
Abstract
Prolific breeding and production of high percentage of recruits are main problems in mixed-sex Nile tilapia (Oreochromis niloticus) culture in earthen ponds. The current study assessed the efficiency of different sizes of African sharptooth catfish (Clarias gariepinus) in controlling recruitment in mixed-sex Nile tilapia cultured in tanks and its effect on growth performance, percentage survival and yield. Three treatments consisting of triplicate ponds stocked with mixed-sex Nile tilapia in monoculture and in polyculture with large (62.50 ± 3.26 g) and small (40.00 ± 2.68 g) African sharptooth catfish were fed on a 297.50 g kg−1 crude protein diet for 126 days. Results showed that, the number of recruits was significantly lower in larger African sharptooth catfish predator than smaller ones and monoculture of Nile tilapia. Polyculture with larger African sharptooth catfish resulted in significantly higher growth performance of Nile tilapia. Large African sharptooth catfish in polyculture reduced the amount of small, low-value recruits, while the yield of large and high value Nile tilapia was increased. This study revealed that fish farmers can reduce prolific breeding, obtain higher growth performance and produce larger size of marketable Nile tilapia by predominantly stocking ponds with large African sharptooth catfish predator of at least 60 g.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Nile tilapia (Oreochromis niloticus, Linnaeus) is one of the most cultured species in the world (FAO 2012) providing major sources of animal protein and income (Chakraborty et al. 2011; Yakubu et al. 2012) in the majority of tropical countries (Lowe et al. 2012). Production of Nile tilapia is carried out in ponds, cages, tanks and raceways through mixed-sex monoculture, all-male monoculture and polyculture with predators (Hernández et al. 2014). Its production in developing countries occurs primarily in semi-intensive ponds with fertilization and/or supplementary feeding. In Tanzania, more than 95 % of fish farmers culture Nile tilapia in earthen ponds under mixed-sex culture (Kaliba et al. 2006).
One of the main problems facing the culture of mixed-sex Nile tilapia in earthen ponds is early maturation leading to prolific breeding and production of high percentage of recruits (Offem et al. 2009; Chakraborty et al. 2011; Yakubu et al. 2012; Dagne et al. 2013). As a result, the total biomass at harvest comprises 28–70 % of small-size, low-value fish originating from spontaneous breeding (de Graaf et al. 1996; Ofor et al. 2011). The increased number of recruits in mixed-sex culture causes stunted growth due to shortage of space and competition for food (Yi et al. 2002; Abdel-Tawwab 2005; Offem et al. 2009).
Various methods for controlling recruitment in mixed-sex Nile tilapia have been proposed. The methods and their limitations have been reviewed by Mair and Little (1991). The methods include culture with predators, culture in cages, intermittent harvesting, as well as hybridization, induction of sterility and production of mono-sex progenies. However, most of these methods are not only expensive to the local fish farmers in Tanzania, but also increase the production cost and limit the expansion of the aquaculture industry (Carlberg et al. 2000). Using a predatory fish to control recruitment in mixed-sex Nile tilapia cultured in ponds is perceived as one of the most appropriate, safe and affordable method in developing countries (Abdel-Tawwab 2005; Offem et al. 2009). However, the efficiency of a predator fish is determined by its capacity to prey upon the recruits of Nile tilapia (de Graaf et al. 1996; Hernández et al. 2014).
African sharptooth catfish (Clarias gariepinus, Burchell) is one of the commonly used predators in controlling recruits in mixed-sex Nile tilapia culture (Lin and Diana 1995; de Graaf et al. 1996). Stocking African sharptooth catfish at a proper size reduces the number of recruits in mixed-sex culture of Nile tilapia, enhances growth performance and produces large marketable Nile tilapia sizes due to adequate food and space (Offem et al. 2009). It has been shown that, recruitment of Nile tilapia can be completely controlled by large (6.8–130 g) rather than small-size (<3.65 g) African sharptooth catfish (de Graaf et al. 1996). However, in certain circumstances, African sharptooth catfish has been criticized as a lazy predator with a low predatory efficiency in controlling recruits in mixed-sex Nile tilapia (de Graaf et al. 1996; Yi et al. 2002; Ofor et al. 2011). This is due to the limited availability of appropriate large-sized African sharptooth catfish with optimum predation efficiency (Abdel-Tawwab 2005; El-Naggar 2007; Offem et al. 2009). Low predatory efficiency in certain stocks of African sharptooth catfish has resulted in similar yields between polyculture with Nile tilapia and monoculture systems (Offem et al. 2009). Choosing an efficient African sharptooth catfish predator of a specific size capable of completely controlling recruits in mixed-sex Nile tilapia represents a limitation to the success of this technique (Fagbenro 2004). To limit the proliferation of small-size Nile tilapia, it is imperative to stock ponds with predators of an adequate size, as predation is size dependent (Shrestha et al. 2011).
