
Abstract
How the ciliates of fish can be cultured and be used to study ciliate/fish interactions are reviewed. The culturing of ciliates is currently done in basal solutions based on either freshwater, seawater, or bodily fluids of vertebrates. These are supplemented either with bacteria, fish cells, or organic matter, which can be defined or more commonly undefined, with proteose peptone being a prominent example. Among pathogenic ciliates, the most difficult to culture has been Ichthyophthirius multifiliis. To contrast, Cryptocaryon irritans caused a similar disease and has been maintained successfully in co-cultures with fish cells. Pathogenic scuticociliates and tetrahymenas are more amenable to culture, and can be grown axenically. These cultures have been used to study pathogenic mechanisms and to screen drugs for their potential chemotherapeutic value. Ciliates may act directly on fish to cause disease, but may also influence fish health indirectly through their interactions with other types of fish pathogens, for example bacteria and fungi. For example, in culture Tetrahymena spp. have been shown to phagocytose pathogenic bacteria, Yersinia ruckerii, and microsporidia, Glugea hertwigi, where ciliates are not linked to disease, very different ciliate/fish interactions are possible. For some fish larvae, free-living ciliates are a source of nutrients. Large-scale cultures of both freshwater and marine ciliates have been achieved and could be a source of feed for fish larvae in aquaculture. Finally, ciliates have the potential to feed on fish carcasses and in doing so make nutrients available to the ecosystem. In the future cell cultures should be invaluable in studying these and other possible relationships between fish and ciliates.
Similar content being viewed by others

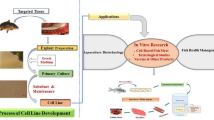
Introduction
Ciliates are single-celled organisms of the kingdom Protista. They are placed in the protozoan subdivision and belong to the Phylum Ciliophora [1]. Most ciliates are heterotrophs and can be broadly considered as either free-living or symbiotic. The boundary between free-living and symbiotic forms can be hard to draw [2], especially so, if consideration is given to the ciliates that retain functional prey chloroplasts or contain photosynthetic symbionts [3]. Here, the definitions will hinge simply on the ciliate’s relationship to metazoans. Free-living ciliates survive independently of metazoans. Symbiotic ciliates live closely with a metazoan either in mutual, commensal, or parasitic relationships. As heterotrophs, ciliates depend on an external source of organic compounds.
Free-living ciliates and symbiotic ciliates obtain organic compounds by similar processes, phagotrophy, and osmotrophy. Most ciliates have “mouths” and acquire nutrients by engulfing solid organic matter. This is phagotrophy and has been referred to as holozoic nutrition [4]. Free-living ciliates usually get nutrients by eating microbes, such as bacteria, algae, fungi, or other protozoa. Symbiotic ciliates eat microbes associated with metazoans, such as the rumen bacteria of cattle [5], or organic matter arising from the metazoans themselves. The organic matter can be solid, such as dead tissue or living cells. The eating of living metazoan cells is referred to as histophagy or “tissue eating” [6]. Some ciliates can also utilize organic compounds in solution. This is osmotrophy and has been referred to as saprozoic nutrition [4]. Osmotrophy is found among both symbiotic and free-living ciliates [4, 7]. The free-living ciliate, Tetrahymena pyriformis Ehrenberg, 1830 can simultaneously eat bacteria and consume dissolved organic matter [8]. The relative importance of phagotropy and osmotrophy for a particular species is largely unknown. A few mutants of T. thermophila Nanney and McCoy, 1976 appear capable of relying only on osmotropy [9].
Many ciliates are found in water and ciliate/fish interactions have long been of interest [7, 10–12] but increasingly they are becoming economically important as well [13–15, 16, 17]. Usually the focus has been on symbiotic relationships but here a more conservative approach is taken. For the purpose of this document, any of four types of interactions with fish will define a ciliate as a fish ciliate, and these interactions can be seen as a continuum from complex and dependent to simple and independent. First, the ciliates that are pathogens of fish will be considered fish ciliates. These ciliates could be in opportunistic, mutualistic, commensal, or parasitic relationships. A second group would be ciliates that feed on detritus from living, dying, and dead fish. They could be ectocommensals or free-living. A third group would be ciliates that either inactivate or protect viral and microbial pathogens of fish. These could be free-living or symbiotic ciliates. A fourth group would be ciliates that can serve as food for the early life stages of fish. These would be free-living ciliates.
One way to study some of these interactions is through the use of cell cultures. Cell cultures can be defined as the maintenance of cells in engineered structures. They allow engineers and scientists the ability to control either an industrial process or a research experiment. For industry, the yield can be either the cells or cell products; for science, the output is information. The histories for culturing single-cell eukaryotic organisms and cells of multicellular animals are long, but they have only infrequently crossed paths. Yet ciliates and fish do cross paths in numerous ways. The goals of this document are to provide a quick overview of ciliate/fish interactions, to review efforts to culture ciliates alone or with fish cells, and to comprehensively review the use of ciliate cultures in fish biology.
Ciliates of Fish
The four ciliate/fish interactions defined in the Introduction are reviewed briefly in the following sections. All the ciliates mentioned in the review are listed in Table 1 along with a single word or phrase describing their possible relationship with fish.
Ciliates as Fish Pathogens
As pathogens of fish, ciliates show a continuum of host dependencies that make interactions difficult to define formally in certain cases. Some ciliates are completely dependent on fish to complete their life cycle and act internally so are defined as obligate parasites or obligate endoparasites. This is the case for Ichthyophthirius multifiliis Fouquet, 1876, which causes “ich,” and Cryptocaryon irritans Brown, 1951, which causes “marine ich.” A second group acts internally but opportunistically and so are termed facultative parasites or facultative endoparasites. Belonging in this category are some members of the order Scuticociliatida Small, 1967 that cause the disease scuticociliatosis, and some species of Tetrahymena Furgason, 1940 that cause the disease tetrahymenosis. A third group is found on the outer surfaces of living fish without harming them but under some conditions can be harmful and so can be considered facultative parasites. On the skin and gills are Chilodonella piscicola Zacharias, 1894 and Brooklynella hostilis [18] that can damage fish by feeding directly on epithelial cells and cause respectively the diseases, chilodonellosis and brooklynellosis. In the intestinal lumen are species of Balantidium Claparède and Lachmann, 1858, which can cause balantidiasis. A final group feeds on bacteria and mucus on the surface of living fish but under some conditions can damage fish by irritating skin and gill epithelium and so go from being ectocommensals to facultative ectoparasites. This is the case for species of Trichodina Ehrenberg, 1838, which cause trichodiniasis.
Ich and Marine Ich
The most complex and destructive of the fish ciliate pathogens are the obligate parasites, I. multifiliis and C. irritans. Although belonging to different Classes, Oligohymenophorea for I. multifiliis and Prostomatea for C. irritans [1], they have similar polymorphic life cycles [19, 20]. Prey-seeking theronts swim in search of moribund fish host, and although they have fully formed oral features, do not feed. Once a host is found, ciliates may enter the body cavity through the skin or attack the gill epithelium, and upon invasion theronts increase greatly in size forming trophonts. Trophonts begin feeding on tissue and will eventually exit the host and form a divisional cyst, which will ultimately release between 50 and several thousand, for I. multifiliis, daughter cells, or tomites. Tomites are a small life stage that transform soon after release to theronts [19]. Ichthyophthirius multifiliis causes ichthyopthiriosis and has a near universal host range on freshwater fish [21–23]. Ich causes massive economic losses to the aquarium and aquaculture industries. Cryptocaryon irritans causes the marine equivalent of ich and has a more restrictive host range [24]. The species is of a concern in the warm water food fish aquaculture [25, 26].
Scuticociliatosis and Tetrahymenosis
Scuticociliatosis is caused by ciliates belonging to the Subclass Scuticociliatia Small, 1967 and since the 1990s has been a problem around the world for marine fish aquaculture [23]. Impacted industries include Asian Japanese flounder [27, 15, 28], Mediterranean turbot and European seabass [13], and Australian bluefin tuna [29]. Scuticociliatosis is caused by a number of species including Uronema marinum Dujardin, 1841, Miamiensis avidus Thompson and Moewus, 1964 (senior syn. of Philasterides dicentrarchi Dragesco et al., 1995), and Parauronema virginianum Thompson, 1967 [1]. Characteristically marine organisms, recently Pseudocohnilembus persalinus Evans and Thompson, 1964 was isolated from the ovarian fluid of freshwater rainbow trout spawning in Idaho, suggesting a wider range of osmotic tolerance than otherwise expected for scuticociliates [30]. Early infections with scuticociliates may appear as lesions on skin or gill tissue, but the ciliates can quickly infest deeper tissues and organs, including muscle, peritoneal cavity, kidney, pancreas, liver, urinary bladder, spinal cord, and brain [23]. Treatment and control of scuticociliatosis is especially difficult not only because of invasion of the fish host tissues, but also due to their success as free-living bacterivores in the environment [31].
Tetrahymenosis is the freshwater counterpart to scuticociliate-borne infestations. Members of the genus Tetrahymena have long been noted to harm fish [10, 32, 33, 34], although the ciliates are able to grow independently of fish or any other animal and so are considered free-living but facultative endoparasites. In the production of tropical fish for the aquarium trade, tetrahymenosis has caused considerable economic losses [16]. Infected fish have necrotic skin lesions that extend into the musculature, and the pathogen can reach the circulatory system and destroy internal organs. The most frequent reports of infection have been in the guppy, Poecilia reticulata [14] but infections have also been noted in cichlids, black mollies, and tetras [11, 35]. As well, infections have been discovered in fish being farmed for human consumption. Tetrahymenosis appeared to be the cause of rapid perch die-offs in two pond facilities in Australia [36, 37] and Tetrahymena were found in cranial ulcerations in farmed Atlantic salmon in Canada [38]. Ciliates have also been isolated from diseased common carp, catfish, and rainbow trout [39]. Usually the causative agent has been identified simply as belonging to genus Tetrahymena (Tetrahymena sp.). However, in the case of the Atlantic salmon the ciliate was thought to be T. corlissi Thompson, 1955 or possibly T. rostrata Kahl, 1926 [38]. Experimentally, T. pyriformis was found to infect several species of ornamental fish if the fish had received a deep skin wound [40].
Chilodonellosis and Brooklynellosis
Two species of Chilodonella Strand, 1928, C. hexasticha Kiernik, 1909 and C. piscicola cause chilodonellosis in fresh- and brackish water fish, and was at one time the most significant single disease to the commercial tropical fish industry [41]. However, the broad host range is believed to include a universal range of freshwater teleosts [23, 39]. Mass mortalities of feral fish have also been reported resulting from Chilodonella hexasticha infection [42]. These organisms are able to survive by feeding on bacteria when apart from the host, but are facultative ectoparasites, feeding directly on epithelial or gill cells with a penetrating cytostome [43]. In marine fish aquaculture, a similar disease, brooklynellosis, is caused by Brooklynella hostilis and may cause recurrent mass mortalities [44]. Aquarium-raised teleosts are affected similarly to those parasitized with Chilodonella sp., however B. hostilis is restricted to attacking gill tissue where epithelial and blood cells are fed upon resulting in serious lesions [18]. Currently no cases of brooklynellosis have been identified in feral fish, and the ability of B. hostilis to survive free of the host is unknown, however a recently identified addition to the genus, B. sinensis [45], is free-living [45].
Balantidiasis
Balantidiasis has been reported in the intestinal lumen of several fish species and occasionally has been associated with enteric diseases [7]. For example, in the grass carp Balantidium ctenopharyngodonis Chen, 1955 appeared to be the cause of intestinal lesions in older fish [46]. However, the ciliates usually are harmless.
