
Abstract
An epidemiologic approach for the understanding of the public health relevance of Strongyloides stercoralis is currently much needed, due to the renewed interest in the control of soil-transmitted helminthiasis (STH). Despite this fact, the biologic difference of S. stercoralis compared to the other STH in terms of life cycle, diagnostic approach and therapeutic options leaves this nematode under uncertainties that hamper the development of control strategies. However, evidence regarding distribution and prevalence allows for the identification of its global distribution, with lack of adequate water and sanitation supplies as the most significant risk factors. The clinical aspects of this infection in immunocompromised populations also highlight its potentially severe morbidity in groups that are at higher risk for complications during infection. The availability of more accurate diagnostic tools appears as a key element for the performance of more reliable surveys, case control studies and even monitoring for the emergence of drug resistance.
Similar content being viewed by others

Introduction
Understanding epidemiology as one of the main sources of analytic information that guides rational public health decisions, and that “evidence-based public health” is supported by a solid knowledge base of disease frequency and distribution of the determinants and consequences of disease, and of the safety, efficacy and effectiveness of interventions [1], it is of fundamental practical importance to identify the epidemiologic aspects of the different diseases in order to design interventions with a measurable public health impact. The case of neglected tropical diseases (NTDs) and the launching of new coalitions with funds and drug donations that allow the framing of ambitious control, and in some cases, eliminations, is an example of the need for clear and up-to-date information of affected people and all the determinants that need to be considered in the design of these strategies [2, 3]. As programs start their activities, data on the epidemiologic aspects of NTDs are required to target treatment to areas and communities at greatest need and to estimate drug and resource requirements. In addition, as interventions are scaled up, governments and donors require clear information on the impact of these control efforts.
Strongyloides stercoralis is probably among the most significant examples of the need to set a clear agenda of knowledge acquisition in order to brake the vicious circle where the lack of adequate diagnostic and therapeutic tools causes its exclusion from the goals set for the other soil transmitted helminthes (STH). This results in its omission from the list of priorities, despite the lack of knowledge on its clinical, sanitary and epidemiologic implications [4, 5].
The Parasite and the Infection
The parasitic nematodes belonging to the Strongyloides group are members of the Rhabditida family of free-living nematodes. Among the different Strongyloides species that have been identified, S. stercoralis stands out as the species with the widest distribution and most significant importance as a human pathogen. Originally described in 1876 among French troops in current Vietnam by Louis Normand while evaluating stools from soldiers with diarrhea, it was later that same year that in an autopsy performed by Normand, the adult forms were first described; though he did not realize that were different stages of the same parasite [6, 7]. Strongyloides fullerborni, restricted to some parts of Africa and Papua New Guinea, is a parasite of Old World primates that can also infect humans [7, 8].
The distinctively complex life cycle of S. stercoralis, compared to other STH of medical importance, makes this species a particularly interesting subject for study and investigation, with the importance placed in the proper identification of features that are shared with the other STH and those features that are both different and of clinical and epidemiologic significance in terms of disease control [5, 9–11]. This alternative switch between parasitic and free-living modes of existence and the rapid evolution into infective forms that allows for reinfection are the most significant, but not the only features that define the biology, the clinical aspects and also the epidemiology of S. stercoralis [12].
Strongyloidiasis, the disease caused by infections with S. stercoralis, identifies more of an etiologic origin than a particular syndrome, since the infection with this helminth could range from asymptomatic states to diarrheal disease and the hyperinfection state in the extremes of aggressiveness that makes of this infection a life-threatening condition in immune deficient individuals [13–15].
Control strategies against STH and other NTDs, which for the most part have ignored this parasite, require an adequate body of evidence on the epidemiology of this infection and its different clinical presentations; this includes, as in any public health issue, evidence-based and updated data. This package is currently missing and therefore hampering the implementation of control efforts [5].