The objective of this study was therefore to assess the efficiency of two sizes of African sharptooth catfish in controlling recruitment in mixed-sex Nile tilapia cultured in concrete tanks. The study also evaluated the growth performance, survival and production of Nile tilapia in monoculture and polyculture with African sharptooth catfish.
Materials and methods
Experimental design
The study was conducted at the Sokoine University of Agriculture (SUA) in Morogoro, Tanzania. Nine outdoor circular concrete tanks each having 7 m2 and mean depth of 1 m were used with fortnightly complete renewal rate of water in each tank. The experiment consisted of three replicates of mixed-sex Nile tilapia in monoculture (as a control) and a polyculture system with African sharptooth catfish. Both the mixed-sex population of Nile tilapia brooders and African sharptooth catfish predators were collected from grow-out ponds at the site and acclimatized in separate tanks for 2 weeks prior to the start of the experiment. The sex ratio of Nile tilapia broodstock was determined by looking at genital papilla prior to stocking to produce homogenous sex ratios across treatments and replicates. This was necessary because the sex ratio has definite impact on the production of offspring (Mair and Van Dam 1996). A homogenous sex ratio of 3:1 equivalent to 12:4 females to males, respectively, was used in all treatments and replicates. In both systems, mixed-sex Nile tilapia brooders and African sharptooth catfish were stocked at a density of 3 fish m−2. The stocking density of 3 fish m−2 used in this study was done intentionally to reflect the practice by rural fish farmers in Tanzania. In the monoculture system, 21 mixed-sex Nile tilapia brooders with an initial average weight (mean ± standard deviation) of 51.80 ± 2.30 g were stocked alone to act as a control of the experiment. In the polyculture system, a stocking ratio of about 3:1 for Nile tilapia and African sharptooth catfish, respectively, was used. Sixteen mixed-sex Nile tilapia brooders with an initial average weight (mean ± standard deviation) of 53.50 ± 4.12 g and five large (62.50 ± 3.26 g) African sharptooth catfish were stocked in the first polyculture tanks. A similar number of mixed-sex Nile tilapia brooders (16) with an initial average weight (mean ± standard deviation) of 52.30 ± 3.58 g and five small African sharptooth catfish (40.00 ± 2.68 g) were stocked in the second polyculture tank. The choice of the two sizes of African sharptooth catfish was based on ease of availability. The experiment lasted for 126 days.
Fish in both culture systems were fed on a 297.50 g kg−1 crude protein diet made using locally available cotton seed cake (683.40 g kg−1 of dry feed), maize bran (315.60 g kg−1) and vitamin premix (1.00 g kg−1) (Table 1) for the entire period of the study. A daily ration of 5 % average body weight of Nile tilapia in monoculture or of the combined body weights of Nile tilapia and African sharptooth catfish in the case of polyculture was adopted. The ration was divided into two portions and fed twice daily in the morning (between 0900 and 1000 hours) and evening (between 1500 and 1600 hours).
Data collection
Before the experiment, all African sharptooth catfish individuals were weighed to the nearest 0.01 g by using a sensitive weighing balance (Electronic Precision Balance Model EJB-KD-3000 g, Endel Global Weighing Company). Before weighing, the specimens were blot dried to ensure accuracy. Thereafter all fishes were weighed every 14 days from each tank. Feeding allowances were calculated on the basis of the weights of the African sharptooth catfish and large Nile tilapia. Feeding regimes were re-computed fortnightly after each sampling using the average weights.