Trichodiniasis
The ectocommensal trichodinids are commonly found gliding along and loosely adhering to fish skin and gills, where they feed on waterborne particulates, bacteria, and fish detritus. Seven genera within the family are known to interact with freshwater or marine fish, the most notable being species of Trichodina (reviewed in [7]), and they are characterized by the presence of an adoral adhesive disk incorporating blade-like denticles. Fish that are stressed due to changing or poor environmental conditions, or juvenile fish, are susceptible to infestation of excessive numbers of trichodinids on the skin or gill surface. Trichodiniasis results from the ciliates repeatedly detaching and reattaching from the host, which causes irritation and damage to epithelial cells [23, 47]. At this point the ciliates may feed on disrupted cells, and subsequent bacterial population, and may be considered facultative ectoparasites.
Ciliates as Fish Passengers and Carcass Consumers
The sessiline peritrich ciliates (Order Peritrichia Stein, 1933) of fish might be considered as passengers or more formally as ectocommensals [7]. They are motile with specialized features for transient adherence to fish surfaces (reviewed in [1, 7]). Among the sessilids found associated with fish, notable species are included within the Families Epistylididae Kahl, 1933; Scyphidiidae Kahl, 1933; Operculariidae Fauré-Fremiet in Corliss, 1979; and Ellobiophryidae Chatton and Lwoff, 1929. Attachment to the host skin or gill surface occurs at the scopula—a thigmotactic structure at the posterior end of the ciliate—either directly, or through a secreted stalk, and ciliates may occur singly or as macroscopic colonies. These ciliates use their hosts as a platform from which they feed on organic debris and bacteria. In general, their numbers are low and they do not damage their host [12].
Ciliates might also have an ecologically important interaction after the death of fish, by whatever cause. Ciliates would be expected to feed on fish carcasses because they have been observed around moribund fish [48] and are known to feed on dead material [4]. However, as a process this activity appears unexplored but could be ecologically important as part of carcass flocculation. Flocculation is the aggregation of fine organic and/or inorganic particulates into larger fragile structures, floc, and comes about as a result of biotic and abiotic processes. Floc is part of industrial systems, such as wastewater treatment, and ecological systems, such as rivers and lakes [49]. Ciliates make a significant contribution to floc formation in sewage treatment plants [50]. In fish biology, the post-spawning die-off of Pacific salmon is an interesting example of floc formation, with fish flocs delivering salmon nutrients to stream beds and freshwater food chains [51].
Ciliates Interacting with Fish Microbial and Viral Pathogens
Increasingly the interaction between ciliates and microbial/viral pathogens of mammals is being studied, and although early work was done with fish pathogens, fish generally have received less attention. The overarching question is whether the transmission of the disease can be potentially modulated by ciliates, and although no definitive answers have been possible to date, several fascinating interactions have been revealed. The ciliate that has been most often been used in these studies has been T. pyriformis, which itself can be pathogenic to fish [40]. Overall, two contrasting themes have emerged: the ciliates either destroy or protect the pathogen.
Ciliates can eat and inactivate bacteria. Often this has been studied to develop predator–prey models for describing sewage treatment plants and aquatic microbial ecosystems [52, 53–55]. The most common combination has been either T. thermophila or T. pyriformis with Escherichia coli. While E. coli is not pathogenic to fish, the species is found in the fish intestine [56], and some E. coli strains are pathogenic to humans [57]. In common co-culture, T. pyriformis greatly reduced but never completely eliminated E. coli populations. A small, stable bacterial population persisted along with the ciliates and might have been supported by products of ciliate lysis and/or metabolism [58, 59]. Interestingly, Matsui et al. [60] noted that when bacteria density was low, the growth of E. coli was stimulated by T. thermophila. One explanation advanced for this surprising result was that T. thermophila metabolites detoxified metabolites of E. coli, which normally inhibited the growth of the bacteria [60].
As well as potentially destroying pathogens, ciliates can enhance the survival of bacterial pathogens in the environment through several mechanisms. This has been demonstrated with several species of Tetrahymena and several human pathogens, including Campylobacter jejuni, Salmonella enterica, and Legionella pneumophila. None of these would be considered fish pathogens, although L. pneumophila is found in freshwater and can kill fish in the laboratory [61]. However, the mechanisms by which the ciliates act likely apply to fish pathogens as well. To date, the mechanisms all require initial internalization by the ciliates. Once engulfed by T. pyriformis, bacterial pathogens, like C. jejuni, remained viable inside and were better able to resist chlorine residuals [62]. Other protective mechanisms involve the subsequent externalization of bacteria by the ciliates either inside vesicles [63, 64] or in pellets [65]. Salmonella enterica released in vesicles were viable, less susceptible to disinfection treatments than free bacteria [63], and had increased acid resistance, which could contribute to the bacteria surviving in humans [66]. Legionella pneumophila were released in pellets as mature intracellular forms and these were more resistant to several stresses than stationary-phase forms [67]. Thus, ciliates could be constantly creating a protected reservoir of pathogenic bacteria.
Some ciliates appear to help bacteria maintain their virulence to animals when the pathogens are external to the host [68]. In fact, exotoxins might have evolved as a bacterial antipredator defense and their actions on vertebrates incidental to their original purpose [69]. Survival within ciliates can correlate with expression of virulence genes, as illustrated with T. thermophila and two strains of the fish pathogen, Aeromonas hydrophila, virulent (J-1) and avirulent (NJ-4) [70]. Inside the ciliates, J-1 up-regulated genes for bacterial virulence factors and survived, whereas NJ-4 did not and was digested. Ciliates and other protozoa have been considered as training grounds for intracellular bacterial pathogens [71]. Therefore, ciliates can serve as experimental surrogates for phagocytic cells of vertebrates and ciliate cultures can be used to explore the cellular basis of bacterial pathogenesis in animals [72].
Ciliates have been investigated sporadically over the last 70 years for possible associations with viruses of humans, and less frequently with viruses of fish, with intriguing but not always definitive results. Perhaps, the first example of this was a study on Stockholm sewage for the possibility that ciliates harbored poliovirus [73]. Subsequently, laboratory investigations were done, commonly with Tetrahymena. The viruses included influenza virus [74], encephalomyocarditis (EMC) virus [75], measles virus [75], vaccinia virus [76], coxsackie B-5 virus [77], adenovirus 3 [78], poliovirus [79], and Simian rotavirus SA11 [80]. Usually the ciliates were either assumed or demonstrated not to support production of these mammalian viruses [78, 80] but sometimes the results were equivocal [75, 77]. What has been clear was that ciliates destroyed or inactivated some viruses of mammals. This was shown for influenza virus by T. geleii Furgason, 1940 (syn. T. pyriformis) and T. pyriformis [74, 81], adenovirus by T. pyriformis [78], and poliovirus by T. pyriformis [79]. For fish, connections between infectious pancreatic necrosis virus (IPNV), the ciliate, Miamiensis avidus, and the seahorse were explored in the laboratory [82]. IPNV did not replicate in the ciliates but could be picked up by the M. avidus and transmitted to the seahorse. Thus, fish provide perhaps the best example of a potential role for ciliates in viral disease transmission.
Ciliates as Fish Food: Protozooplankton–Ichthyoplankton Link
Ciliates are among the protozooplankton that can serve as food for ichthyoplankton. Ichthyoplankton is a metazoan grouping of plankton, consisting of fish larvae, although fish eggs and embryos are often put into this grouping as well. In aquatic food chains, the protozooplankton are the link between bacteria/phytoplankton and metazoan consumers. By consuming bacteria/phytoplankton, planktonic protozoa package the nutrients into larger particles that can be eaten directly by metazoans [83]. Usually the metazoans are considered to be invertebrates such as copepods. However, the protozooplankton can also be eaten by ichthyoplankton. This is the protozooplankton–ichthyoplankton link (reviewed in [84]).
Field studies have demonstrated that ciliates can be food for the larvae of marine fish [84–86]. This demonstration requires examining larval gut contents, which is a difficult task. Usually marine ciliates are grouped into loricate (shell-like outer coverings) and aloricate (naked) forms, with naked forms being more abundant in ocean samples. By contrast, in the gut, the hard parts of the loricate ciliates; such as members of Family Tintinnidae Claparède and Lachmann, 1858; are indigestible, making them easier to see and the first ones to be considered as food for larval fish (Last [87, 88]). Yet, naked ciliate genera have been found in the guts of most fish taxa that have been examined carefully, such as Ammodytidae, Gadidae, and Gobiidae [89]. Ciliates include Myrionecta Jankowski, 1976; Tontonia Fauré-Fremiet, 1914; Leegaardiella Lynn and Montagnes, 1988; Laboea Lohmann; and Strombidium Claparède and Lachmann, 1859 [89]. Thus ciliates are prey items for ichthyoplankton, but the ecological importance of this requires more study [84].
Laboratory studies support the general conclusion of field observations: fish larvae can eat ciliates. Studies have been done with the larvae of Northern anchovy, Engraulis mordax, surgeonfish, Paracanthurus hepatus, Atlantic cod, Gadus morhua, and red snapper, Lutjanus campechanus. When presented as a first food, species of Strombidium Claparède and Lachmann, 1859, were detected immunochemically in larval guts of Northern anchovy [90]. Larval surgeonfish were fed species of Euplotes Ehrenberg, 1830 that had been first allowed to engulf on fluorescent beads (1 μm) [91]. This allowed Euplotes sp. to be easily visualized inside the gut of larvae. Video microscopy of first-feeding larvae of Atlantic cod showed them eating the ciliate Balanion sp. Wulff, 1919, and preferring them over other feed items [92]. The marine ciliate, Fabrea salina Powers and Mitchell, 1910, was used to successfully feed red snapper larvae [17].
Culturing Ciliates Associated with Fish
The culturing of fish ciliates has been done in many ways but collectively media have been developed along three themes. Media can be seen as modifications of either freshwater for freshwater ciliates, seawater for marine ciliates, or vertebrate bodily fluids for ciliates capable of growing inside teleosts. The approximate osmolalities of freshwater, fish blood, and seawater are 50, 300, and 1,000 mOsm/kg H2O, respectively. Some ciliates from one environment are able to grow to varying extents in solutions of other osmolalities [93, 94]. To feed the ciliates and complete the medium, three general types of additions have been made. These are either bacteria, complex but undefined organic supplements, or a set of completely defined organic compounds.
Media Based on Freshwater
Freshwater ciliates have perhaps the longest history of laboratory culture of any eurkaryotic cell. Culturing began over 100 years ago with organic matter decaying in water. In early reports the water was often described as “tap” water but sometimes salts were added to make balanced and buffered salt solutions [95]. The ciliates grew along with other microbes, especially bacteria, and these were often referred to as “infusoria.” They might also be described as agnotobiotic cultures, meaning that the identity of the microbes was unknown or poorly defined. Research on culturing was most intensive with primarily just one ciliate genus, Tetrahymena, with species such as T. pyriformis and T. thermophila often being used to optimize procedures. Subsequently Tetrahymena were grown with known bacterial species, gnotobiotic cultures, and then without any other micorganisms, axenic cultures. For axenic cultures, the nutrients were either complex mixtures of organic molecules or completely defined. These successes led to efforts to scale up the production of Tetrahymena.