Transmission
The most salient aspect of a review of the literature with regard to the epidemiologic characteristics of S. stercoralis, unfortunately, is the scarcity of information regarding the distribution, prevalence, and burden of disease. The roots of this deficiency are, for the most part, the same that are shared by all the diseases that despite their biologic and clinical differences belong to the group of NTDs; these are the historic lack of investments and efforts for control by the governments and industry, and the uniform link to poverty and inequality suffered by those affected by these diseases [16].
The life cycle of S. stercoralis defines its distribution, as explained above, through the elimination of larvae and not eggs. It also defines a critical transmission characteristic, which is the auto-infective stage, where the filariform larval stage is reached before leaving the human environment, whether intraluminal in the gut or cutaneous in the perianal region, allowing autoinfection and the perpetuation of infection to its chronic stages for periods of as long as 75 years [17]. Currently, with a renewed interest on the use of transmission dynamics modeling as a source of information for STH control strategies, the classification between microparasites and macroparasites in the classic works by Anderson & May for the study of population dynamics [18] requires a particularly cautious approach in the case of S. stercoralis, which unlike the other STH and despite having the characteristics of a macroparasite for the most part, also features a distinctive ability for direct multiplication at large scale within the host. This characteristic also has therapeutic implications in terms of the requirement for curative treatments rather than the much-accepted regimens against STH, which aim for lowering the worm burden.
The approach to the identification of the determinants for the transmission of S. stercoralis to the human host can also be inferred from studies carried out on human volunteers, before the potential lethality of this parasite was known, which showed the ability of the filariform larvae to penetrate the human skin [19•]. As per the risk of infection through drinking water, the inability of the larvae to swim, an issue that can be verified in the Baermann diagnostic method, leaves the larvae to settle in the bottom of the water source. This issue suggests that WASH (water, sanitation and hygiene) efforts should impact the prevalence and distribution of Strongyloides through the sanitation component to a higher degree than the water component. Sexual transmission and inter-human transmission through close contact, despite being feasible, do not pose significant transmission routes [7, 20].
S. stercoralis infections are unusual in countries where fecal contamination of soil is rare and the exposure to third-stage larvae by the general population is not present due to sanitation improvements. A recent study in a cohort of children aged 8–18 years old from rural villages of Cambodia showed that children who reported having shoes and defecating in toilets were less likely to be infected with S. stercoralis than those who did not possess shoes and used unimproved latrines [21•].
Diagnosis
S. stercoralis has some particularities that make it harder to diagnose, compared to other STH; central to this situation are the diagnostic difficulties posed by this parasite, which is systematically missed by the diagnostic methods recommended by the World Health Organization for the diagnosis of STH, like Kato Katz and secondarily McMaster, since these methods are based on the detection of eggs, a stage that is very rarely, if ever, found aside from in hyperinfective states of infection [5]. This issue, added to the requirement of multiple serial samples in order to properly call an individual non-infected, renders most surveys useless to estimate prevalence and distribution, unless alternative methods specific for the detection of larvae in stools are performed. These limitations in diagnosis cancel the possibilities of determining the risk factors and infection determinants through case control studies, since it is not possible to rule out infections in individuals from endemic areas with stool analyses that are negative for S. stercoralis.
In contrast to other STH such as Necator americanus, Ancylostoma duodenale, Trichuris trichura and Ascaris lumbricoides, the diagnosis of strongyloidiasis is primarily based on the detection of the filariform larvae (L3) by microscopic examinations of stool samples. Nowadays, the standard parasitological diagnostic methods for detecting S. stercoralis infections are Baermann and Agar plate culture methods, or, if possible, a combination of these methods [22, 23]. These are techniques with higher sensitivity, but are not considered as “Gold Standard” for the diagnosis of the strongyloidiasis. Direct fecal smear examination, formol-ether concentration and Harada-Mori are other parasitologic methods that are available, all with sub-optimal sensitivity, which is lowered further when performed on a single specimen per patient due to the fluctuating nature of larva elimination in stool [24–26].