At the end of the experiment, water was completely drained out of each tank and all fishes including the recruits were collected, sorted according to species and size (fingerlings and adults) and counted. Nile tilapia adults and their recruits were weighed individually to determine the final mean weight (FMW, g) and weight classes (g), respectively, at the time of harvesting. African sharptooth catfish, large Nile tilapia and their recruits were then weighed for determination of yield. The yield (g) and tank area (m2) were extrapolated into kilogram (kg) and hectare (ha), respectively, for reporting purposes. Daily weight gain (DWG, g day−1), specific growth rate (SGR, %), percentage survival (SR, %), net fish yield (NFY) and net annual yield (NAY) were calculated using the following formulae:
where W f is the final mean weight (g), W i the initial mean weight (g) and t is the growth period (days).
where N h is the total number of fish harvested and N s is the total number of fish stocked.
where W h is the total weight of fish harvested (kg), W s the total weight of fish stocked (kg) and A is the tank area (ha)
Water quality parameters were measured twice daily in the morning (0830–0900 hours) and in the afternoon (1630–1700 hours) to gain an insight into the prevailing environmental conditions in fish tanks. Temperature, pH and dissolved oxygen (DO) were measured using a multi-probe kit (model KTO, HQ, 40D PHC 101-LD 101-01 by Hach Company Ltd, Loveland, CO, USA).
Statistical analyses
Data are presented as mean ± standard error (SE) of mean values and tested for homogeneity of variance using Levene’s test based on mean values. After confirming homogeneity of variances, one-way analysis of variance (ANOVA) was used to test if there were any significant differences in the number of recruits, their mean weights and their percentage contribution on yield among treatments. The contributions (%) to yield of adult Nile tilapia and African sharptooth catfish were compared using one-way ANOVA and independent-samples t test, respectively. ANOVA was similarly used to identify any significant differences in FMW, NFY and NAY among the treatments. The differences among treatments for DWG and SGR were assessed using repeated measures ANOVA as the same groups of individuals were measured repeatedly over a period of time. The individuals are one factor (usually considered as random and serving as replication) and the time dimension is the second factor, a fixed treatment effect (Winer 1971). When significant differences were detected, Tukey post hoc test was performed to determine specific significant differences among treatments. Percentage data were arcsine-transformed prior to ANOVA and back-transformed afterwards for reporting purposes (Zar 2010). All statistical analyses were performed using SPSS version 20 for Windows. Significant differences were judged at a probability level of p ≤ 0.05 for ANOVA, t test and Tukey tests (Zar 2010).
Results
Number of recruits and their weights
Results on the effect of size of predator on controlling the number of Nile tilapia recruits are shown in Fig. 1. The initial size of the predator African sharptooth catfish had a significant effect on reducing the number of recruits in mixed-sex Nile tilapia culture (F 2,6 = 14.712; p = 0.005). The multiple comparisons test showed significantly lower mean number of Nile tilapia recruits in larger (19.33 ± 3.18) compared to smaller African sharptooth catfish predators (40.33 ± 3.18; p = 0.046) and monoculture of Nile tilapia (240.33 ± 54.77; p = 0.006). Likewise, significantly lower number of Nile tilapia recruits were recorded in polyculture with smaller African sharptooth catfish compared to monoculture of Nile tilapia (p = 0.010). Large African sharptooth catfish predators controlled the number of Nile tilapia recruits to less than ten percent (only 6 % remained) at the end of the experiment. Comparatively, this represents about 12 (80 %) and 2 (13 %) times the number of recruits that remained in monoculture of Nile tilapia and polyculture with smaller African sharptooth catfish, respectively.

The mean number of Nile tilapia recruits in the three experimental treatments at the end of the experiment. Different letters above the bars show significant differences (p ≤ 0.05) and vertical bars indicate standard error (SE) of the mean
At the end of the 126-day rearing period, the body weight classes of tilapia recruits ranged from 0.10 to 30.00 g (Fig. 2). In all situations, with or without African sharptooth catfish predators, very small tilapia recruits (<3 g) were abundant. In the ponds stocked with small catfish, recruits >20 g were rare, whereas in those stocked with large African sharptooth catfish, they were more abundant. These variations in weight classes contributed to the significant difference in the mean body weight of Nile tilapia recruits among the treatments (F 2,387 = 8.470, p < 0.001; Fig. 3). The mean body weight of Nile tilapia recruits was significantly higher in the tanks stocked with larger (14.78 ± 1.39 g) than smaller African sharptooth catfish predator (8.71 ± 0.59 g, p < 0.001) and monoculture of Nile tilapia (11.52 ± 0.70 g; p = 0.050). Nile tilapia recruits stocked in monoculture tanks attained significantly higher mean body weights than those stocked in tanks with smaller African sharptooth catfish predator (p = 0.024).