Culturing Freshwater Ciliates with Bacteria
A single bacterial species or a combination of several known species has been found to support the growth of Tetrahymena and a few other freshwater, free-living ciliates. In nature, these ciliates feed on a variety of bacteria or other microbes in streams, ponds and lakes, but in cultures the identity of the microbes can be defined. Specific bacteria can be added to crude suspensions of organic matter that alone allow bacteria but not ciliate growth. Examples include sterile 0.10 % hay fusion [96] and 0.15 % cerophyl [54]. The ciliates feed on the growing bacteria population and proliferate. Under some conditions, T. pyriformis simultaneously consumed bacteria and dissolved organic matter [8]. Growth on bacteria alone was demonstrated more conclusively using medium with no energy source and by starting the co-cultures with high densities of bacteria [60, 97]. Greater than 106 E. coli cells/ml were required to support growth [60]. Dead as well as live bacteria could serve as food [98, 99]. Perhaps, the first report of a ciliate being cultured on a single bacteria species was by Hargitt and Fray [96] who grew Paramecium Müller, 1773, on only Bacillus subtilis. Many early studies noted that for a given ciliate some species of bacteria were suitable food organisms while other were not [95]. Some bacteria in fact were toxic to ciliates [100, 101].
Culturing Freshwater Ciliates Axenically
Growing ciliates axenically began in the 1920s [102]. Since then, two research themes have emerged with respect to culturing. One is the simple routine maintenance of Tetrahymena for a wide variety of purposes through the use of complex but undefined supplements in water or a buffered salt solution. This has been referred to as rich axenic nutrient media [103]. The second is growth in completely defined medium for specific experimental goals, such as studying cellular nutrition. For axenic cultures, starting solutions and components are sterilized through autoclaving and filtration.
Complex Undefined Supplements
For the routine maintenance of Tetrahymena, the most common nutrient source is a proteose peptone (PP). PPs are enzymatic digests of protein. A peptic digest of beef extract at ~2 % in water of high purity is a frequent usage [103]. The PP is often supplemented with one or more of yeast extract, liver extract, glucose, and iron [103]. The ciliates grow vigorously in these simple to prepare and inexpensive media. In fact, this consideration was probably critical in making these ciliates a popular laboratory organism, especially T. thermophila [104]. However, defined media also have roles.
Defined Media
A defined medium is one in which all the components are known chemically and in which the cells grow. Such media were developed for Tetrahymena in the early 1950s [105], making ciliates one of the first groups of non-photosynthetic eukaryotic cells to be grown in a defined medium. Yet, for Tetrahymena these media have not received wide usage, in part due to the perceived difficulties in preparing them [106]. Several research groups have worked to improve the original medium [106–110]. The essence of these media is 19 l-amino acids, four ribonucleosides, B-vitamins, salts, trace metals, and glucose. A minor problem has been the observation of “interface-mediated death” in cultures at a low cell density, which results in non-programed cell death due to lysis at the point of contact between the medium and air above [107]. This has been overcome by making several different kinds of additions [111, 112], the simplest being a surfactant like Pluronic F68 [108]. Defined media offer researchers the most control and can be used for many experimental purposes, especially for nutritional and genetic studies [113], and will likely be used more in the future. One interesting future question is how similar the nutritional requirements for Tetrahymena are for the approximately 4,000 other free-living ciliates.
Media Based on Seawater
Source seawater, instant seawater, and artificial seawater have been used to prepare media to culture marine ciliates [114, 115]. The salinity of seawater is ~35 ‰ but varying salinities have been used. In general terms the media have been completed by the addition of bacteria, complex undefined organic supplements, or chemically defined components. Culturing of marine ciliates began later, in the 1960s, and has been less intensive than the culturing of freshwater ciliates.
Culturing with Bacteria
The first attempts at marine ciliate culturing focused on the marine benthic ciliate Uronema marinum and used bacteria as food [116]. Hamilton and Preslan [116] found that proliferation of this ciliate was supported by Serratia marinorubra but only by 2 of 10 unidentified marine bacterial isolates. Subsequently U. marinum was grown monoxenically on Pseudomonas sp. [117] and on Vibrio spp. [118]. In seawater with peptone, Vibrio natriegens also supported U. marinum growth [119, 120]. A Uronema sp. from an infected fish, the silver pomfret (Pampus argenteus), was cultured on brain heart infusion broth in which a Vibrio sp. was growing [121]. Also natural bacterial assemblages from sediment sustained U. marinus [122].
Other marine ciliates have been cultured on bacteria. A mixed bacterial flora supported the growth of several microaerobic ciliates, including species of Paranophrys Thompson and Berger, 1965, and Strombidium [123]. The scutiociliate M. avidus was maintained on autoclaved Vibrio anguillarum [124].
Culturing Axenically
Several marine ciliates have been grown axenically. The seawater has been supplemented with either complex biological extracts or sets of chemically defined compounds.
Complex Undefined Supplements
Several kinds of undefined supplements have been used, often as combinations, to support marine ciliate growth. Cerophyl, PP, trypticase, and yeast nucleic acids were used to grow M. avidus, P. virginianum, and U. marinum [115]. The same species were grown in seawater supplemented with fish tissue homogenates [125]. For these and other marine ciliates, Nerad and Daggett [126] gave directions for the preparation of media that use several different extracts. These include powdered cereal grass leaves, brown rice, yeast extract, and dried seaweed. The authors note that Cerophyl is no longer available from the original manufacturer and was a mixture of powdered wheat, rye, oat, and barley leaves. Some limited success has been achieved in culturing a marine peritrich ciliate using cerophyl supplements [127].
Defined Medium
A defined medium has been developed for at least one marine ciliate. U. marinum grew in a medium that had 17 amino acids, 4 nucleotides, 5 fatty acids, stigmasterol, and 8 vitamins [93]. Growth occurred in complete seawater but was optimal at 25 % seawater. A medium, 1651 MA, similar to this was available from the American Type Culture Collection (ATCC). However, 1651 MA had to be supplemented with FBS to support growth of six ciliates isolated from parasitized turbot [128].
Media Based on Mammalian Body Fluids
For the ciliates that can survive inside teleosts, growth media that reflect the internal milieu of bony fish and support fish cell proliferation in vitro would seem to be appropriate starting points for culturing the ciliates. Unfortunately, a medium has yet to be developed specifically for growing fish cells. However, many media have been developed for mammalian cells. With very little modification, these media work well for the cells of bony fish [129]. A brief review on using these media for teleost cells follows.
Basal Media and Sera for Culturing Teleost Cells
The complete growth medium for the propagation of fish cells has two essential parts, a basal medium and a supplement [129]. A basal medium constitutes an aqueous solution of buffering agents, bulk ions, and nutrients. Basal media have been constructed to mimic in vivo fluids, which would be primarily extracellular fluid and blood plasma [130]. In general, the inorganic blood constituents of mammals and fish are similar, although only a few fish species have been examined [131]. For all basal media the nutrients include a hexose, amino acids, and vitamins, but many variations are possible and available commercially. Most of these, such as minimum essential medium (MEM) or Eagle’s MEM (EMEM), have a sodium bicarbonate (NaHCO3) buffering system, which requires an atmosphere of 5 % CO2. Others are formulated specifically for use in free gas exchange with air. One of these is Leibovitz’s L-15, which maintains physiological pH through a combination of salts, high basic amino acid concentrations, and galactose as the hexose [132]. In order to support the proliferation of fish cell lines, the basal media needs to be supplemented. The most common supplement is a bovine serum, usually fetal bovine serum (FBS). For the routine maintenance of cell lines, antibiotics often are added to media. Usually these are penicillin at 100 I.U. with streptomycin at 100 μg/ml (Pen/Strep). Media like these have formed the basis of attempts to culture several ciliates of fish.
Basal Media and Sera for Culturing Ciliates
Miamiensis avidus (syn. Philasterides dicentrarchi) has been the most intensively studied ciliate for growth in basal media originally developed for mammalian tissue culture. Several additions to L-15 led to the first success [133]. The additions were lipids (lecithin and Tween 80), nucleosides (adenosine, guanosine, cytidine, and uridine), and glucose, and the undefined supplement of 10 % FBS. FBS was essential for growth and the lipids enhanced the growth with FBS. The medium was prepared at 10 and 27 ‰ salinity. Over 7 days the best growth was observed in 10 ‰ salinity at temperatures between 18 and 23 °C. Growth was negligible at 13 °C. A turbot cell homogenate together with FBS and lipids further enhanced growth but not if the homogenate had been autoclaved [134]. The authors suggest that toxic substances released from broken cells impeded growth but were inactivated by autoclaving. In addition to L-15, MEM formed the basis of a culture medium for M. avidus [135]. Growth occurred when the MEM was supplemented with yeast extract and FBS.
Other marine ciliates of fish have been cultured in media based on L-15. With supplements, L-15 allowed the axenic culture for up to 24 months of six Philasterides isolates from parasitized turbot [128]. In L-15 alone, the ciliates survived for long periods but did not proliferate and instead became slender and acquired unusual shapes. Supplementing L-15 with turbot blood cells or fish brain extract supported growth. For convenient routine growth, the best medium was L-15 supplemented with 1.28 % artificial marine salts, 10 % heat-inactivated FBS, 1X Eagle’s basal medium vitamin mixture, and 1 % ribonucleic acid from torula yeast. The final salinity was ~20 ‰. At 20 °C, growth was exponential between 72 and 96 h and stationary phase started at about 120 h. Another ciliate, U. marinum, was cultivated in media based on L-15 [136]. Growth was best in L-15 with FBS, lipids, and an additional supplement of grouper tissue homogenate.
Two mammalian basal media, EMEM and L-15, have been tried with limited success to grow the obligate parasites, I. multifiliis and C. irritans. For I. multifiliis, which is from freshwater, EMEM supplemented in various ways was compared with simpler media, including water [137]. For C. irritans, which is a marine ciliate, media were built on L-15 mixed with seawater [138].
Compared with water, EMEM extended the survival of all stages of I. multifiliis, but failed to support the in vitro growth and development of the parasite [137]. In EMEM, theronts survived up to 5 days. In EMEM with 10 % FBS, trophonts survived for up to 16 days. The theronts failed to transform to trophonts.
L-15/seawater supplemented with sera supported C. irritans tomonts and trophonts in short-term cultures that either had an attachment substrate (solid medium) or not (liquid medium) [138]. Fetal calf serum (FCS), tilapia serum, or grouper serum was compared. The attachment substrate was a strip of trypticase soy agar and was meant to mimic invasion of a fish host. Theronts transformed into trophonts, which enlarged in cultures with ~20 % FCS but not with other sera. Growth occurred in both liquid and solid media. After transformation, trophonts survived from 4 to 13 days. The trophonts that were raised in vitro were not observed to continue development into tomonts. To improve the culturing of C. irritans and I. multifiliis, investigators have explored the use of fish cell lines as support in co-cultures.
Teleost Cell Lines
Cell lines have been reported from about 75 of the estimated 29,000 teleost species. Several compilations of the species and tissue origins of fish cell lines have been published [139–142]. About 283 cell lines have been described and nearly all can be cryopreserved [140]. Many of these likely have been lost over time through inattention [129] but some are available from the ATCC. Formally, cell lines arise when primary cultures are passaged or subcultivated into new culture vessels [143]. For many fish species, serially subcultivating cell cultures leads to the spontaneous immortalization of the cells and a cell line [129]. Some common fish cell lines are bluegill fin (BF-2), Chinook salmon embryo (CHSE-214), fathead minnow (FHM), and EPC, which is now known to be from FHM but is still referred to as EPC [144].