Several immunological tests, such as ELISA, western blot, immunofluorescence and luciferase immunoprecipitation system (LIPS), have also been described for the diagnosis of strongyloidiasis [27–29]. These assays show variable sensitivity and specificity, depending on the antigen used (crude extract or recombinant antigens) and population tested [27, 30]. Currently, there is a need to improve the tools available for the diagnosis of this parasite, and for this, it will be very important to incorporate new technologies (i.e., LIPS or multiplex assays), as well as the search for new recombinant antigens (i.e., phage display technology) [29, 31, 32]. In order to be useful for the understanding of the epidemiology of strongyloidiasis and its clinical circumstances, there is a need for a better understanding of the accuracy and dynamics of antibody decay in response to curative antiparasitary regimens [27]. Polymerase chain reaction (PCR) has been developed and used for the detection of DNA from Strongyloides and other helminths in fecal samples. Recently, the evaluation of conventional PCR and real-time PCR has proved to be a sensitive and specific method for the diagnosis of S. stercoralis [33–36].
Epidemiological Determinants
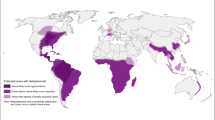
Strongyloidiasis caused by S. stercoralis has been estimated to affect 30–100 million people worldwide [4, 22]. These numbers are, however, just estimates that need better evidence and updates. This threadworm intestinal parasite that infects dogs, cats and primates including humans is endemic in tropical and subtropical regions with poor sanitation conditions and where climatic conditions are warm and humid, such as Central and South America, sub-Saharan Africa, and South and Southeast Asia [37•].
In a recent study aimed at the description of the global distribution of strongyloidiasis through a literature search of surveys and reports, including hospital records, on all types of populations from around the world, the conclusions suggest that S. stercoralis infections affect between 10 % and 40 % of the population in many tropical and subtropical countries. This information is taken mainly from community-based studies. Brazil and Thailand provided the greatest amount of epidemiological information from community-based studies, while Australia and United States were the main sources of information from hospital-based reports and immigrant and refugees surveys [37•]. The application of methods with poor sensitivity is a problem that leads to underestimation of the prevalence; of 194 community surveys, only 9 % included techniques with relatively high sensitivity to detect S. stercoralis [23, 37•]. In Table 1, we summarize the prevalence in different endemic countries, understanding that these studies do not represent the overall prevalence in the whole country, but rather in communities where the socio-economic conditions support the persistence and proliferation of this parasite. In each of these studies, the diagnostic method used was Baermann and/or Agar plate.
The reports with the highest prevalence are from Argentina and Cambodia (>40 %). In Argentina, data about the prevalence of S. stercoralis is scarce and limited to few areas, mainly in the northwest, but significant for the presence of surveys with prevalences of over 80 % in school children and community-based surveys including children and adults [30, 38]. In northern Argentina, particularly in the region of the Yungas rainforest and the Chaco region, different reports indicate prevalence rates of over 20 % [39]. In a recent study in Salta (unpublished data), examination of stool samples including several parasitological methods (concentration-sedimentation, Agar plate, Harada-Mori and Baermann) showed that 16 % of the populations were infected, while an enzyme-linked immunosorbent assay for detecting S. stercoralis antibody (NIE-ELISA) revealed a prevalence of 31 % in the same population. These surveys, performed mainly by academic centers with activities in that region, probably represent a marker of similar levels of infection prevalence across a wider territory, including the whole Chaco region where ecological and socio-economic conditions allow the transmission of S. stercoralis and other STH.