The number of Nile tilapia recruits at various weight classes obtained at the end of the experiment

The mean weight of Nile tilapia recruits in the three experimental treatments at the end of the experiment. Different letters above the bars show significant difference (p ≤ 0.05) and error bars indicate standard error (SE) of the mean
Growth performance and percentage survival
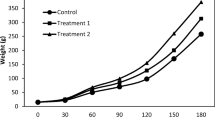
The mean weight increase in Nile tilapia in the three experimental treatments is shown in Fig. 4. The size of predator African sharptooth catfish affected significantly the final weight of Nile tilapia (F 2,42 = 278.283, p < 0.001). The multiple comparisons test showed significantly higher FMW (198.74 ± 0.39 g) of Nile tilapia in larger than smaller African sharptooth catfish predators (140.52 ± 3.27 g) and monoculture of Nile tilapia (128.39 ± 2.10 g) (p < 0.001; Table 2). Similarly, Nile tilapia reared together with smaller African sharptooth catfish revealed significantly higher FMW than those reared in monoculture (p = 0.01; Table 2). Nile tilapia stocked with large African sharptooth catfish had comparatively higher FMW (43 %) than those in polyculture with small African sharptooth catfish (30 %) and monoculture system (27 %).

Mean weight increase of Nile tilapia cultured in monoculture and polyculture with smaller and larger African sharptooth catfish predator. Vertical bars indicate standard error (SE) of the mean
Furthermore, DWG of Nile tilapia was also significantly affected by the size of the predator (F 8,64 = 2.829, p = 0.012). The polyculture of Nile tilapia with larger African sharptooth catfish resulted in significantly higher DWG of Nile tilapia compared with those reared with smaller African sharptooth catfish and Nile tilapia alone (p < 0.001; Table 2). Furthermore, the DWG for Nile tilapia polycultured with small African sharptooth catfish was significantly higher than those reared alone (p < 0.001). Similarly, SGR of Nile tilapia was significantly affected by the size of the predator African sharptooth catfish (F 8,64 = 161.071, p < 0.001). Nile tilapia reared with larger African sharptooth catfish had significantly higher SGR than those raised with small African sharptooth catfish and in monoculture (p < 0.001; Table 2). Likewise, Nile tilapia reared with smaller African sharptooth catfish had a significant SGR than those reared alone in monoculture (p < 0.001; Table 2). Percentage survival of Nile tilapia was not affected in any case by the size of the predator (p > 0.05).
Yield
The size of the predator affected significantly the yield performance of the different treatments (F 2,6 = 8.179, p = 0.019). The NFY and NAY were significantly lower in polyculture with large African sharptooth catfish than in monoculture of Nile tilapia (p = 0.016; Table 3). However, NFY and NAY were not significantly different between monoculture of Nile tilapia and polyculture with small African sharptooth catfish (p = 0.115).
The percentage weight of Nile tilapia recruits reared in monoculture contributed almost 60 % to the yield, while those in polyculture contributed less than 20 % (Table 3). The contribution differed significantly among the culture systems (F 2,6 = 28.663, p < 0.001). Nile tilapia recruits reared in monoculture contributed significantly more to the yield than those in polyculture with smaller (p = 0.001) and larger African sharptooth catfish (p = 0.002).
In the polyculture setup, the 21 stocked adult fishes (Nile tilapia and African sharptooth catfish) contributed over 80 % to the total yield, while in the tilapia monoculture the contribution was only 42 % of the total yield. The contribution of adult Nile tilapia to the yield differed significantly among the experimental setup (F 2,6 = 12.174, p = 0.008). The percentage weight of adult Nile tilapia cultured in polyculture with smaller African sharptooth catfish contributed significantly higher to the yield compared to those cultured in monoculture (p = 0.011) and polyculture with larger African sharptooth catfish (p = 0.014; Table 3). Larger African sharptooth catfish exhibited significantly higher percentage weight contribution to yield than smaller ones (t (4) = −13.270, p < 0.001; Table 3).