Except for temperature, the environment for culturing piscine and mammalian cells is similar. Nearly all the fish cell lines are adherent, requiring a surface on which to attach, spread and grow. An exception is a monocyte/macrophage cell line, RTS11, from the rainbow trout spleen [145]. Fish adherent cell lines are maintained on conventional tissue culture plastic manufactured for mammalian cells. Fish cell lines grow over wide temperature ranges, which vary with the species and have been referred to as the proliferation zones [146]. Within the proliferation zones, growth is optimal over narrower ranges. For warm-water fish, this is 26–30 °C; for cold-water fish, this is 20–23 °C. The warm water fish cell lines, BF-2 and EPC, and the coldwater fish cell line, CHSE-214, have been explored as agents to support ciliate growth in vitro.
Co-culturing Ciliates and Fish Cell Lines
The maintenance of I. multifilis has been studied in co-culture with piscine cell lines and with fish tissue fragments, including skin, gill, and fin [147–149]. One cell line partner was BF-2 in either EMEM or medium 119 with FBS [148, 149]. BF-2 was compared with explant cultures of channel catfish tissues but the explants were superior in promoting ciliate growth [149]. Theronts attached and penetrated the cells undergoing tissue explant outgrowth and trophonts began to grow. By contrast, theronts swam in the medium of BF-2 cultures, moved on the cell surface, and briefly adhered to the cells, but did not penetrate the monolayer. Although initially supporting trophont development, the co-cultures of tissue fragments and I. multifilis deteriorated over time and the majority of the trophonts were dying by 48 h. The other cell line partner was EPC in either EMEM or L-15 that was supplemented or not with 8 % rainbow trout serum and 8 % mucus [150]. With or without EPC and with or without the supplements, theronts transformed within 2 days into trophont-like stages. However, they grew and survived best when co-cultured with EPC in the presence of rainbow trout serum and mucus. Although surviving for up to 13 days, trophonts never developed into tomonts.
More success has been achieved in co-cultures for the marine counterpart of I. multifiliis. The entire life cycle of C. irritans was supported in a double-layered co-culture system with FHM cells [151]. The system had as a medium, L-15 with FBS and penicillin/streptomycin, and was initiated by having the FHM cells attached and spreaded in culture dishes. After spreading, ultra-low-melting agarose was layered on top of the cells. A micropipette was used to inoculate theronts beneath the agarose and at the center of the cell layer. Over 4 h, theronts transformed to trophonts. Trophonts appeared to detach and eat FHM cells and gradually grew. Some transformed to encysted tomonts. When transferred into seawater, some of the encysted tomonts released theronts. These theronts were able to infect fish. By large culture dishes the theront production could be scaled up [151].
The in vitro growth of the scuticociliate M. avidus has been done conveniently in co-cultures with a salmon cell line. CHSE-214 in EMEM with 10 % FBS supported the growth of M. avidus in co-cultures at 20 °C [152–155].
Uses of Ciliate Cultures in Fish Biology
The culturing of free-living ciliates can allow the study and control of at least four aspects of fish biology. First, they aid the study of ciliates as pathogens and disease-causing agents. Second, they could be exploited to study the contribution that ciliates make to nutrient cycling from fish carcasses. Third, they may be used to study whether ciliates modulate the transmission of viral and microbial pathogens of fish. Finally, cell cultures of ciliates can provide food for early life stages, the protozooplankton-ichthyoplankton link.
Ciliates as Fish Pathogens
Cell cultures can aid the study of the diseases caused by ciliates. As well as supplying the pathogen, they can be used to identify and study, host responses, pathogenic mechanisms, and cures.
Scaling up Production and Attenuation of Pathogens
Cultures can potentially produce large quantities of ciliates for use in experiments to study the development of pathogenicity and immunity in fish. However, culturing has been noted to change the virulence of several pathogenic ciliates. For U. marinum from infected olive flounder, protease activity from short-term cultures was higher than activity from long-term cultures [156]. In the case of Tetrahymena sp. isolates from infected guppies, one isolate, Tet-NI 1, was attenuated after 15 months of culture, whereas another isolate, Tet-NI 6, was cultured for only a few months and remained virulent [157]. For Philasterides, the virulence of two isolates from turbot changed in different ways during long-term culture [128]. After 35–42 passages, isolate A became attenuated, whereas after 20–42 passages, isolate B became more virulent. These phenomena deserve further study and can be exploited to develop vaccination strategies.
Host Responses
The cell culture approach has been used to study responses of fish and identify possible host defense mechanisms. Cysteine proteinases were obtained from M. avidus cultures and added to primary cultures of turbot head kidney leukocytes [158]. The leukocytes increased production of superoxide radicals and of mRNA for the proinflammatory cytokine interleukin-1β, which mediates host inflammatory responses. When in contact with or phagocytosed by M. avidus, turbot leukocytes were activated [159]. Activation included degranulation and a respiratory burst but these activities were insufficient to kill the ciliates. However complement that was activated through the classical pathway was a potent killer of M. avidus [159, 160]. These results suggested that the humoral responses were more important than cellular immune responses in defense against M. avidus [161, 159].
Pathogenic Mechanisms
Ciliate cultures have been used to understand the mechanisms by which the ciliates evade or modulate the defense mechanisms of the host, with a focus on reactive oxygen species (ROS) and proteinases. Host leukocytes produce ROS to generally kill parasites but the ciliates appear to evade ROS by scavenging them. This has been observed in cultures of U. marinum and turbot head kidney leukocytes [162] and of M. avidus and turbot peritoneal leukocytes [163]. Proteinase activities have been examined as virulence factors among Tetrahymena and scutitociliates. For a Tetrahymena sp. pathogenic to guppies, the virulence of different isolates correlated with their cysteine protease [157]. The avirulent Tet-NI 1 had no cysteine protease activity, whereas the virulent Tet-NI 6 had high levels of activity. Cultures of M. avidus have been used as source of proteinases and a way of evaluating the killing activity of sera from fish that had or had not been injected with the ciliate [158, 161]. Through activation of the classical complement pathway, serum from vaccinated turbot killed M. avidus in culture [160, 161]. However, proteinases from P. dicentrarchi were able to degrade the antibodies and factors necessary for complement activation and to decrease killing of the ciliates. Proteases also inhibited leukocyte migration [164] and caused apoptosis of leukocytes [158]. These might be mechanisms by which ciliates evade the fish immune response.
Another mechanism by which histophagous ciliates might survive within their fish hosts is the possession of acid phosphatases. Modulation of phosphotyrosine signaling through protein tyrosine phosphatases (PTPs), such as PTP1b, is emerging as a general point of interaction between host and pathogen [165]. Hints of this are seen with ciliates and fish as well. The pathogenicity of different Tetrahymena isolates correlated with their acid phosphatase activity, in addition to cysteine protease [157]. PTP1b was detected in the plasma membrane of M. avidus [125]. The levels of PTP1b increased upon exposure to grouper skin mucus and leukocytes.
Destruction of fish cells by ciliates has been studied in co-cultures, including an investigation of a possible cytopathogenic mechanism. When a Tetrahymena sp. was co-incubated with a guppy-fin cell line [166] and M. avidus with turbot fibroblast cell line [167], the number of fish cells declined but the decrease was prevented by the addition of the cysteine protease inhibitor E64. These results suggest cysteine proteases have a role in the histolytic activity of the ciliates. In co-cultures with P. dicentrachi, turbot leukocytes served as nourishment, supporting ciliate growth [159], and grouper leukocytes were engulfed by the ciliates, up-regulating ciliate acid phosphatases [125]. The acid phosphatase activities might destroy leukocytes, but other mechanisms might be contributing as well. Work with free-living ameba such as Naegleria fowleri and flagellated protozoa such as Trichomonas vaginalis suggest possible mechanisms to be explored in the future. Amebae can injure and kill mammalian cells in culture by repeated “nibbling” and ingesting plasma membrane fragments [168], a process which has been termed trogocytosis [169, 170]. In co-cultures with bovine epithelial cells, T. vaginalis clustered around the cells, causing mechanical stress, membrane damage and cell death [171]. Only necrotic cells were then phagocytosed by T. vaginalis.
Chemotherapy
Cultures of M. avidus, C. irritans, and Tetrahymena sp. have been used to study the efficacy of chemotherapeutic drugs and treatments. As no effective control exists for M. avidus in fish, many potential drugs have been screened for their effects on these ciliates in culture [124, 135, 166, 172–175]. Formalin, hydrogen peroxide, and resveratrol were among 17 drugs found to be toxic to the ciliates. However, unlike formalin and resveratrol susceptibility, the susceptibility to hydrogen peroxide varied considerably between different P. dicentrarchi isolates [172]. Other toxic treatments were chitosan microspheres cross linked with glutaraldehyde and containing beta-cyclodextrin [174] and the anti-inflammatory drug indomethacin [175]. Indomethacin reduced the growth of P. dicentrachi and eventually caused cell death, possibly by apoptosis [175]. For C. irritans, the double-layered co-culture system with FHM cells was used to study the effects of antiprotozoal compounds on trophonts [176]. This identified sodium salinomycin as a candidate drug for the control of C. irritans infections. In another study on C. irritans cultures, epigallocatechin gallate was found to kill theronts [177]. For Tetrahymena sp., the toxicity of eight chemicals to the ciliates was evaluated in cultures [166]. The most potent compound was niclosamide. These studies show the promise of the cell culture approach for identifying effective treatments and likely will be used more in the future.
Ciliates as Fish Passengers and Carcass Consumers
The process by which peritrich ciliates attach to fish is largely unknown but might be explored through organ cultures of fish skin or gill as well as cell cultures. Organ cultures would be maintained in very similar media as used for cell cultures and might allow attachment to be recorded more easily than on swimming fish.
Cell cultures also could be used in the future to study the role of ciliates independent of other microbes in the formation of fish floc. Ciliate cultures have been used for this purpose in the context of wastewater treatment. Capsule secretion by T. thermophila was shown to contribute to flocculation [178]. Ciliates have been grown in the laboratory on an undefined fish powder [179], fish homogenates [134, 136] and on fish cells [159], so fish fragments might be expected to support ciliate cultures. These cultures could be used to investigate mechanisms by which ciliates form floc and the nature of the floc from fish tissues.
Ciliates Interacting with Fish Microbial and Viral Pathogens
Culturing fish pathogens and ciliates together has been done on just a few occasions but should be a useful approach in the future for studying how ciliates can modulate the interactions between pathogens and fish. Ciliate cultures have not been used to study fish viruses, although mammalian viruses have been studied on many occasions as pointed out earlier in Sects. 1, 2, and 3. However, they have been used on one occasion to study a fungal pathogen, the microsporidian Glugea hertwigi [180], and in a preliminary study on a bacterial pathogen, Yersinia ruckerii [181], which is presented here. Microsporidia need to infect animal cells, including fish cells [182], in order to complete their life cycle. Outside host cells, microsporidia exist as spores. Tetrahymena was shown to phagocytose G. hertwigi spores, but once inside ciliate lysosomes, G. hertwigi blocked lysosomal acidification [183], which is necessary for intracellular digestion and might be expected to inactivate spores. Whether internalization under these circumstances protects and moves spores in the environment would be interesting to explore. Internalization of Y. ruckerii does lead to digestion as shown in Fig. 1. Yersinia ruckerii is the cause of enteric redmouth disease [181] and a strain expressing green fluorescent protein (gfp) has been constructed [184]. With other bacteria expressing gfp, engulfment and digestion by ciliates have been visualized by the appearance and disappearance of fluorescence over time [185]. This was also observed in cultures of gfp Y. ruckeri and T. thermophila (Pinheiro and Bols, unpublished results, Fig. 1), which suggests that this ciliate engulfs and digests this fish bacterial pathogen.