Developed countries like Japan, Australia, Western Europe and United States currently have prevalence levels of S. stercoralis that are low and mostly restricted to immigrants from endemic countries [37•, 40]. In Japan, it is only endemic in the Okinawa island area, where it has for a long time been identified at high levels in association with HTLV-1 infections; there, the prevalence in community surveys was 18.7 % and 13.6 % in hospital investigations. The cases reported are mostly among the older population [37•]. In Australia, data about the prevalence of S. stecoralis infection derives from studies on refugees and immigrants, with prevalence estimates of 25 %, and also from aboriginal communities in the northern territories [37•, 41, 42]. In the United States, the prevalence estimate of the infection rates among refugees and immigrants, mainly from Southeast Asia, was found to be up to 40 % [37•]; in Canada, a similar finding was obtained in a seroepidemiology study of refugees from Southeast Asia, with Cambodians being distinctively affected [43].
The current strategy of the World Health Organization (WHO) for the control of the morbidity caused by STH has preventive chemotherapy as the mainstay [44]; however, neither of the drugs used for this effort, albendazole and mebendazole, has significant activity against S. stercoralis at single doses. Adding ivermectin would be the complement to the benzimidazol drugs if activity against S. stercoralis for a wider anti-STH regimen was to be attained. The mass distribution of ivermectin had its benefits, but re-infection is possible; in a 2-year follow-up in children treated for S. stercoralis infection, 31 % were re-infected, while 20 % of the healthy children were infected during follow-up [21•].
Risk Groups
The epidemiology of hyperinfection syndromes in strongyloidiasis, which occur due to the amplification of the auto-infective path, is mostly secondary to the presence of immunosuppressive conditions due to drugs or health conditions, with the added effect of corticosteroids in the enhanced fecundity of the parthenogenetic adult females of S. stercoralis in the gastrointestinal tract [14]. These groups of health conditions with impaired immune responses include lymphomas, alcoholism, HTLV-1 infections and corticosteroids as the most widely used among a longer list of immunosuppressive drugs. It should be noted, however, that these conditions have not been linked to an increased risk of infection, but rather to the development of severe clinical forms of the disease [37•]. HIV infection, despite initial impressions, does not predispose to hyperinfection, and strongyloidiasis is therefore not in the list of opportunistic infections related to this retroviral infection [37•]. Among the impoverished populations of the world, where the sanitary conditions favor the exposure to all STH, malnutrition among children is the most common condition affecting adequate immune responses, and therefore is a risk factor most frequently seen related to severe strongyloidiasis. Malnutrition is also probably related to the occurrence of these clinical forms among alcoholics, since despite studies demonstrating a higher prevalence of S. stercoralis in alcoholics than controls, animal studies have shown that chronic alcohol consumption does not affect specific immune responses against S. stercoralis in mice kept with diets that were adequate regarding caloric and micronutrient components [45, 46].
Strongyloides fuelleborni
Among the other species of Strongyloides, S. fuelleborni is the only other potentially significant human pathogen; this species infects principally non-human primates. The infection has been found in humans in Africa and Papua New Guinea [4]. In a recent study in wild Japanese macaques (Macaca fuscata), S. fuelleborni was detected among other helminths (Trichuris trichiura and Oesophagostoma maculeatum) [47]. In mountain gorillas (Gorilla beringei) from Bwindi Impenetrable National Park (BINP), Uganda, the presence of S. fuelleborni was also detected [48]. This parasitic infection is zoonotic and rare in humans, with the first cases of infection with S. fuelleborni reported in Zambia [8]. In regions where there is frequent contact between humans and non-human primates, this zoonotic nematode poses a potential health issue to local people.
Antihelmintic Resistance in S. stercoralis
The emergence of antihelmintic drug resistance is a major problem for animal and human health. To date, there is no evidence of anthelmintic resistance in humans among STH, but this absence of evidence should be interpreted with caution as large-scale deworming programs are deployed, coverage is increased and a larger group of people receives repeated cycles of drugs at not uniformly curative regimens [49•, 50]. The case for S. stercoralis falls within this situation, and there is no information on resistance that might alter response to therapy and the epidemiologic aspects related to the response to intervention programs.