Water quality parameters
The water quality parameters (mean ± standard deviation) were stable in all treatments with minor fluctuations. Water temperature ranged from 23.90 to 28.10 °C with mean values between 26.23 ± 1.13 and 26.51 ± 0.96 °C recorded in Nile tilapia alone and in polyculture with small African sharptooth catfish, respectively. Dissolved oxygen ranged from 2.90 to 10.19 mg L−1 with mean values between 5.99 ± 1.65 and 6.24 ± 1.54 mg L−1 obtained in polyculture with small and large African sharptooth catfish, respectively. pH values ranged from 5.59 to 7.20 with a mean of 6.55 ± 0.66 in all treatments. Results of all water quality parameters measured did not show any significant differences among treatments (p > 0.05).
Discussion
The results have shown that, large-sized African sharptooth catfish have a significant effect on reducing the number of recruits in mixed-sex Nile tilapia culture. A similar efficiency of larger compared to smaller size of catfish predation on recruits of Nile tilapia has been reported by Abdel-Tawwab (2005). Moreover, Offem et al. (2009) found highest percentage (71 %) of sub-marketable Nile tilapia recruits in monoculture followed by polyculture stocked with small (49 %) and large (7 %) Vundu catfish (Heterobranchus longifilis) predator.
The efficiency of larger compared to smaller catfish predators on recruitment control of Nile tilapia is related to body size. Body size of a predator has been shown as a critical factor in predator–prey relationships (Fessehaye et al. 2006). Variation in size between them is considered the most significant factor in predation efficiency owing to mouth width limitations (Wysujack and Mehner 2005; Offem et al. 2009), because prey recruits are generally swallowed whole by predators (Fessehaye et al. 2006). In the present study, African sharptooth catfish were able to swallow Nile tilapia recruits depending on initial size of the predator and this ability was two times in larger African sharptooth catfish than the smaller ones. Thus, based on the present experimental design, it is recommended to stock African sharptooth catfish of at least 60 g to control mixed-sex Nile tilapia recruitment in the culture systems depending on initial size of the prey.
The existence of 6 and 13 % in the number of recruits at the end of the experiment from polyculture with large and small African sharptooth catfish, respectively, indicates that the recruitment of Nile tilapia was only partially controlled. These findings are in agreement with those of de Graaf et al. (1996), Abdel-Tawwab (2005) and Fessehaye et al. (2006) who reported failure of large-sized African sharptooth catfish to control completely the recruitment of Nile tilapia. This indicates that other factors such as supplementary feeding in the culture systems also play a role in affecting the predation efficiency. In their study, de Graaf et al. (1996) reported increasing efficiency in controlling Nile tilapia recruitment by African sharptooth catfish in low food systems. Equally, El-Gamel et al. (1998) observed reduced predatory efficiency by African sharptooth catfish in earthen ponds when artificial feed was offered. Moreover, Abdel-Tawwab (2005) using different sizes of African sharptooth catfish (35, 75, 180, 275, 400 and 650 g) indicated that artificial feeding reduced the predatory rate of all African sharptooth catfish weights. Furthermore, Oyelese (2007) reported reduced predation efficiency on Nile tilapia recruits by African sharptooth catfish fed to satiation compared to those fed sparingly. These findings imply that, feeding the fishes in polyculture reduced the ability of African sharptooth catfish to completely control recruitment because of nutritional satisfaction. Consequently, the numbers of recruits at the end of the experiment from polyculture with large and small African sharptooth catfish were 6 and 13 %, respectively.
Polyculture of Nile tilapia with large African sharptooth catfish depicted significantly higher mean weights of the number of recruits remaining at the end of the experiment. These results are corroborated by the findings of Offem et al. (2009) using Vundu catfish. The predatory ability of African sharptooth catfish depends on growth rate of the prey population. Oyelese (2007) showed that, predation of African sharptooth catfish on Nile tilapia recruits stopped after the sixth week owing to the increased body size of the prey. This shows that, there is a critical period of predation by African sharptooth catfish on Nile tilapia recruits which is controlled by growth rate of the prey. As the prey grew bigger, it became less vulnerable to predation by African sharptooth catfish due to development of fin rays which complicate predation (Abdel-Hadi et al. 2011). This allowed a few Nile tilapia recruits available in polyculture with large African sharptooth catfish to utilize the available resources such as space and food to grow faster resulting into significantly bigger size (weight).
