
Abstract
Key message
Recruitment strategies of Mexican beech seedlings depend on mother tree distribution and light incidence in early stages of development. Spatial patterns are also affected by the structure and composition of tree species within Mexican beech forests.
Context
Canopy openness and conspecific trees play a key role in Mexican beech spatial distribution and might strongly influence ecosystem functioning in the Tropical Montane Cloud Forest. The observed relationship between diameter at breast height of conspecific trees and the spatial distribution of beech seedlings indicates that structure and composition of tree species are crucial for the establishment and survival of seedlings, providing protection during the first developmental stages.
Aims
To describe the spatial patterns of beech seedlings’ distribution after a masting event and to evaluate the association of these patterns with canopy openness, conspecific tree distribution, and occurrence of other canopy species.
Methods
We sampled individual Mexican beech seedlings on two highly conserved beech forests. We selected 100 subplots (10 × 10 m) on each one to measure seedlings and spatial attributes. We counted the number of beech seedlings and number of mature trees species on each subplot. In addition, we measured the levels of canopy openness and the diameter at breast height for all mature trees in each subplot. Spatial pattern of beech seedlings and their association with adult trees and other species were examined.
Results
Mexican beech seedlings showed significant pattern of spatial aggregation. Significant associations were found between beech seedlings, canopy openness, and beech tree adults, while significant disassociations exist among beech seedlings and other species of trees such as Quercus meavei, Q. delgadoana, Q. trinitatis, and Magnolia schiedeana.
Conclusion
The presence of oak species and Magnolia schiedeana in the surroundings and the structure and composition within forests may play a key role in the maintenance of the specific micro-environmental conditions required by Mexican beech recruitment after a masting event.
Similar content being viewed by others

1 Introduction
Spatial patterns during seedlings’ recruitment are an important factor influencing the dynamics of forest communities (Kunstler et al. 2004; Nathan and Muller-Landau 2000). Forest regeneration is the result of sexual and asexual reproduction, seed dispersal, and recruitment such as seed germination and seedling survival, which are a crucial stage in the regeneration of beech forests (Beaudet and Messier 2008; Kunstler et al. 2004; E Silva et al. 2012). Seed dispersal is strongly dependent on the spatial disposition of mature trees, which protect seedlings from direct sunlight during the first developmental stages and causes one-sided competition for light (Alvarez-Aquino and Williams-Linera 2002; Camacho-Cruz et al. 2000; Queenborough et al. 2007; Wang et al. 2012). The sheltering effect, mostly associated with micro-environmental factors (i.e., canopy openness, pH, moisture, leaf litter), probably has a main effect on the structure and floristic composition of forests ecosystems (Leibold and McPeek 2006; Messaoud and Houle 2006; Peña et al. 2011; Tuomisto et al. 2012). Specifically, the type of spatial distribution shown by seedlings and adult trees (i.e., regular, aggregated or random) has been used to determine the dispersal capabilities and recruitment probabilities of plants (Camarero et al. 2005; Dusan et al. 2007; Meier et al. 2010; Snell et al. 2014; Szymura et al. 2015).
In septentrional beeches, root suckers play a key role increasing competitiveness and survival under adverse conditions (Beaudet and Messier 2008). Peters (1992) registered asexual reproduction by multi-stemmed plants in northern beech individuals. Asexual reproduction is more important and common than sexual reproduction in northern counterparts. The regeneration ecology of Mexican beech is different to northern species from the same genus, because the vegetation type where it grows in Mexico, the Tropical Montane Cloud Forest, is a favorable environment for shade-tolerant tree species.
