immune responses. PLoS Pathog. 2009;5(12):e1000681.
Zhou R, Gong AY, Eischeid AN, Chen XM. miR-27b targets KSRP to coordinate TLR4-mediated epithelial defense against Cryptosporidium parvum infection. PLoS Pathog. 2012;8(5):e1002702.
Hu G, Gong AY, Roth AL, Huang BQ, Ward HD, Zhu G, et al. Release of luminal exosomes contributes to TLR4-mediated epithelial antimicrobial defense. PLoS Pathog. 2013;9(4):e1003261. This study evaluates miRNA regulation of exosome release in the context of Cryptosporidium infection.
Gong AY, Hu G, Zhou R, Liu J, Feng Y, Soukup GA, et al. MicroRNA-221 controls expression of intercellular adhesion molecule-1 in epithelial cells in response to Cryptosporidium parvum infection. Int J Parasitol. 2011;41(3–4):397–403.
Linker K, Pautz A, Fechir M, Hubrich T, Greeve J, Kleinert H. Involvement of KSRP in the post-transcriptional regulation of human iNOS expression-complex interplay of KSRP with TTP and HuR. Nucleic Acids Res. 2005;33(15):4813–27.
Singh I, Carville A, Tzipori S. Cryptosporidiosis in rhesus macaques challenged during acute and chronic phases of SIV infection. AIDS Res Hum Retroviruses. 2011.
Preidis GA, Wang HC, Lewis DE, Castellanos-Gonzalez A, Rogers KA, Graviss EA, et al. Seropositive human subjects produce interferon gamma after stimulation with recombinant Cryptosporidium hominis gp15. Am J Trop Med Hyg. 2007;77(3):583–5.
Pantenburg B, Castellanos-Gonzalez A, Dann SM, Connelly RL, Lewis DE, Ward HD, et al. Human CD8(+) T cells clear Cryptosporidium parvum from infected intestinal epithelial cells. Am J Trop Med Hyg. 2010;82(4):600–7.
Kvac M, Kodadkova A, Sak B, Kvetonova D, Jalovecka M, Rost M, et al. Activated CD8+ T cells contribute to clearance of gastric Cryptosporidium muris infections. Parasite Immunol. 2011;33(4):210–6.
Boulter-Bitzer JI, Lee H, Trevors JT. Molecular targets for detection and immunotherapy in Cryptosporidium parvum. Biotechnol Adv. 2007;25(1):13–44.
Ajjampur SS, Sarkar R, Allison G, Banda K, Kane A, Muliyil J, et al. Serum IgG response to cryptosporidium immunodominant antigen gp15 and polymorphic antigen gp40 in children with cryptosporidiosis in South India. Clin Vaccine Immunol. 2011;18(4):633–9.
Heimburg-Molinaro J, Priest JW, Live D, Boons GJ, Song X, Cummings RD, et al. Microarray analysis of the human antibody response to synthetic Cryptosporidium glycopeptides. Int J Parasitol. 2013;43(11):901–7.
Borad AJ, Allison GM, Wang D, Ahmed S, Karim MM, Kane AV, et al. Systemic antibody responses to the immunodominant p23 antigen and p23 polymorphisms in children with cryptosporidiosis in Bangladesh. Am J Trop Med Hyg. 2012;86(2):214–22.
Allison GM, Rogers KA, Borad A, Ahmed S, Karim MM, Kane AV, et al. Antibody responses to the immunodominant Cryptosporidium gp15 antigen and gp15 polymorphisms in a case–control study of cryptosporidiosis in children in Bangladesh. Am J Trop Med Hyg. 2011;85(1):97–104.
Wanyiri JW, Kanyi H, Maina S, Wang DE, Steen A, Ngugi P, et al. Cryptosporidiosis in HIV/AIDS patients in Kenya: clinical features, epidemiology, molecular characterization and antibody responses. Am J Trop Med Hyg. 2014;91(2):319–28. This study evaluates the protective effect of antibody responses in patients with cryptosporidiosis.
Lai O, Morris C, Ahmed S, Karim MM, Khan W, Ward H, et al. Serum antibody responses to polymorphic Cryptosporidium mucin antigen in Bangladeshi children with cryptosporidiosis. Am J Trop Med Hyg. 2011;85(3):464–70.
Mead JR. Challenges and prospects for a Cryptosporidium vaccine. Future Microbiol. 2010;5:335–7.
Widmer G, Lee Y, Hunt P, Martinelli A, Tolkoff M, Bodi K. Comparative genome analysis of two Cryptosporidium parvum isolates with different host range. Infect Genet Evol. 2012;12(6):1213–21.
Sheoran A, Wiffin A, Widmer G, Singh P, Tzipori S. Infection with cryptosporidium hominis provides incomplete protection of the host against cryptosporidium parvum. J Infect Dis. 2012;205(6):1019–23.
Steinhagen F, Kinjo T, Bode C, Klinman DM. TLR-based immune adjuvants. Vaccine. 2011;29(17):3341–55.
Manque PA, Woehlbier U, Lara AM, Tenjo F, Alves JM, Buck GA. Identification and characterization of a novel calcium-activated apyrase from cryptosporidium parasites and its potential role in pathogenesis. PLoS One. 2012;7(2):e31030.
Singh I, Theodos C, Tzipori S. Recombinant proteins of Cryptosporidium parvum induce proliferation of mesenteric lymph node cells in infected mice. Infect Immun. 2005;73(8):5245–8.