Co-culture of Tetrahymena thermophila and Yersinia ruckeri. a Mixtures of T. thermophila and gfp-expressing Y. ruckeri were established and changes in fluorescence (480 nm excitation/530 nm emission) monitored over a 6 h period. Ciliate- and bacteria-free controls did not show decreases over this period, however the presence of ciliates together with bacteria quickly reduced fluorescence. These data are consistent with other examples of easily phagocytosed and digested bacterial prey [185]. Initial densities of organisms were 50,000 ciliates mL−1, and 106 cfu mL−1 of bacteria. b When viewed by epifluoresence microscopy, ciliates could be seen filled with fluorescent particles, localized to areas characteristic of food vacuoles. The images are taken soon after initiaion of co-culture of bacteria and ciliate, and this fluorescence was no longer visible by 24 h after mixture
Ciliates as Fish Food: Protozooplankton–Ichthyoplankton Link
The practical application of ciliates as fish larval feed has been considered for a long time and has become an established protocol for some species [84]. Thinking on this subject has its origins in the 19th century (see review by [186]) and has mostly focused on larvae of marine fish that have the potential for aquaculture. For some larval marine fish, the small mouth opening makes ciliates an attractive alternative to other feed such as rotifers [187]. However, the most routine use of ciliates in fish husbandry has been with a freshwater species, zebrafish (Danio rerio). Zebrafish are used both by aquarists and by researchers, and zebrafish larvae are often raised on ciliates [188]. In some cases, ciliates have been collected from the environment for feeding to fish larvae, but in other cases the ciliates have been grown in the laboratory for this purpose. Optimizing the production of ciliates in culture will make their use as larval feed in aquaculture, zebrafish husbandry, and laboratory food chain studies more practical, convenient, and controlled.
Scaling up Cultures of Freshwater Ciliates
The mass cultivation or scale up of freshwater ciliates has been done primarily to produce commercially important enzymes [189–191], but the methods might also be used to produce ciliates for use as fish larval feed. Large-scale production has been explored with T. thermophila, T. pyriformis, and T. rostrata [192]. Usually a variety of undefined supplements have been tested to reduce or replace peptones so that growth can be maximized but costs for running large reactors can be kept low. Good growth has been achieved with skimmed milk [193], hemoglobin hydrolysate [192], and yeast extract [194]. Other undefined supplements have been less successful [179]. In medium with a fish powder supplement, the ciliates only grew slightly. This was attributed to the loss of amino acids during fish powder preparation [179]. On the other hand, yeast extract together with 0.5 % glucose supported ciliate densities as high as 1.7 × 106 cells/mL in 100 L fermentors (bio-reactors) at 28 °C [189]. Other milestones in Tetrahymena scale up have been the use of a defined medium and of an airlift reactor with a working volume of 1,200 L [195].
However, the culturing of ciliates for use as feed might require more than just scaling up production. For the rearing of zebrafish larvae, both Paramecium multimicronucleatum Powers and Mitchell, 1910, and T. pyriformis were helpful feed supplements [188, 196]. However, Tetrahymena were easier to grow, and Paramecium appeared to have some nutritional limitations. Paramecium alone did not satisfy the nutritional requirements for the transition of zebrafish from larvae to juveniles [188]. The suggestion was made that because the ciliates had been grown up on bacteria they lacked essential minerals such as iodine for the larval/juvenile transition [188]. Clearly, the culturing of ciliates to support fish larval growth has several fascinating scientific issues, but the engineering of ciliate production on a large scale is being developed.
Scaling up Cultures of Marine Ciliates
For saltwater, one of the most intensively studied ciliates is the hypersaline ciliate F. salina [197]; Pandey et al. [17, 198]. Different feeds for the ciliate have been tried, including egg custard, yeast, and nanoplanktonic green algae. Egg custard (10 mg/mL) yielded the highest densities, 90 ciliates/mL [198]. Mass cultures of 300 L were achieved but the maximal population growth was found in 200 L cultures [198].
Summary and Future
The culturing of some free-living ciliates, such as T. thermophila, has a long history and is easy, whereas culturing some pathogenic ciliates, such as Ichthyophthirius multifiliis, is difficult, but for both of these extremes there are many interesting issues to explore in the future. The culturing of fish ciliates can be viewed as being done in media that is based on either freshwater, seawater, or vertebrate bodily fluids together either with bacteria, fish cells, or organic matter that is either undefined, such as PP, or defined. For species like T. thermophila, the end goal of straightforward culturing has been achieved but questions could still be asked about monoxenic and axenic cultures of this species. In monoxenic cultures are E. coli the best food or do other bacterial species, or even other microbes such as yeast, support better growth? In axenic cultures, the components of defined media might still be optimized to support more vigorous growth. For obligate parasites with complex life cycles, such as I. multifiliis, more sophisticated culture systems need to be developed to obtain all stages. Possibly this requires improvements in fish cell culturing because a particular ciliate life cycle stage might need a specific fish cell type. In final analysis, some parasites might prove impossible to culture but efforts to do so can nonetheless provide insights into their life cycle.
The culturing of fish ciliates aids studies of ciliate/fish interactions and holds out the possibility of revealing new ones. Cultures allow many aspects of ciliate pathogenicity to be examined. These include how histophagous ciliates kill fish cells and what drugs might protect the fish cells from the histophages. Cultures could be used to examine processes after fish death because ciliates can eat dead matter and might participate in flocculation of fish carcasses, which is important in nutrient cycling. Cultures can be used to study the relationships of free-living ciliates, and even pathogenic ones, with other fish pathogens, viral, bacterial, or fungal. This approach is only beginning but promises to reveal the contrasting actions of ciliates in protecting and inactivating pathogens. This knowledge could be used to understand possible roles of ciliates in disease transmission and how to control the spread of diseases. Cultures could be a source of feed for fish larvae because large-scale cultures of both freshwater and marine ciliates have been achieved. Finally, cultures might be used to delineate new relationships, with xenohormesis being an example. Xenohormesis is the sensing of chemical cues from one species by another [199]. This could reveal new compounds that might be used to promote fish growth and health.
References
Lynn DH (2008) The ciliated Protozoa: characterization, classification, and guide to the literature, 3rd edn. Springer, New York
Finlay BJ, Corliss JO, Esteban G, Fenchel T (1996) Biodiversity at the microbial level: the number of free-living ciliates in the biosphere. Q Rev Biol 71:221–237
Sanders RW (2011) Alternative nutritional strategies in protists: symposium introduction and a review of freshwater protists that combine photosynthesis and heterotrophy. J Eukaryot Microbiol 58(3):181–184
Jones AR (1974) The Ciliates. Hutchinson and Co. Ltd, London
Jarvis BDW, Hungate RE (1968) Factors influencing agnotobiotic cultures of the rumen ciliate, Entodinium simplex. Appl Microbiol 16:1044–1052
Strüder-Kypke MC, Wright AG, Jerome CA, Lynn DH (2001) Parallel evolution of histophagy in ciliates of the genus Tetrahymena. BMC Evolut Biol 1:5
Basson L, Van As J (2006) Trichodinidae and other ciliophorans (Phylum Ciliophora). In: Woo PTK (ed) Fish diseases and disorders, vol 1, 2nd edn. CAB International Publishing, Oxon, pp 154–182
Glaser D (1988) Simultaneous consumption of bacteria and dissolved organic matter by Tetrahymena pyriformis. Microbiol Ecol 15:189–201
Orias E, Rasmussen L (1976) Dual capacity for nutrient uptake in Tetrahymena. IV. Growth without food vacuoles and its implications. Exp Cell Res 102:127–137
Corliss JO (1960) Tetrahymena chironomi sp. nov., a ciliate from the midge larvae and the current status of facultative parasitism in the genus Tetrahymena. Parasitology 50:111–153
Hoffman GL, Landolt M, Camper JE, Coast DW, Stockey JL, Burek JD (1975) A disease of freshwater fish caused by Tetrahymena corlissi Thompson, 1955, and a key for the identification of holotrich ciliates of freshwater fish. J Parasitol 61:217–233
Lom JR, Corliss JO (1968) Observations on the fine structure of two species of the peritrich ciliate genus Scyphidia and on their mode of attachment to their host. Trans Am Microsc Soc 87:493–509
Iglesias R, Paramá A, Álvarez MF, Leiro J, Fernández J, Sanmartin ML (2001) Philasterides dicentrarchi (Ciliophora, Scuticociliatida) as the causative agent of scuticociliatosis in farmed turbot Scophthalmus maximus in Galicia (NW Spain). Dis Aquat Org 46:47–55
Imai S, Tsurimaki S, Goto E, Wakita K, Hatai K (2000) Tetrahymena infection in guppy, Poecilia reticulata. Fish Pathol 35:67–72
Kim SM, Cho JB, Kim SK, Nam YK, Kim KH (2004) Occurrence of scuticociliatosis in olive flounder Paralichthys olivaceus by Philasterides dicentrarchi (Ciliophora: Scuticociliatida). Dis Aquat Org 62:233–238
Pimenta Leibowitz M, Ariav R, Zilberg D (2005) Environmental and physiological conditions affecting Tetrahymena spp. infection in guppies, Poecilia reticulata Peters. J Fish Dis 28(9):539–547
Rhodes MA, Phelps RP (2008) Evaluation of the ciliated protozoa, Fabrea salina as a first food for larval red snapper, Lutjanus campechtanus in a large scale rearing experiment. J Appl Aquac 20:120–133
Lom JR, Nigrelli RF (1970) Brooklynella hostilis, n.g., n.sp., a pathogenic cyrtophorine ciliate in marine fishes. J Protozool 17:224–232