Current guidelines for the control of STH recommend mass drug administration (MDA) once a year when STH prevalence is ≥20 % but less than 50 %, and twice a year when the prevalence is >50 % [49•, 51]. The benzimidazole drugs, albendazole and mebendazole, are commonly used for MDA, while ivermectin is recommended for strongyloidiasis, lymphatic filariasis and onchocerciasis [5, 52]; however, its use in MDA programs is currently limited to onchocerciasis, with other indications not covered by drug donations therefore used only for individual case management [53]. The MDA programs reduce the incidence and intensity of infections; however, this strategy could trigger the development of anthelmintic resistance [49•].
Many studies have demonstrated that the frequent use of anthelmintic drugs (up to eight times per year) in veterinary nematodes has led to the development of resistance. In humans, the frequency of treatment is low (once or twice per year) and the coverage is far from the 100 % achieved in veterinary nematodes; therefore, the selection pressure is less than for parasites in animals and development of anthelmintic resistance in STH will be slow and less likely to occur [49•].
Conclusion
The epidemiology of S. stercoralis has distinctive features that require careful consideration for the management of affected communities and individuals. The lack of accurate diagnostic and monitoring tools is a significant obstacle for a more complete understanding of its dynamics, risk factors and response to interventions. These issues should trigger renewed efforts and creativity in epidemiologic research in order to find ways to tackle this NTD.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance
Victora CG, Habicht J-P, Bryce J. Evidence-based public health: moving beyond randomized trials. Am J Public Health. 2004;94:400–5.
Zhou X-N, Bergquist R, Tanner M. Elimination of tropical disease through surveillance and response. Infect Dis Poverty. 2013;2:1.
Webster JP, Molyneux DH, Hotez PJ, Fenwick A. The contribution of mass drug administration to global health: past, present and future. Philos Trans R Soc Lond B Biol Sci. 2014;369(1645):20130434. doi:10.1098/rstb.2013.0434.
Olsen A, van Lieshout L, Marti H, Polderman T, Polman K, Steinmann P, et al. Strongyloidiasis—the most neglected of the neglected tropical diseases? Trans R Soc Trop Med Hyg. 2009;103:967–72.
Krolewiecki AJ, Lammie P, Jacobson J, Gabrielli A-F, Levecke B, Socias E, et al. A public health response against Strongyloides stercoralis: time to look at soil-transmitted helminthiasis in full. PLoS Negl Trop Dis. 2013;7(5):e2165. doi:10.1371/journal.pntd.0002165.
Cox FEG. History of human parasitology. Clin Microbiol Rev. 2002;15:595–612.
Grove DI. Strongyloidiasis: a major roundworm infection of man. Taylor & Francis. 1989. pp. 336.
Hira P, Patel B. Strongyloides fulleborni infections in man Zambia. Am J Trop Med Hyg. 1977;26:640–3.
Streit A. Reproduction in Strongyloides (Nematoda): a life between sex and parthenogenesis. Parasitology. 2008;135:285–94.
Viney M. The biology and genomics of Strongyloides. Med Microbiol Immunol. 2006;195:49–54.
Gabrielli A-F, Montresor A, Chitsulo L, Engels D, Savioli L. Preventive chemotherapy in human helminthiasis: theoretical and operational aspects. Trans R Soc Trop Med Hyg. 2011;105:683–93.
Lok JB. Strongyloides stercoralis: a model for translational research on parasitic nematode biology. WormBook. 2007;1–18.
Mejia R, Nutman TB. Screening, prevention, and treatment for hyperinfection syndrome and disseminated infections caused by Strongyloides stercoralis. Curr Opin Infect Dis. 2012;25:458–63.
Keiser PB, Nutman TB. Strongyloides stercoralis in the Immunocompromised population. Clin Microbiol Rev. 2004;17:208–17.