Benitez A, Priest JW, Ehigiator HN, McNair N, Mead JR. Evaluation of DNA encoding acidic ribosomal protein P2 of Cryptosporidium parvum as a potential vaccine candidate for cryptosporidiosis. Vaccine. 2011;29(49):9239–45.
Wang C, Luo J, Amer S, Guo Y, Hu Y, Lu Y, et al. Multivalent DNA vaccine induces protective immune responses and enhanced resistance against Cryptosporidium parvum infection. Vaccine. 2010;29(2):323–8.
Zheng J, Ren W, Pan Q, Wang Q, Elhag IA, Li J, et al. A recombinant DNA vaccine encoding C. andersoni oocyst wall protein induces immunity against experimental C. parvum infection. Vet Parasitol. 2011.
Yu Q, Li J, Zhang X, Gong P, Zhang G, Li S, et al. Induction of immune responses in mice by a DNA vaccine encoding Cryptosporidium parvum Cp12 and Cp21 and its effect against homologous oocyst challenge. Vet Parasitol. 2010;172(1–2):1–7.
Liu K, Zai D, Zhang D, Wei Q, Han G, Gao H, et al. Divalent Cp15-23 vaccine enhances immune responses and protection against Cryptosporidium parvum infection. Parasite Immunol. 2010;32(5):335–44. This study compares the protective effect of univalent and divalent Cryptosporidium DNA-based vaccines.
Khan KH. DNA vaccines: roles against diseases. Germs. 2013;3(1):26–35.
Benitez AJ, McNair N, Mead JR. Oral immunization with attenuated Salmonella enterica serovar Typhimurium encoding Cryptosporidium parvum Cp23 and Cp40 antigens induces a specific immune response in mice. Clin Vaccine Immunol. 2009;16(9):1272–8.
Roche JK, Rojo AL, Costa LB, Smeltz R, Manque P, Woehlbier U, et al. Intranasal vaccination in mice with an attenuated Salmonella enterica Serovar 908htr A expressing Cp15 of Cryptosporidium: impact of malnutrition with preservation of cytokine secretion. Vaccine. 2013;31(6):912–8. This study suggests that malnutrition negatively impacts mucosal immune responses to Cryptosporidium.
Galen JE, Zhao L, Chinchilla M, Wang JY, Pasetti MF, Green J, et al. Adaptation of the endogenous Salmonella enterica serovar Typhi clyA-encoded hemolysin for antigen export enhances the immunogenicity of anthrax protective antigen domain 4 expressed by the attenuated live-vector vaccine strain CVD 908-htrA. Infect Immun. 2004;72(12):7096–106.
Manque PA, Tenjo F, Woehlbier U, Lara AM, Serrano MG, Xu P, et al. Identification and immunological characterization of three potential vaccinogens against Cryptosporidium species. Clin Vaccine Immunol. 2011;18(11):1796–802. This study uses “reverse vaccinology” to identify potential Cryptosporidium vaccine candidates.
Xu RG, Jia H, Terkawi MA, Xuan X, Zhang H. Immunogenicity of orally administrated recombinant lactobacillus casei Zhang expressing Cryptosporidium parvum surface adhesion protein P23 in mice. Curr Microbiol. 2011;62(5):1573–80.
Kim K, Weiss LM. Toxoplasma gondii: the model apicomplexan. Int J Parasitol. 2004;34(3):423–32.
O’Connor RM, Wanyiri JW, Wojczyk BS, Kim K, Ward H. Stable expression of Cryptosporidium parvum glycoprotein gp40/15 in Toxoplasma gondii. Mol Biochem Parasitol. 2007;152(2):149–58.
Shirafuji H, Xuan X, Kimata I, Takashima Y, Fukumoto S, Otsuka H, et al. Expression of P23 of Cryptosporidium parvum in Toxoplasma gondii and evaluation of its protective effects. J Parasitol. 2005;91(2):476–9.
Yao L, Yin J, Zhang X, Liu Q, Li J, Chen L, et al. Cryptosporidium parvum: identification of a new surface adhesion protein on sporozoite and oocyst by screening of a phage-display cDNA library. Exp Parasitol. 2007;115(4):333–8.
Priest JW, Kwon JP, Montgomery JM, Bern C, Moss DM, Freeman AR, et al. Cloning and characterization of the acidic ribosomal protein P2 of Cryptosporidium parvum, a new 17-kilodalton antigen. Clin Vaccine Immunol. 2010;17(6):954–65.
Xiao D, Yin C, Zhang Q, Li JH, Gong PT, Li SH, et al. Selection and identification of a new adhesion protein of Cryptosporidium parvum from a cDNA library by ribosome display. Exp Parasitol. 2011.
Compliance with Ethics Guidelines
Conflict of Interest
Jacob G. Ludington and Honorine D. Ward declare that they have no conflict of interest.
Human and Animal Rights and Informed Consent
This is a review article, in which no studies with human or animal subjects have been performed by either of the authors.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is part of the Topical Collection on Protozoa
Rights and permissions
About this article

Cite this article
Ludington, J.G., Ward, H.D. Systemic and Mucosal Immune Responses to Cryptosporidium—Vaccine Development. Curr Trop Med Rep 2, 171–180 (2015). https://doi.org/10.1007/s40475-015-0054-y
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40475-015-0054-y