In this study, we analyzed the spatial patterns of Mexican beech seedlings (Fagus grandifolia subsp. mexicana (Martínez) A.E. Murray), a Miocene relict species endemic to the Tropical Montane Cloud Forest in eastern Mexico. This forest community covers approximately 144.54 ha through a series of fragmented patches at altitudes of 1400–2000 m in the states of Hidalgo, Nuevo León, Puebla, Tamaulipas, and Veracruz (Montiel-Oscura et al. 2013; Peters 1992; Rodríguez-Ramírez et al. 2013; Rowden et al. 2004). Mexican beech is considered an endangered species under Mexican law (González-Espinosa et al. 2011; SEMARNAT 2010) and is currently threatened by human activities, such as illegal logging and deforestation (Téllez-Valdés et al. 2006; Williams-Linera et al. 2003).
The beechnuts of Mexican beech are shade-tolerant and can survive under low light incidence (Godínez-Ibarra et al. 2007). These beechnuts fall from the mature trees mainly on August, the more humid month of the year. They quickly respond to humidity when they fall to the ground. Seemingly, these beechnuts of this species have low dormancy (approximately 1 month sensu Peters (1992)) and personal observations; however, they respond to canopy openness by accelerating their development (Gazol and Ibáñez 2010; Pedraza-Pérez and Williams-Linera 2005). Development and growth of seedlings may also be affected by canopy openness, inter-specific competition (or cooperation sensu Kiers et al. (2011)), and accumulation of leaf litter (Collet and Chenost 2006; Jennings et al. 1999). Moreover, Mexican beech is characterized by the occurrence of synchronous, inter-annual variation in reproductive effort, commonly known as “masting events” at 2- to 8-year intervals (Ehnis 1981; Godínez-Ibarra et al. 2007; Pérez-Rodríguez 1999; Rodríguez-Ramírez et al. 2013). Unfortunately, the adaptive significance and evolutionary precursors of masting events have not yet been fully determined (Kelly 1994; Kelly and Sork 2002). The resource-matching hypothesis provides the most parsimonious explanation for masting (Monks and Kelly 2006; Sork et al. 1993). This hypothesis states that plants allocate a constant fraction of assimilated carbon for reproduction each year. Thus, highly favorable climatic conditions in specific years promote high rates of carbon assimilation and thus a greater beechnut production (Piovensan and Adams 2001, 2005).
In this context, the structure and floristic composition of forest ecosystems are intimately related to micro-environmental factors such as proportion of mature healthy trees, canopy openness, soil type, pH, and moisture (Taugourdeau and Sabatier 2010; Peña et al. 2011; Rodríguez-Ramírez et al. 2016). The spatial distribution of seedlings is strongly correlated by the random opening and closure of the canopy, which produces micro-habitat heterogeneity (Meier et al. 2010; Offord et al. 2014; Barna and Bosela 2015). Thus, the study of the spatial pattern of seedlings can aid in the understanding of the biotic and abiotic requirements for the establishment and survival of currently endangered forest species, such as Mexican beech (Godínez-Ibarra et al. 2007; Övergaard et al. 2007; Hardiman et al. 2011).
Mexican beechnuts are dispersed randomly near the mother trees in a segregate pattern (Peters 1995, 1997). After the masting event, germination is extensive and the produced seedlings compete for light and water (Godínez-Ibarra et al. 2007). Moreover, canopy openness is an essential factor for beech seedling development, notwithstanding the shade-tolerant natural of beech (E Silva et al. 2012). The ecology of beech seedlings has been extensively studied in northern beeches, but few studies focusing on the spatial distribution at local level have been done (i.e., Gazol and Ibáñez (2010), Godínez-Ibarra et al. (2007), Kunstler et al. (2004) and Shibata et al. (2002)).
In this study, we describe for the first time the spatial patterns of Mexican beech seedlings within a beech-dominated Tropical Montane Cloud Forest in the state of Hidalgo, Mexico. Our aims were (1) to describe the spatial patterns of beech seedlings distribution after a masting event and (2) to evaluate the association of these patterns with canopy openness, conspecific tree distribution, and occurrence of other canopy species. With this context, the present analysis provides a deeper understanding of the auto-ecological processes that affect the establishment and survival of Mexican beech in the Tropical Montane Cloud Forest.