Buchmann K, Sigh J, Nielsen CV, Dalgaard M (2001) Host responses against the fish parasitizing ciliate Ichthyophthirius multifiliis. Vet Parasitol 100(1–2):105–116
Colorni A (1985) Aspects of the biology of Cryptocaryon irritans, and hyposalinity as a control measure in cultured gilt-head sea bream Sparus aurata. Dis Aquat Org 1:19–22
Dickerson HW, Dawe DL (2006) Ichthyophthirius multifiliis and Cryptocaryon irritans. In: Woo PTK (ed) Fish diseases and disorders, vol 1, 2nd edn. CAB International Publishing, Oxon, pp 116–153
Matthews RA (2005) Ichthyophthirius multifiliis Fouquet and ichthyophthiriosis in freshwater teleosts. Adv Parasitol 59:159–241
Noga EJ (2010) Fish disease: diagnosis and treatment, 2nd edn. Wiley-Blackwell Publishing, Ames, pp 129–144
Colorni A, Burgess P (1997) Cryptocaryon irritans Brown 1951, the cause of “white spot disease” in marine fish: an update. Aquar Sci Conserv 1:217–238
Montero FE, Cuadrado M, Crespo FP, Raga JA (2007) Cryptocaryon irritans and Enteromyxum leei, two threats for the culture of Diplodus puntazzo in the mediterrranean. Bull Eur Assoc Fish Pathol 27:242–249
Tookwinas S (1990) Larviculture of seabass (Lates calcarifer) and grouper (Epinephelus malabaricus) in Thailand. Advances in tropical aquaculture, AQUACOP, IFREMER, Actes de Colloque 9, Tahiti, February 20–March 4, 1989, pp 645–659
Yoshinaga T, Nakazoe JI (1993) Isolation and in vitro cultivation of an unidentified ciliate causing scuticociliatosis in Japanese flounder (Paralichthys olivaceus). Gyobo Kenkyu 28:131–134
Kim SM, Cho JB, Lee EH, Kwon SR, Kim SK, Nam YK, Kim KH (2004) Pseudocohnilembus persalinus (Ciliophora: Scuticociliatida) is an additional species causing scuticociliatosis in olive flounder (Paralichthys olivaceus). Dis Aquat Org 62:239–244
Munday BL, O’Donoghue PJ, Watt M, Rough K, Hawkesford T (1997) Fatal encephalitis due to the scuticociliate Uronema nigricans in sea-caged, southern bluefin tuna Thunnus maccoyii. Dis Aquat Org 30:17–25
Jones SR, Prosperi-Porta G, LaPatra SE (2010) First isolation of Pseudocohnilembus persalinus (Ciliophora: Scuticociliatida) from freshwater-reared rainbow trout, Oncorhynchus mykiss. J Parasitol 96:1014–1016
Jee BY, Kim YC, Park MS (2001) Morphology and biology of parasite responsible for scuticociliatosis of cultured olive flounder Paralichthys olivaceus. Dis Aquat Org 47:49–55
Nigrelli RF, Jakowska S, Padnos M (1956) Tetrahymena as pathogenic epibiont in fishes and urodeles. J Protozool 3:10
Speidel CC (1954) Some histological changes in tissues of tadpoles under attack by ciliated protozoa. Anat Rec 120:756
Stolk A (1960) Glaucoma sp. in the central nervous system of carp. Nature 184:1737
Johnson SK (1978) Tet disease of tropical fishes and an evaluation of correction techniques. A & M University Fish Disease Diagnostic Laboratory Leaflet F12, pp 7
Callinan RB, Rowland SJ (1994) Diseases of silver perch. In: Rowland SJ, Bryant C (eds) Proceedings of silver perch aquaculture workshops. Austasia Aquaculture and NSW Fisheries, Sydney, pp 67–75
Herbert B, Graham P (2008) Tetrahymenosis, columnaris disease and motile aeromonad septicaemia in golden perch, Macquaria ambiqua (Richardson), from Australia. In: Bondad-Reantaso MG, Mohan CV, Crumlish M, Subasinghe RP (eds) Diseases in Asian aquaculture VI. Fish Health Section,Asian Fisheries Society, Manila, pp 179–192
Ferguson HW, Hick BD, Lynn DH, Ostland VE, Bailey J (1987) Cranial ulceration in Atlantic salmon Salmo salar associated with Tetrahymena sp. Dis Aquat Org 2:191–195
Shulman SS, Jankovski AV (1984) Phylum Ciliates—Ciliophora Doflein, 1901 (in Russian). In: Shulman SS (ed) Parasitic protozoa, vol 1, Bauer ON (ed) Key to parasites of freshwater fishes of the USSR, vol 140 of keys to the fauna of the USSR. Nauka, Leningrad, pp 252–280
Ponpornpisit A, Endo M, Murata H (2000) Experimental infection of a ciliate Tetrahymena pyriformis on ornamental fishes. Fish Sci 66:1026–1031
Leibovitz L (1980) Chilodonelliasis. J Am Vet Med Assoc 177(3):222–223
Langdon JS, Gudkovs N, Humphrey JD, Saxon EC (1985) Death in Australian freshwater fishes associated with Chilodonella hexasticha infection. Aust Vet J 62:409–413
Wiles MD, Cone D, Odense PH (1985) Studies on Chilodonella cyprini and C. hexasticha (Protozoa: ciliata) by scanning electron microscopy. Can J Zool 63:2483–2487
Lom JR, Dykova I (1992) Protozoan parasites of fishes. Developments in aquaculture and fisheries science, vol 26. Elsevier, Amsterdam
Gong J, Song W (2006) Description of a new marine cyrtophorid ciliate, Brooklynella sinensis n.sp. from the China Sea with a new definition of the genus Brooklynella (Protozoa, Ciliophora, Cyrtophorida). Zootaxa 1113:41–49
Molnar K, Reinhardt M (1978) Intestinal lesions in grass carp Ctenopharyngodon idella (Valenciennes) infected with Balantidium ctenopharyngodonis. Chem J Fish Dis 1:151–156
Lom J (1973) The adhesive disk of Trichodinella epizootica: ultrastructure and injury to the host tissue. Folia Parasitol 20:193–202
Beck BH, Bakal RS, Brunner CJ, Grizzle JM (2006) Virus distribution and signs of disease after immersion exposure to largemouth bass virus. J Aquat Anim Health 18(3):176–183
Liss SN, Droppo IG, Flannigan DT, Leppard GG (1996) Floc architecture in wastewater and natural riverine systems. Environ Sci Technol 30:680–686
Curds CR (1963) The flocculation of suspended matter by Paramecium caudatum. J Gen Microbiol 33:357–363
Petticrew EL, Rex JF, Albers SJ (2011) Biodirectional delivery of organic matter between freshwater and marine systems: the role of flocculation in Pacific salmon streams. J N Am Benthol Soc 30:779–786
Fredrickson AG (1991) Segregated, structured, distributed models and their role in microbial ecology: a case study based on work done on the filter-feeding ciliate Tetrahymena pyriformis. Microb Ecol 22:139–159
Habet M, Alexander M (1978) Protozoan density and the coexistence of protozoan predators and bacterial prey. Ecology 59:140–146
Taylor WD (1978) Growth responses of ciliate protozoa to the abundance of their bacterial prey. Microb Ecol 4:207–214
Vayenas DV, Pavlou S (1999) Chaotic dynamics of a food web in a chemostat. Math Biosci 162:69–84
Hansen DL, Clark JJ, Ishii S, Sadowsky MJ, Hicks RE (2008) Sources and sinks of Escherichia coli in benthic and pelagic fish. J Great Lakes Res 34:228–234
Chaudhuri RR, Henderson IR (2012) The evolution of the Escherichia coli phylogeny. Infect Genet Evolut 12:214–226
Sambanis A, Fredrickson AG (1987) Long-term studies of ciliate-bacterial interactions. J Gen Microbiol 133:1619–1630
Sambanis A, Fredrickson AG (1988) Persistence of bacteria in the presence of viable, nonencysting, bacterivorous ciliates. Microb Ecol 16:197–211
Matsui K, Kono S, Sakei A, Ishii N, Min MG, Kawabata ZI (2000) Direct and indirect interactions for coexistence in a species-defined microcosm. Hydrobiologia 435:109–116
Weeks BA, Sommer SR, Dalton HP (1988) Chemotactic response of fish macrophages to Legionella pneumophila: correlation with pathogenicity. Dis Aquat Org 5:35–38
King CH, Shotts EB, Wooley RE, Porter KG (1988) Survival of coliforms and bacterial pathogens with protozoa during chlorination. Appl Environ Microbiol 54:3023–3033
Brandl MT, Rosenthal BM, Haxo AF, Berk SG (2005) Enhanced survival of Salmonella enterica in vesicles released by a soilborne Tetrahymena species. Appl Environ Microbiol 71(3):1562–1569
Ghaffari P, St. Denis CH, Power ME, Jin X, Tsou V, Bols NC, Tang X (2008) Impact of carbon nanotubes on the ingestion and digestion of bacteria by ciliate protozoa. Nat Nanotechnol 3:347–351
Berk SG, Faulkner G, Garduno E, Joy MC, Orti-Jimenez MA, Garduno RA (2008) Packaging of live Legionella pneumophila into pellets expelled by Tetrahymena spp. does not require bacterial replication and depends on a Dot/Icm-mediated survival mechanism. Appl Environ Micrbiol 74:2187–2199
Rehfuss MYM, Parker CT, Brandl MT (2011) Salmonella transcriptional signature in Tetrahymena phagosomes and role of acid tolerance in passage through the protist. ISME J 5:262–273
Koubar M, Rodier MH, Garduno RA, Frere J (2011) Passage through Tetrahymena tropicalis enhances the resistance to stress and the infectivity of Legionella pneumophila. FEMS Microbiol Lett 325:10–15
Pushkareva V, Ermolaeva S (2010) Listeria monocytogenes virulence factor Listeriolysin O favors bacterial growth in co-culture with the ciliate Tetrahymena pyriformis, causes protozoan encystment and promotes bacterial survival inside cysts. BMC Microbiol 10:26
Lainhart W, Stolfa G, Koudelka GB (2009) Shiga toxin as a bacterial defense against a eukaryotic predator, Tetrahymena thermophila. J Bacteriol 191(16):5116–5122
Li J, Zhang XL, Liu YJ, Lu CP (2011) Development of an Aeromonas hydrophila infection model using the protozoan Tetrahymena thermophila. FEMS Microbiol Lett 316(2):160–168
Molmeret M, Horn M, Wagner M, Santic M, Abu Kwaik Y (2005) Amoebae as training grounds for intracellular bacterial pathogens. Appl Environ Microbiol 71:20–28
Benghezal M, Adam E, Lucas A, Burn C, Orchard MG, Deuschel C, Valentino E, Braillard S, Paccaud JP, Cosson P (2007) Inhibitors of bacterial virulence identified in a surrogate host model. Cell Microbiol 9(5):1336–1342
Kling C, Olin G, Fahraeus J, Norlin G (1942) Sewage as a carrier and disseminator of Poliomyelitis virus. J Intern Med 112(3–4):217–249
Groupe V, Pugh LH (1952) Inactivation of influenza virus and of viral hemagglutinin by the ciliate Tetrahymena geleii. Science 115:307–308
Kovacs E, Bucz B, Kolompar G (1966) Propagation of mammalian viruses in protista I. Visualization of fluorochrome labelled EMC virus in yeast and Tetrahymena. Life Sci 5:2117–2126
Jareno MA (1987) Some ultrastructural-changes produced by vaccinia virus in Onchodromus-acuminatus (ciliata, hypotricha). Acta Protozool 26:9–13
Teras J, Entzeroth R, Scholtyseck E, Kesa L, Schrauf I (1988) Light and electron microscope observation of virus-induced Tetrahymena pyriformis in newborn mice (Mus musculus albinicus) brain. Parasitol Res 74:221–227
Sepp T, Jarvekulg L, Saarma M (1992) Investigations of virus–protozoa relationships in the model of the free-living ciliate Tetrahymena pyriformis and adenovirus type-3. Eur J Protistol 28:170–174
Kim TD, Unno H (1996) The roles of microbes in the removal and inactivation of viruses in a biological wastewater treatment system. Water Sci Technol 33:243–250