Becker SL, Sieto B, Silué KD, Adjossan L, Koné S, Hatz C, et al. Diagnosis, clinical features, and self-reported morbidity of Strongyloides stercoralis and hookworm infection in a co-endemic setting. PLoS Negl Trop Dis. 2011;5(8):e1292. doi:10.1371/journal.pntd.0001292.
Conteh L, Engels T, Molyneux DH. Socioeconomic aspects of neglected tropical diseases. Lancet. 2010;375:239–47.
Prendki V, Fenaux P, Durand R, Thellier M, Bouchaud O. Strongyloidiasis in man 75 years after initial exposure [letter]. Emerg Infect Dis. 2011;17:931–2.
Anderson RM, May RM. Population biology of infectious diseases: part I. Nature. 1979;280:361–7.
Freedman DO. Experimental infection of human subject with Strongyloides species. Rev Infect Dis. 1991;13:1221–6. A very comprehensive recount of historic data with significant experimental pieces for the understanding of the clinical and transmission aspects of the infection.
Grove DI. Strongyloidiasis: is it transmitted from husband to wife? Br J Vener Dis. 1982;58:271–2.
Khieu V, Hattendorf J, Schär F, Marti H, Char MC, Muth S, et al. Strongyloides stercoralis infection and re-infection in a cohort of children in Cambodia. Parasitol Int. 2014;63(5):708–12. doi:10.1016/j.parint.2014.06.001. A major study of the infection and re-infection rates, important to understand the transmission dynamics of Strongyloides stercoralis in endemic countries.
Siddiqui AA, Berk SL. Diagnosis of Strongyloides stercoralis infection. Clin Infect Dis. 2001;33:1040–7.
Requena-Méndez A, Chiodini P, Bisoffi Z, Buonfrate D, Gotuzzo E, Muñoz J. The laboratory diagnosis and follow up of strongyloidiasis: a systematic review. Bottazzi ME, editor. PLoS Negl Trop Dis. 2013;7(1):e2002. doi: 10.1371/journal.pntd.0002002.
Uparanukraw P, Phongsri S, Morakote N. Fluctuations of larval excretion in Strongyloides stercoralis infection. Am J Trop Med Hyg. 1999;60:967–73.
Knopp S, Mgeni AF, Khamis IS, Steinmann P, Stothard JR, Rollinson D, et al. Diagnosis of soil-transmitted helminths in the era of preventive chemotherapy: effect of multiple stool sampling and use of different diagnostic techniques. PLoS Negl Trop Dis. 2008;2(11):e331. doi:10.1371/journal.pntd.0000331.
Dreyer G, Fernandes-silva E, Alves S, Rocha A, Albuquerque R, Addiss D. Patterns of detection of Strongyloides stercoralis in stool Specimens : implications for diagnosis and clinical trials. J Clin Microbiol. 1996;34:2569–71.
Bisoffi Z, Buonfrate D, Sequi M, Mejia R, Cimino RO, Krolewiecki AJ, et al. Diagnostic accuracy of five serologic tests for Strongyloides stercoralis infection. PLoS Negl Trop Dis. 2014;8(1):e2640. doi:10.1371/journal.pntd.0000331.
Gonzaga HT, Ribeiro VS, Feliciano ND, Manhani MN, Silva DAO, Ueta MT, et al. IgG avidity in differential serodiagnosis of human strongyloidiasis active infection. Immunol Lett. 2011;139:87–92.
Ramanathan R, Burbelo PD, Groot S, Iadarola MJ, Neva FA, Nutman TB. A luciferase immunoprecipitation systems assay enhances the sensitivity and specificity of diagnosis of Strongyloides stercoralis infection. J Infect Dis. 2008;198:444–51.
Krolewiecki AJ, Ramanathan R, Fink V, McAuliffe I, Cajal SP, Won K, et al. Improved diagnosis of Strongyloides stercoralis using recombinant antigen-based serologies in a community-wide study in northern Argentina. Clin Vaccine Immunol. 2010;17:1624–30.