2 Materials and methods
2.1 Study sites
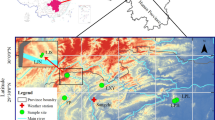
The study sites are located within two extensive and highly preserved forest stands of Mexican beech in the state of Hidalgo: La Mojonera, Zacualtipán de Ángeles (20° 38′ N, 98° 36′ W; 1780–1950 m a.s.l.), and Medio Monte, San Bartolo Tutotepec (20° 24′ N, 98° 14′ W; 1800–1944 m a.s.l.; Fig. 1(A), Table 1; Rodríguez-Ramírez et al. 2013). These sites have a temperate Cb climate (sensu García (1988)) characterized by mild temperatures, dry cool season from November to January, dry warm season from early February to May, cool summer (June to July), and wet cool season from August to October (Peters 1995; Williams-Linera 2007). The high frequency of fog and precipitation causes persistent moisture (60–85%) throughout the year (García 1988; Tinoco-Rueda 2009). The dominant lithologies are vitric (Tv) and humic (Th) andosols (FAO-UNESCO; Batjes 1997) with a slimy sandy-clay loam texture and acidic pH values ranging between 4 and 6 (Peters 1995, 1997).

(A) Geographical location of the two study sites of Fagus grandifolia subsp. mexicana (Martínez) A.E. Murray (Mexican beech) in the Tropical Montane Cloud Forests of the Sierra Madre Oriental, Mexico. (B) Schematic representation of the 100 × 100 m plots subdivided into 10 × 10 m subplots
La Mojonera has one of the most extensive Mexican beech forests in Mexico (42.5 ha); it is the most studied and shows the least disturbance by human activities (Rowden et al. 2004; Rodríguez-Ramírez et al. 2013). In addition to Mexican beech (Fagus grandifolia subsp. mexicana), the high-canopy (> 20 m) of the forest is composed of Pinus patula Schltdl. & Cham., Quercus trinitatis Trel., Liquidambar styraciflua L., and Magnolia schiedeana Schltdl. The mid-canopy (10–20 m) of the forest is dominated by Clethra mexicana DC., Befaria aestuans L., and Quercus delgadoana S. Valencia, Nixon & L. M. Kelly.
The Mexican beech forest at Medio Monte (34.25 ha) is located on slightly rugged terrains and is characterized by trees reaching up to 25 m in height. The forest is dominated by F. grandifolia subsp. mexicana, Magnolia schiedeana, and Quercus meavei Valencia-A., Sabás & Soto. The mid-canopy of the forest is mainly composed of Clethra mexicana, Q. delgadoana, and Q. trinitatis, whereas the low canopy consists mainly of Cyathea fulva (M. Martens & Galeotti) Fée and Dicksonia sellowiana Hook. (Table S1). A more detailed description of plant species composition and structure in the studied forests can be found in Rodríguez-Ramírez et al. (2016).
2.2 Sampling protocol
Individual beech seedlings were counted (December 2012 to January 2013) shortly after the occurrence of a masting event (August–September 2012). A permanent plot of 100 × 100 m2 was set up in representative areas of forest stands, as far as possible from human settlements and roads. In each permanent plot, we defined 100 subplots of 10 × 10 m (Fig. 1(B); Roberts-Pichette and Gillespie 1999). We used a manual counter (Base Mount Tally Counter) in each subplot to quantify each beech seedling with a height ≤ 10 cm (Peters 1995). In each subplot, all mature trees were measured following Kindt and Coe (2005) method; these authors considered that individual trees reach maturity when they achieve (1) a diameter at breast height ≥ 15 cm, and (2) ≥ 30 m high. All the trees were identified to the species level and compared with herbaria specimens from HGOM (Universidad Autónoma del Estado de Hidalgo) and the Herbario del Instituto de Ecología, A.C., México (IE-XAL).