Benyahya M, Laveran H, Bohatier J, Senaud J, Ettayebi M (1997) Interactions between the ciliated protozoan Tetrahymena pyriformis and the simian rotavirus SA11. Eur J Protistol 33:211–213
Groupe V, Hermann EC, Rauscher FJ (1955) Ingestion and destruction of influenza virus by free living ciliate Tetrahymena pyriformis. Proc Soc Exp Biol Med 88:479–482
Moewus-Kobb L (1965) Studies with IPN virus in marine hosts. Ann NY Acad Sci 126:329–342
Gifford DJ (1991) The protozoan–metazoan trophic link in pelagic ecosystems. J Protozool 38:81–86
Montagnes DJS, Dower JF, Figueiredo GM (2010) The protozooplankton–ichthyoplankton trophic link: an overlooked aspect of aquatic food webs. J Euk Microbiol 57:223–228
Fukami K, Watanabe A, Fujita S, Yamaoka K, Nishijima T (1999) Predation on naked protozoan microzooplankton by fish larvae. Mar Ecol Prog Ser 185:285–291
Nagano N, Iwatsuki Y, Okazaki Y, Nakata H (2001) Feeding strategy of Japanese sand lance larvae in relation to ciliate protozoa in the vicinity of a thermohaline front. J Oceanography 57:155–163
Last JM (1978) The food of four species of pleuronectiforn larvae in the eastern English Channel and southern North Sea. Mar Biol 45:359–368
Last JM (1978) The food of three species of gadoid larvae in the eastern English Channel and southern North Sea. Mar Biol 48:377–386
Figueiredo GM, Nash RDM, Montagnes DJS (2007) Do protozoa contribute significantly to the diet of larval fish in the Irish Sea? J Mar Biol Assoc 87:843–850
Ohman MD, Theilacker GH, Kaupp SE (1991) Immunocytochemical detection of predation on ciliate protists by larvae of the Northern Anchovy (Engraulis mordax). Biol Bull 181:500–504
Nagano N, Iwatsuki Y, Kamiyama T, Nakata H (2000) Effects of marine ciliates on survivability of the first-feeding larval surgeonfish, Paracanthurus hepatus: laboratory rearing experiments. Hydrobiologia 432:149–157
Hunt von Herbing I, Gallager SM (2000) Foraging behavior in early Atlantic cod larvae (Gadus morhua) feeding on a protozoan (Balanion sp.) and a copepod nauplius (Pseudodiaptomus sp.). Mar Biol 136:591–602
Hanna BA, Lilly DM (1974) Growth of Uronema marinum in chemically defined medium. Mar Biol 26:153–160
St. Denis CH, Pinheiro MDO, Power ME, Bols NC (2010) Effect of salt and urban water samples on bacterivory by the ciliate, Tetrahymena thermophila. Environ Pollut 158:502–507
Johnson WH (1941) Populations of ciliates. Am Nat 75:438–457
Hargitt GT, Fray WW (1917) The growth of Paramecium in pure cultures of bacteria. J Exp Zool 22:24–55
Watson PJ, Ohtaguchi K, Fredrickson AG (1981) Kinetics of growth of the ciliate Tetrahymena pyriformis on Escherichia coli. J Gen Microbiol 122:323–333
Ducoff HS, Williams SM, Syn TB, Butler BD (1964) Quantitative studies of metabolism in phagotrophic ciliates. I. Growth of Tetrahymena pyriformis W on viable and on killed bacteria. J Protozool 11:309–313
Sambanis A, Duke CH (1993) Glucose-uptake by Tetrahymena pyriformis in batch cultures with nutritive or inert particulate matter. J Environ Sci Health A 28:729–749
Burbanck WD (1942) Physiology of the ciliate Colpidium colpoda. I. The effect of various bacteria as food on the division rate of Colpidium colpoda. Physiol Zool 15:342–362
Kidder GW, Stuart CA (1939) Growth studies on ciliates. II. The food factor in the growth, reproduction and encystment of Colpoda. Physiol Zool 12:341–347
Lwoff A (1923) Sur la nutrition des infusoires. C R Acad Sci Paris 176:928–930
Orias E, Hamilton EP, Orias JD (2000) Tetrahymena as a laboratory organism: useful strains, cell culture, and cell line maintenance. Methods Cell Biol 62:189–211
Asai DJ, Forney JD (1999) Tetrahymena thermophila. Methods Cell Biol 62:1–579
Kidder GW, Dewey VC (1951) The biochemistry of ciliates in pure culture. In: Lwoff A (ed) Biochemistry and physiology of protozoa. Academic Press, New York, pp 323–400
Szablewski L, Andreasen PH, Tiedtke A, Florin-Christensen J, Florin-Christensen M, Rasmussen L (1991) Tetrahymena thermophila: growth in synthetic nutrient medium in the presence and absence of glucose. J Protozool 38:62–65
Hagemeister JJ, Grave M, Kristiansen TB, Assaad F, Hellung-Larsen P (1999) Interface-mediated death of unconditioned Tetrahymena cells: effect of the medium composition. J Eukaryot Microbiol 46:6–11
Hellung-Larsen P (2005) Cloning of Tetrahymena cells in a chemically defined medium is possible in the presence of surfactants or at reduced temperature. J Biotechnol 115:167–177
Holz GG, Erwin J, Rosenbaum N, Aaronson S (1962) Triparanol inhibition of Tetrahymena and its prevention by lipids. Arch Biochem Biophys 98:312–322
Sripati CE (1987) An improved chemically defined medium for mass-cultures of Tetrahymena nutrient uptake and growth-regulation. J Gen Microbiol 133:2581–2588
Christensen ST, Rasmussen L (1992) Evidence for growth-factors which control cell multiplication in Tetrahymena thermophila. Acta Protozool 31:215–219
Kristiansen TB, Hagemeister JJ, Grave M, HellungLarsen P (1996) Surface mediated death of unconditioned Tetrahymena cells: effect of physical parameters, growth factors, hormones, and surfactants. J Cell Physiol 169:139–148
Sanford YM, Orias E (1981) Phenylketonuric Tetrahymena: phenylalaninine hydroxylase mutants and other tyrosine auxotrophs. Proc Natl Acad Sci USA 78:7614–7618
Henglong XU, Shu M, Jiang Y, Gao S, Min GS, Al-Rasheid KAS (2011) Population dynamics of marine ciliate Euplotes vannus (Protozoa, Ciliphora) in different artificial seawaters. Chin J Ocean Limnol 29:109–117
Soldo AT, Merlin EJ (1972) The cultivation of symbiote-free marine ciliates in axenic medium. J Protozool 19:519–524
Hamilton RD, Preslan JE (1969) Cultural characteristics of a pelagic marine hymenostome ciliate, Uronema species. J Exp Mar Biol Ecol 4:90–99
Lee JJ, Tietjen JH, Mastropaolo CA (1971) Axenic culture of the marine hymenostome ciliate Uronema marinum in chemically defined medium. J Protozool 18(Suppl):10
Parker JG (1976) Cultural characteristics of the marine ciliated protozoan, Uronema marinum Dujardin. J Exp Mar Biol Ecol 24:213–226
Crosbie PBB, Munday BL (1999) Environmental factors and chemical agents affecting the growth of the pathogenic marine ciliate Uronema nigricans. Dis Aquat Org 36:213–219
Perez-Uz B (1995) Growth rate variability in geographically diverse clones of Uronema (Ciliphora: Scuticociliatida). FEMS Microbiol Ecol 16:193–204
Al-Marzouk A, Azad IS (2007) Growth kinetics, protease activity and histophagous capability of Uronema sp. infesting cultured silver pomfret Pampus argenteus in Kuwait. Dis Aquat Org 76:49–56
First MR, Hollibaugh JT (2009) The high molecular weight DOC compound, dextran, is ingested by the benthic ciliate Uronema marinum but does not supplement ciliate growth. Aquat Microb Ecol 57:79–97
Bernard C, Fenchel T (1996) Some microaerobic ciliates are facultative anaerobes. Eur J Protistol 32:293–297
Iglesias R, Paramá A, Álvarez MF, Leiro J, Sanmartin ML (2002) Antiprotozoals effective in vitro against the scuticociliate fish pathogen Philasterides dicentrarchi. Dis Aquat Org 49:191–197
Salinas I, Maas EW, Muñoz P (2011) Characterization of acid phosphatases from marine scuticociliate parasites and their activation by host’s factors. Parasitology 138:836–847
Nerad TA, Daggett PM (1992) Cultivation of scuticociliates. In: Lee JJ, Soldo AT (ed) Protocols in protozoology. Allen Press Inc., Lawrence, pp A-50.1–A-50.6
Clamp JC, Coats DW (2000) Planeticovorticella finleyi n.g., n.sp. (Peritrichia, Vorticellidae), a planktonic ciliate with a polymorphic life cycle. Invertebr Biol 119:1–16
Alvarez-Pellitero P, Palenzuela O, Padros F, Sitja-Bobadilla A, Riaza A, Silva R, Aran J (2004) Histophagous scuticociliatids (Ciliophora) parasitizing turbot Scophthalmus maximus: morphology, in vitro culture and virulence. Folia Parasitol 51:177–187
Bols NC, Dayeh VR, Lee LEJ, Schirmer K (2005) Use of fish cell lines in toxicology of fish. In: Moon TW, Mommsen TP (eds) Biochemistry and molecular biology of fishes-environmental toxicology, vol 6. Elsevier Science, Amsterdam, pp 43–84
Ham RG (1984) Formulation of basal nutrient media. In: Barnes DW, Sirbasku DA, Sato GH (eds) Cell culture methods for molecular and cell biology, vol 1. Allan R Liss, New York, pp 2–21
Wolf K, Ahne W (1982) Fish cell culture. In: Maramorosch K (ed) Advances in cell culture, vol 2. Academic Press, New York, pp 305–328
Leibovitz A (1963) The growth and maintenance of tissue/cell cultures in free gas exchange with the atmosphere. Am J Hyg 78:173–180
Iglesias R, Paramá A, Álvarez MF, Leiro J, Aja C, Sanmartin ML (2003) In vitro growth requirements for the fish pathogen Philasterides dicentrarchi (Ciliophora, Scuticociliatida). Vet Parasitol 111:19–30
Castro R, Paramá A, Barja JL, Leiro J, Sanmartin ML, Lamas J (2007) Culture of the histophagous ciliate Philasterides dicentrarchi (Ciliophora: Scuticociliatia) in fish tissue. J Fish Dis 30:239–242
Harikrishnan R, Jin CN, Kim MC, Kim JS, Balasundaram C, Heo MS (2010) Effectiveness and immunomodulation of chemotherapeutants against scuticociliate Philasterides dicentrarchi in olive flounder. Exp Parasitol 124:306–314
Anderson SA, Hulston DA, McVeagh SM, Webb VL, Smith PJ (2009) In vitro culture and cryopreservation of Uronema marinum isolated from farmed New Zealand groper (Polyprion oxygeneios). J Microbiol Methods 79:62–66
Ekless LM, Matthews RA (1993) Ichthyophthirius multifiliis: axenic isolation and short-term maintenance in selected monophasic media. J Fish Dis 16:437–447
Yambot AV, Song YL (2004) Short term in vitro culture of Cryptocaryon irritans, a protozoan parasites of marine fishes. Fish Pathol 39:175–181
Fryer JL, Lannan CN (1994) Three decades of fish cell culture: a current listing of cell lines derived from fishes. J Tissue Cult Methods 16:87–94
Lakra WS, Swaminathan TR, Joy KP (2011) Development, characterization and storage of fish cell lines: a review. Fish Physiol Biochem 37:1–20
Lee LEJ, Dayeh VR, Schirmer K, Bols NC (2009) Applications and potential uses of fish gill cell lines: examples with RTgill-W1. In Vitro Cell Dev Biol Anim 45:127–134
Wolf K, Mann JA (1980) Poikilotherm vertebrate cell lines and viruses: a current listing for fishes. In Vitro 16:168–179
Schaffer WI (1990) Terminology associated with cell, tissue and organ culture, molecular biology and molecular genetics. In Vitro Cell Dev Biol Anim 26:97–101