Feliciano ND, da Silva Ribeiro V, de Almeida Araújo Santos F , Fujimura PT , Gonzaga HT, et al. Bacteriophage-fused peptides for serodiagnosis of human strongyloidiasis. PLoS Negl Trop Dis. 2014;8(5):e2792.doi: 10.1371/journal.pntd.0002792
Moss DM, Priest JW, Boyd A, Weinkopff T, Kucerova Z, Beach MJ, et al. Multiplex bead assay for serum samples from children in Haiti enrolled in a drug study for the treatment of lymphatic filariasis. Am J Trop Med Hyg. 2011;85:229–37.
Mejia R, Vicuña Y, Broncano N, Sandoval C, Vaca M, Chico M, et al. A novel, multi-parallel, real-time polymerase chain reaction approach for eight gastrointestinal parasites provides improved diagnostic capabilities to resource-limited at-risk populations. Am J Trop Med Hyg. 2013;88:1041–7.
Repetto SA, Alba Soto CD, Cazorla SI, Tayeldin ML, Cuello S, Lasala MB, et al. An improved DNA isolation technique for PCR detection of Strongyloides stercoralis in stool samples. Acta Trop. 2013;126:110–4.
Sitta RB, Malta FM, Pinho JR, Chieffi PP, Gryschek RCB, Paula FM. Conventional PCR for molecular diagnosis of human strongyloidiasis. Parasitology. 2014;141(5):716–21.
Verweij JJ, Canales M, Polman K, Ziem J, Brienen EAT, Polderman AM, et al. Molecular diagnosis of Strongyloides stercoralis in faecal samples using real-time PCR. Trans R Soc Trop Med Hyg. 2009;103:342–6.
Schär F, Trostdorf U, Giardina F, Khieu V, Muth S, Marti H, et al. Strongyloides stercoralis: global distribution and risk factors. PLoS Negl Trop Dis. 2013;7(7):e2288. doi:10.1371/journal.pntd.0002288. An important systematic review of the global prevalence of Strongyloides stercoralis and risk factors.
Taranto NJ, Bonomi de Filippi H, Orione O. Prevalence of Strongyloides stercoralis infection in childhood. Orán, Salta, Argentina. Bol Chil Parasitol. 1993;48:49–51.
Socías ME, Fernández A, Gil JF, Krolewiecki AJ. Geohelmintiasis en la Argentina una revisión sistemática. Medicina (Buenos Aires). 2014;74:29–36.
Genta RM. Global prevalence of strongyloidiasis: critical review with epidemiologic insights into the prevention of disseminated disease. Rev Infec Dis. 1989;11:755–67.
Prociv P, Luke R. Observations on strongyloidiasis in Queensland aboriginal communities. Med J Aust. 1993;158:160–3.
Fisher D, McCarry F, Currie B. Strongyloidiasis in the Northern Territory. Under-recognised and under-treated? Med J Aust. 1993;159:88–90.
Gyorkos TW, Genta RM, Viens P, MacLean JD. Seroepidemiology of Strongyloides infection in the Southeast Asian refugee population in Canada. Am J Epidemiol. 1990;132:257–64.
Albonico M, Montresor A, Crompton DWT, Savioli L. Intervention for the control of soil-transmitted helminthiasis in the community. Adv Parasitol. 2006;61:311–48.
Zago-Gomes MP, Aikawa KF, Perazzio SF, Gonçalves CS, Pereira FEL. Prevalence of intestinal nematodes in alcoholic patients. Rev Soc Bras Med Trop. 2002;35:571–4.
Krolewiecki AJ, Leon S, Scott PA, Nolan TJ, Schad GA, Abraham D. Effect of chronic ethanol consumption on protective T-helper 1 and T-helper 2 immune responses against the parasites Leishmania major and Strongyloides stercoralis in mice. Alcohol Clin Exp Res. 2001;25:571–8.