The present study demonstrated that, weight gained by Nile tilapia stocked with large African sharptooth catfish was more than 40 % compared to those stocked with small African sharptooth catfish and in monoculture system. Similarly, all the growth parameters were significantly higher in Nile tilapia stocked with large African sharptooth catfish. These results are similar to those obtained by de Graaf et al. (1996) using African sharptooth catfish and African snakehead (Ophiocephalus obscuris), Oyelese (2007) using African sharptooth catfish and Offem et al. (2009) using Vundu catfish and Nile tilapia. Likewise, Rudolf (2006) using spring salamander (Gyrinophilus porphyriticus) as a predator and Southern two-lined Salamander (Eurycea cirrigera) as a prey found an increased growth rate of prey at large size of the predator.
The increased growth performance of Nile tilapia stocked in polyculture with large African sharptooth catfish is due to predation on recruits by the latter. Predation by African sharptooth catfish reduced competition for space and food caused by recruits derived from the initially stocked adults through reproduction (Oyelese 2007). The availability of food and space influenced increased growth performance because of enhanced feeding (Offem et al. 2009). On the contrary, the presence of 80 and 13 % recruits in monoculture and polyculture with smaller African sharptooth catfish, respectively, interfered with the growth performance through feed competition and space between the stocked adult Nile tilapia and their recruits. Consequently, the growth of adult Nile tilapia was restricted in polyculture with small African sharptooth catfish and in monoculture. The restriction in growth of the adult Nile tilapia by recruits is significant because of the higher metabolic rates of young ones (de Silva et al. 1986). The restriction problem was reduced by removing the recruits through stocking large predatory African sharptooth catfish resulting in increased growth performance of the adult Nile tilapia in the present study.
Percentage survival of Nile tilapia was not affected by the size of the predator African sharptooth catfish. Ultimately, the percentage survival was 100 % in all the treatments. This indicates that the experimental conditions in this study were conducive for Nile tilapia survival in all treatments (Offem et al. 2009). The high percentage survival of Nile tilapia was caused by enough food and adequate space for co-existence with African sharptooth catfish. Adult Nile tilapia has shown to have high survival when cultured in monoculture and polyculture with African sharptooth catfish (Lin and Diana 1995; Veverica et al. 2001). The two species can be grown together to improve the growth of Nile tilapia and ensure diversified products at the end of the growth period to the producer for increased economic benefits. This is important because the two species are known to be good source of protein and income (Chakraborty et al. 2011; FAO 2012; Yakubu et al. 2012). Since large African sharptooth catfish provide conducive environment (optimal predation efficiency) for the rapid rearing of adult Nile tilapia for food and income, this technology should be advocated to fish farmers.
Recruitment control by large African sharptooth catfish predator reduced significantly the NFY and NAY in polyculture than monoculture of Nile tilapia. A reduction in yield in polyculture systems has been observed in earlier experiments with other catfish species such as African snakehead (de Graaf et al. 1996). The reduced yields in polyculture with large than small African sharptooth catfish and in monoculture is contributed by the minimized number of Nile tilapia recruits by the large African sharptooth catfish. Similar observation has been reported by Abdel-Tawwab (2005) who found lower contribution of recruits to the total yield in polyculture with large African sharptooth catfish than in monoculture. Recruits contribution to the yield in monoculture was higher (58 %) than in polyculture with large catfish (17 %), suggesting that, the consumed recruits were converted into a lower weight by the predator. The lost fingerlings weight could not be fully compensated by the observed higher growth rates of Nile tilapia in polyculture with large African sharptooth catfish. However, this is not worrisome because the loss in biomass is economically compensated by the higher price obtained after selling the larger Nile tilapia produced (de Graaf et al. 1996).