Winton J, Batts W, deKinkelin P, LeBerrre M, Bremont M, Fijan N (2010) Current lineages of the epithelioma papulosum cyprini (EPC) cell line are contaminated with fathead minnow, Pimephales promelas, cells. J Fish Dis 33:701–704
Ganassin RC, Bols NC (1998) Development from the rainbow trout spleen of a cell line, RTS11, in the monocyte/macrophage lineage. Fish Shellfish Immun 8:457–476
Bols NC, Mosser DD, Steels GB (1992) Temperature studies and recent advances with fish cells in vitro. Comp Biochem Physiol 103A:1–14
Hurley LM (1999) Ichthyophthirius multifiliis Fouquet: development and assessment of in vitro systems for long term maintenance. PhD thesis, University of Plymouth, Plymouth, UK
Pugovkin D, Felleisen R, Wahli T (2001) A PCR-based method for the detection of Tetrahymena crolissi contaminations in Ichthyophthirius multifilis in vitro cultures. Bull Eur Assoc Fish Pathol 21:222–227
Xu D, Klesius PH, Shoemaker CA, Evans JJ (2000) The early development of Ichthyophthirius multifiliis in Channel catfish in vitro. J Aquat Anim Health 12:290–296
Nielsen CV, Buchmann CV (2000) Prolonged in vitro cultivation of Ichthyophthirius multifiliis using an EPC cell line as substrate. Dis Aquat Org 42:215–219
Yoshinaga T, Akiyama K, Nishida S, Nakane M, Ogawa K, Hirose H (2007) In vitro culture technique for Cryptocaryon irritans, a parasitic ciliate of marine teleosts. Dis Aquat Org 78:155–160
Lee EH, Kim KH (2008) Can the surface immobilization antigens of Philasterides dicentrarchi (Ciliophora: Scuticociliatida) be used as target antigens to develop vaccines in cultured fish? Fish Shellfish Immunol 24:142–146
Lee EH, Kim KH (2008) Immobilization antigen-independent protection of olive flounder (Paralichthys olivaceus) against Philasterides dicentrarchi (Ciliophora: Scuticociliatia) infection. Aquaculture 279(1–4):211–213
Lee EH, Kim KH (2009) CpG-ODN increases resistance of olive flounder (Paralichthys olivaceus) against Philasterides dicentrarchi (Ciliophora: Scuticociliatia) infection. Fish Shellfish Immunol 26(1):29–32
Jung SJ, Im EY, Strüder-Kypke MC, Kitamura SI, Woo PTK (2011) Small subunit ribosomal RNA and mitochondrial cytochrome c oxidase subunit 1 gene sequences of 21 strains of the parasitic scuticociliate Miamiensis avidus (Ciliophora, Scuticociliatia). Parasitol Res 108:1153–1161
Kwon SR, Kim CS, Kim KH (2003) Differences between short- and long-term cultures of Uronema marinum (Ciliophora: Scuticociliatida) in chemiluminescence inhibitory activity, antioxidant enzyme and protease activity. Aquaculture 221:107–114
Leibowitz MP, Ofir R, Golan-Goldhirsh A, Zilberg D (2009) Cysteine proteases and acid phosphatases contribute to Tetrahymena spp., pathogenicity in guppies Poecilia reticulate. Vet Parasitol 166(1–2):21–26
Paramá A, Castro R, Arranz JA, Sanmartin ML, Lamas J, Leiro J (2007) Scuticociliate cysteine proteinases modulate turbot leucocyte functions. Fish Shellfish Immunol 23:945–956
Piazzon MC, Wiegertjes GF, Leiro J, Lamas J (2011) Turbot resistance to Philasterides dicentrarchi is more dependent on humoral than on cellular immune responses. Fish Shellfish Immunol 30:1339–1347
Leiro J, Piazzón MC, Budiño B, Sanmartin ML, Lamas J (2008) Complement-mediated killing of Philasterides dicentrarchi (Ciliophora) by turbot serum: relative importance of alternative and classical pathways. Parasite Immunol 30:535–543
Piazzon C, Lamas J, Leiro JM (2011) Role of scuticociliate proteinases in infection success in turbot, Psetta maxima (L.). Parasite Immunol 33:535–544
Kwon SR, Kim CS, Ahn KJ (2002) Proteases in cell lysate of Uronema marinum (Ciliata: Scuticociliatida), an opportunistic pathogen of cultured olive flounder (Paralichthys olivaceus). J Fish Sci Technol 5:145–149
Leiro J, Arranz JA, Iglesias R, Ubeira FM, Sanmartin ML (2004) Effects of the histiophagous ciliate Philasterides dicentrarchi on turbot phagocyte responses. Fish Shellfish Immunol 17:27–39
Paramá A, Castro R, Lamas J, Sanmartin ML, Santamarina MT, Leiro J (2007) Scuticociliate proteinases may modulate turbot immune response by inducing apoptosis in pronephric leucocytes. Int J Parasitol 37:87–95
Heneberg P (2012) Finding the smoking gun: protein tyrosine phosphatases as tools and targets of unicellular microorganisms and viruses. Curr Med Chem 19:1530–1566
Leibowitz MP, Chettri JK, Ofir R, Zilberg D (2010) Treatment development for systemic Tetrahymena sp. infection in guppies, Poecilia reticulata Peters. J Fish Dis 33(6):473–480
Paramá A, Iglesias R, Alvarez MF, Leiro J, Ubeira FM, Sanmartin ML (2004) Cysteine proteinase activities in the fish pathogen Philasterides dicentrarchi (Ciliophora: Scuticociliatida). Parasitol 128:541–548
Sohn H-J, Kim J-H, Shin M-H, Song K-J, Shin H-J (2010) The Nf-actin is an important factor for food-cup formation and cytotoxicity of pathogenic Naegleria fowleri. Parasitol Res 106:917–924
Brown T (1978) Observations by light microscopy on the cytopathogenicity of Naegleria fowleri in mouse embryo cell cultures. J Med Microbiol 11:249–259
Brown T (1979) Inhibition by amoeba-specific antiserum and by cytochalasin B of the cytopathogenicity of Naegleria fowleri in mouse embryo cell cultures. J Med Microbiol 12:355–362
Midlej V, Benchimol M (2010) Trichomonas vaginalis kills and eats—evidence for phagocytic activity as a cytopathic effect. Parasitology 137:65–76
Budiño B, Pata MP, Leiro J, Lamas J (2012) Differences in the in vitro susceptibility to resveratrol and other chemical compounds among several Philasterides dicentrarchi isolates from turbot. Parasitol Res 110(4):1573–1578
Jin CN, Harikrishnan R, Moon YG, Kim MC, Kim JS, Balasundaram C, Heo MS (2010) Effectiveness of chemotherapeutants against scuticociliate Philasterides dicentrarchi, a parasite of olive flounder. Vet Parasitol 168:19–24
Paramá A, Luzardo A, Blanco-Méndez J, Sanmartin ML, Leiro J (2005) In vitro efficacy of glutaraldehyde-crosslinked chitosan microspheres against the fish-pathogenic ciliate Philasterides dicentrarchi. Dis Aquat Org 64:151–158
Paramá A, Piazzon MC, Lamas J, Sanmartin ML, Leiro J (2007) In vitro activity of the nonsteroidal anti-inflammatory drug indomethacin on a scuticociliate parasite of farmed turbot. Vet Parastiol 148:318–324
Yoshinaga Y, Im HJ, Ogawa NK (2011) In vitro and in vivo efficacy of ionophores against Cryptocaryon irritans. Aquaculture 321:167–172
Picón-Camacho SM, Ruiz de Ybáñez MR, Holzer AS, Arizcun Arizcun M, Muñoz P (2011) In vitro treatments for the theront stage of the ciliate protozoan Cryptocaryon irritans. Dis Aquat Org 94:167–172
Arregui L, Serrano S, Linares M, Perez-Uz B, Guinea A (2007) Ciliate contributions to bioaggregation: laboratory assays with axenic cultures of Tetrahymena thermophila. Int Microbiol 10:91–96
Calilliere-Ethuin P, Duyme F, Tonon F, Jeanfils J, De Coninck J (1998) Optimization of Tetrahymena rostrata growth using food by-products as nitrogen source. Biotechnol Tech 12:177–181
Legault RO, Delisle C (1967) Acute infection by Glugea hertwigi Weissenberg in young-of-the-year rainbow smelt Osmerus eperlanus mordax (Mitchill). Can J Zool 45(6):1291–1292
Tobback E, Decostere A, Hermans K, Haesebrouck F, Chiers K (2007) Yersinia ruckeri infections in salmonid fish. J Fish Dis 30:257–268
Monaghan SR, Kent ML, Watral VG, Kaufman RJ, Lee LEJ, Bols NC (2009) Animal cell cultures in microsporidial research: their general roles and their specific use for fish microsporidia. In Vitro Cell Dev Biol Anim 45:135–147
Weidner E, Sibley LD (1985) Phagocytosed intracellular microsporidiaan blocks phagosome acidification and phagosome–lysosome fusion. J Protozool 32:311–317
Welch TJ, Wiens GD (2005) Construction of a virulent, green fluorescent protein-tagged Yersinia ruckeri and detection in trout tissues after intraperitoneal and immersion challenge. Dis Aquat Org 67:267–272
Power ME, Sotornik D, Pinheiro MDO, Dayeh VR, Butler BJ, Slawson R, Lee LEJ, Lynn DH, Bols NC (2006) Development of a fluorescent multiwell assay for evaluating the capacity of the ciliated protozoan Tetrahymena for bacterivory in water samples. Water Qual Res J Canada 41(3):307–315
May RC (1970) Feeding larval marine fishes in the laboratory: a review. Calif Mar Res Comm CalCOFI Rep 14:76–83
Decamp O, Moss S, Nagano N (2001) Live protozoa: suitable live food for larval fish and shrimp? Glob Aquac Advocate 4:28–29
Lawrence C (2007) The husbandry of zebrafish (Danio rerio): a review. Aquaculture 269:1–20
deConninck J, Verdier-Denantes I, Duyme F, Bouquelet S, Dumortier V (2000) Modeling Tetrahymena thermophila growth and protease production. J Ind Microbiol Biotechnol 25:58–61
Kiy T (1998) Heterotrophic protists—a new challenge in biotechnology. Protist 149:17–21
Munro IG (1985) Protozoa as source of commercially produced enzymes: a review. Process Biochem 20:139–144
Dive D, Piot JM, Sannier F, Guillochon D, Charet P, Lutrat S (1989) Use of hemoglobin enzymic hydrolysates, prepared on a pilot-plant scale, as a nitrogen source for the cultivation of three species of Tetrahymena. Enzyme Microb Technol 11:165–169
Saliba R, Dive D, Devis R (1983) An inexpensive and practical medium for rearing mass-cultures of Tetrahymena. Protistologica 19:417–421
Ethuin P, de Connick J (1996) Scaling up for mass cultivation of the temperature-sensitive mutant Tetrahymena thermophila SJ180. Biotechnol Tech 10:507–512
Hellenbroich D, Valley U, Ryll T, Wagner R, Tekkanat N, Kessler W, Ross A, Deckwer WD (1999) Cultivation of Tetrahymena thermophila in a 1.5-m3 airlift bioreactor. Appl Microbiol Biotechnol 51:447–455
Speksnijder JE, Bijmolt E (1995) Tetrahymena: an easy alternative for Paramecium. Zebrafish Sci Monitor 3:4
Guermazi W, Elloumi J, Ayadi H, Bouain A, Aleya L (2008) Rearing of Fabrea salina Henneguy (Ciliophora, Heterotrichida) with three unicellular feeds. C R Biol 331(1):56–63
Pandey BD, Yeragi SG (2004) Preliminary and mass culture experiments on a heterotrichous ciliate, Fabrea salina. Aquaculture 232:241–254
Howitz KT, Sinclair DA (2008) Xenohormesis: sensing the chemical cues of other species. Cell 133(3):387–391
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pinheiro, M.D.O., Bols, N.C. Use of Cell Cultures to Study the Interactions of Ciliates with Fish. Springer Science Reviews 1, 95–113 (2013). https://doi.org/10.1007/s40362-013-0008-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40362-013-0008-5