Arizono N, Yamada M, Tegoshi T, Onishi K. Molecular identification of Oesophagostomum and Trichuris eggs isolated from wild Japanese macaques. Korean J Parasitol. 2012;50:253–7.
Rothman JM, Pell AN, Bowman DD. Host-parasite ecology of the helminths in mountain gorillas. J Parasitol. 2008;94:834–40.
Vercruysse J, Levecke B, Prichard R. Human soil-transmitted helminths: implications of mass drug administration. Curr Opin Infect Dis. 2012;25:703–8. Important review on mass drug administration and the implications of the development of anthelmintic resistance in humans.
Diawara A, Drake LJ, Suswillo RR, Kihara J, Bundy DAP, Scott ME, et al. Assays to detect beta-tubulin codon 200 polymorphism in Trichuris trichiura and Ascaris lumbricoides. PLoS Negl Trop Dis. 2009;3(3):e397. doi:10.1371/journal.pntd.0000397.
World Health Organization. Helminth control in school-age children: a guide for managers of control programmes. 2da ed. Geneva; 2011. p. 18.
World Health Organization. Weekly epidemiological record: relevé épidémiologique hebdomadaire. 2013 p. 145–52.
Bush S, Hopkins AD. Public-private partnerships in neglected tropical disease control: the role of nongovernmental organisations. Acta Trop. 2011;120(Suppl):S169–72.
Taranto N, Cajal S, De Marzi M, Fernández M, Frank F, Brú A, et al. Clinical status and parasitic infection in a Wichí Aboriginal community in Salta, Argentina. Trans R Soc Trop Med Hyg. 2003;97:554–8.
Khieu V, Schär F, Marti H, Sayasone S, Duong S, Muth S, et al. Diagnosis, treatment and risk factors of Strongyloides stercoralis in school children in Cambodia. PLoS Negl Trop Dis. 2013;7(2):e2035. doi:10.1371/journal.pntd.0002035.
Schär F, Inpankaew T, Traub RJ, Khieu V, Dalsgaard A, Chimnoi W, et al. The prevalence and diversity of intestinal parasitic infections in humans and domestic animals in a rural Cambodian village. Parasitol Int. 2014;63:597–603.
Khieu V, Schär F, Marti H, Bless PJ, Char MC, Muth S, et al. Prevalence and risk factors of Strongyloides stercoralis in Takeo Province, Cambodia. Parasite Vectors. 2014;7:221.
Khieu V, Schär F, Forrer A, Hattendorf J, Marti H, Duong S, et al. High prevalence and spatial distribution of Strongyloides stercoralis in Rural Cambodia. PLoS Negl Trop Dis. 2014;8(6):e2854. doi:10.1371/journal.pntd.0002854.
Steinmann P, Zhou X-N, Du Z-W, Jiang J-Y, Wang L-B, Wang X-Z, et al. Occurrence of Strongyloides stercoralis in Yunnan Province, China, and comparison of diagnostic methods. PLoS Negl Trop Dis. 2007;1(1):e75. doi:10.1371/journal.pntd.0000075.
Machicado JD, Marcos LA, Tello R, Canales M, Terashima A, Gotuzzo E. Diagnosis of soil-transmitted helminthiasis in an Amazonic community of Peru using multiple diagnostic techniques. Trans R Soc Trop Med Hyg. 2012;106:333–9.
Yori PP, Kosek M, Gilman RH, Cordova J, Bern C, Chavez CB, et al. Seroepidemiology of strongyloidiasis in the Peruvian Amazon. Am J Trop Med Hyg. 2006;74:97–102.
Compliance with Ethics Guidelines
Conflict of Interest
Rubén O. Cimino and Alejandro Krolewiecki declare that they have no conflicts of interest.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Cimino, R.O., Krolewiecki, A. The Epidemiology of Human Strongyloidiasis. Curr Trop Med Rep 1, 216–222 (2014). https://doi.org/10.1007/s40475-014-0035-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40475-014-0035-6