Conclusion
Large-sized African sharptooth catfish used in the present study is more efficient in reducing the number of recruits in mixed-sex Nile tilapia culture. The reduced number of recruits by the large African sharptooth catfish predator lowers competition for space and food for adult Nile tilapia resulting in increased growth performance. The reduction in number of Nile tilapia recruits in polyculture with large African sharptooth catfish contributes to low yields. Nevertheless, the low yields are compensated by the higher price obtained from selling the larger Nile tilapia produced. This study revealed that, fish farmers can reduce prolific breeding, obtain higher growth performance and produce larger-sized marketable Nile tilapia by predominantly stocking them with large African sharptooth catfish predator with initial average weight of at least 60 g.
References
Abdel-Hadi Y, Craig J, Babaluk J, Wassle R (2011) Oxytetracycline marking studies of tilapia; Oreochromis niloticus. Better science, better fish, better life. In: Liping L, Fitzsimmons K (eds) Proceedings of the sixth international symposium on Tilapia in Aquaculture: AquaFish Collaborative Research Support Program, pp 60–66
Abdel-Tawwab M (2005) Predation efficiency of Nile catfish, Clarias gariepinus (Burchell, 1822) on fry Nile tilapia, Oreochromis niloticus (Linnaeus, 1758): effect of prey density, predator size, feed supplementation and submerged vegetation. Turk J Fish Aquat Sci 5:69–74
Carlberg JM, Olst JVC, Massingi MJ (2000) Hybrid striped bass: an important fish in US aquaculture. Aquac Mag 26:26–38
Chakraborty SB, Mazumdar D, Chatterji U, Banerjee S (2011) Growth of mixed-sex and monosex Nile tilapia in different culture systems. Turk J Fish Aquat Sci 11:131–138
Dagne A, Degefu F, Lakew A (2013) Comparative growth performance of mono-sex and mixed-sex Nile tilapia (Oreochromis niloticus L.) in pond culture system at Sebeta, Ethiopia. Int J Aquacult 3(7):30–34
de Graaf G, Galemoni F, Banzoussi B (1996) Recruitment control of Nile tilapia, Oreochromis niloticus, by the African catfish, Clarias gariepinus (Burchell 1822), and the African snakehead, Ophiocephalus obscuris. I. A biological analysis. Aquaculture 146(1–2):85–100
de Silva CD, Premawansa S, Keembiyahetty CN (1986) Oxygen consumption in Oreochromis niloticus (L.) in relation to development, salinity, temperature and time of day. J Fish Biol 29(2):267–277
El-Gamel AA, Abdel-Halim AMM, Abdel-Razek E, Solomon A (1998) Biological studies on the Nile perch, Lates niloticus (L.) and African catfish, Clarias gariepinus (T.) in reference to their food habits and predation pattern in culture ponds. Egypt J Agric Res 76:335–349
El-Naggar G (2007) Efficiency of African catfish, Clarias gariepinus in controlling unwanted reproduction of Nile Tilapia Oreochromis niloticus in low input production system. Egypt Aqua Biol Fish 11(3):105–113
Fagbenro OA (2004) Predator control of overpopulation in cultured tilapias and the alternative uses for stunted tilapias in Nigeria. In: Bolivar RB, Mair GC, Fitzsimmons K (eds) Proceedings from the sixth international symposium on Tilapia Aquaculture (VI ISTA). Bureau of Fisheries & Aquatic Resources, Manila, pp 634–647
FAO (2012) The state of world fisheries and aquaculture. Food and Agriculture Organization of the United Nations, Rome
Fessehaye Y, Kabir A, Bovenhuis H, Komen H (2006) Prediction of cannibalism in juvenile Oreochromis niloticus based on predator to prey weight ratio, and effects of age and stocking density. Aquaculture 255:314–322
Hernández M, Gasca-Leyva E, Milstein A (2014) Polyculture of mixed-sex and male populations of Nile tilapia (Oreochromis niloticus) with the Mayan cichlid (Cichlasoma urophthalmus). Aquaculture 418–419:26–31
Kaliba AR, Osewe KO, Senkondo EM, Mnembuka BV, Quagrainie KK (2006) Economic analysis of Nile tilapia (Oreochromis niloticus) production in Tanzania. J World Aquacult Soc 37(4):464–473
Lin CK, Diana JS (1995) Co-culture of catfish (Clarias macrocephalus × C. gariepinus) and tilapia (Oreochromis niloticus) in ponds. Aquat Living Resour 8:449–454
Lowe MR, Wu W, Peterson MS, Brown-Peterson NJ, Slack WT, Schofield PJ (2012) Survival, growth and reproduction of non-native Nile tilapia II: Fundamental niche projections and invasion potential in the Northern Gulf of Mexico. PLoS One 7(7):1–10
Mair GC, Little DC (1991) Population control in farmed tilapias. NAGA, International Center for Living Aquatic Resources Management (ICLARM) Quarterly 4:8–9
Mair GC, Van Dam AA (1996) The effect of sex ratio at stocking on growth and recruitment in Nile tilapia (Oreochromls niloticus) ponds. In: Pullin RSV, Lazard J, Legendré M, Kothias Amon JB, Pauly D (eds) Third international symposium on Tilapia in Aquaculture: International Center for Living Aquatic Resources Management (ICLARM)., pp 100–107
Offem BO, Ikpi GU, Ayotunde EO (2009) Effect of stocking size of the predatory African catfish (Heterobranchus longifilis V.) on the growth performance of Nile Tilapia (Oreochromis niloticus L.) in pond culture. Int J Fish Aquacult 1(3):038–043
Ofor CO, Udo UI, Udoidiong MO (2011) Effect of repeated partial cropping on population dynamics and yield of Oreochromis niloticus (L.) during polyculture with Heterobranchus longifilis (Val.). Int J Fish Aquacult 3(7):126–135
Oyelese OA (2007) Prey/predator relationship of Clarias gariepinus on tilapia (Oreochromis niloticus) populations. Res J Biol Sci 2(1):17–24
Rudolf VHW (2006) The influence of size-specific indirect interactions in predator–prey systems. Ecology 87(2):362–371
Shrestha MK, Sharma RL, Gharti K, Diana JS (2011) Polyculture of Sahar (Tor putitora) with mixed-sex Nile tilapia. Aquaculture 319(1–2):284–289
Veverica K, Liti D, Were E, Bowman J (2001) Fish yields and economic benefits of Tilapia/Clarias polyculture in fertilized ponds receiving commercial feeds or pelleted agricultural by-products. In: Gupta A, McElwee K, Burke D, Burright J, Cummings X, Egna H (eds) Eighteenth annual technical report pond dynamics/aquaculture CRSP: Oregon State University., pp 27–29
Winer BJ (1971) Statistical principles in experimental design, 2nd edn. McGraw-Hill, New York
Wysujack K, Mehner T (2005) Can feeding of European catfish prevent cyprinids from reaching a sized refuge? Ecol Freshw Fish 14:87–90
Yakubu AF, Obi A, Okonji VA, Ajiboye OO, Adams TE, Olaji ED, Nwogu NA (2012) Growth performance of Nile tilapia (Oreochromis niloticus) as affected by stocking density and feed types in water flow through system. World J Fish Mar Sci 4(3):320–324
Yi Y, Lin CK, Diana JS (2002) Culture of mixed-sex Nile tilapia with predatory snakehead. Corvallis, Oregon
Zar JH (2010) Biostatistical analysis, 5th edn. Prentice Hall Inc, New Jersey
Acknowledgments
We express our sincere thanks to Tanzania Commission for Science and Technology (COSTECH) through “KILIMO KWANZA” programme for provision of a research grant. Tanzania Fisheries Research Institute (TAFIRI), the University of Dar es Salaam and Sokoine University of Agriculture (SUA) are acknowledged for granting permission for authors to participate in the study. SUA is also acknowledged for provision of research facilities.
Conflict of interest
The authors declare that they have no competing interests.
Authors’ contributions
SML designed the study, collected and analysed data and drafted the manuscript. APS designed the study, collected and analysed data. HAL designed the study, collected and analysed data. EDS designed the study, collected and analysed data. MAK-M designed the study, collected and analysed data. HFM designed the study, collected and analysed data. YDM designed and supervised the study. All authors read and approved the final manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Limbu, S.M., Shoko, A.P., Lamtane, H.A. et al. Effect of initial stocking size of the predatory African sharptooth catfish (Clarias gariepinus) on recruits, growth performance, survival and yield of mixed-sex Nile tilapia (Oreochromis niloticus) in concrete tank culture system. Int Aquat Res 7, 63–73 (2015). https://doi.org/10.1007/s40071-014-0093-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40071-014-0093-3