In order to evaluate the influence of canopy openness on the distribution of Mexican beech seedlings, we measured canopy openness for each subplot using a canopy densitometer (Model A, Lemmon 1956; Fiala et al. 2006) between 9:00 am and 12:00 pm to standardize the daily sunlight fluctuation. In each subplot, we performed five independent measurements of canopy openness in four different orientations (i.e., north, south, east, and west; Fig. 1(C)). The data obtained through the measurements were averaged for statistical analyses.
2.3 Spatial analyses of beech seedlings, canopy openness, and conspecific trees
We described the spatial pattern of beech seedling distribution and evaluated its association with the diameter at breast height of mature trees of different species and canopy openness using the software SADIE (Spatial Analysis by Distance IndicEs; Perry 1998; Perry and Dixon 2002; Perry et al. 2006). SADIE incorporates spatial information of the counted data from geographically referenced points (Perry et al. 1999), allowing to identify the exact location of seedling hotspots.
This method is based on “distance regularity” (D), which measures the total distance involved in passing from the observed spatial arrangement of counts to move to a regular arrangement where all units have the same value of the variable under study. To calculate D, which is characterized by a high proportion of zeros and a non-normal distribution, SADIE uses the transportation algorithm from the linear programming literature. One main advantage of this method is that the input data does not need to present a normal distribution, show isotropism (i.e., no environmental changes), or be regularly spaced (Maestre and Cortina 2002; Maestre et al. 2003). Thus, analyses using SADIE are not limited by these restrictions and the results are conditioned on the levels of heterogeneity present in the data (Bell 1998; Perry et al. 1996; Perry et al. 1999).
SADIE evaluates the magnitude and significance of D (distance to regularity) by comparing the observed D value with those obtained from randomized datasets. In the present case, the observed distribution of beech seedlings across subplots was compared to 1000 randomly generated distributions (i.e., random number of seedlings per subplot). SADIE provides two complementary measures of the spatial structure of count data. First, the index of aggregation, Ia, is a global measure of the degree of aggregation or a variable, which was calculated for each data set to determine overall 23,433 randomizations each, where Ia = 1 indicates random arrangement of counts of no significant spatial pattern; Ia > 1, aggregated arrangement giving clusters of observed counts; and Ia < 1, regular arrangement of counts (Perry and Dixon 2002). The probability Pa tests for deviations from random dispersal where Pa > 0.975 indicates regular dispersal; Pa < 0.025, spatial aggregation; and 0.025 < Pa < 0.975, randomness (Perry 1998; Perry and Dixon 2002). Second, the clustering indices (v) are a parameter calculated for each point in the data base which were estimated for subplots with Mexican beech seedling densities above (v i > 1.5) and below the plot’s average (v j < − 1.5) (Perry et al. 1999; Winder et al. 2001; Maestre and Cortina 2002; Perry and Dixon 2002) where subscripts i and j indicate patches and gaps, respectively. Mean clustering indices (\( \overline{vi\ } \) and \( \overline{vj\ } \)) were also calculated for each data set and compared with their corresponding values from the 23,433 randomizations to test the overall clustering at 5% level.
2.4 Evaluating spatial association
The local spatial association was measured using an index X k based on similarities between clustering indices of the variables (i.e., beech seedling vs. canopy openness). Positive values of X k (> 0 indicates a positive spatial association) were indicated by a coincidence of two patches or two gaps while negative association (values < 0 indicate a lack of concordance between variables) resulted from a patch coinciding with a gap in both subplots (Perry 1998). We estimated a general X index from the mean values for each subplot in the two study sites, which summarizes the spatial pattern of beech seedling distribution in relation to mature trees and canopy openness (sensu Winder et al. (2001), Perry and Dixon (2002), and Perry et al. (1999)). The significance of the general X index was evaluated through 1,000 randomizations of clustering indices (v) (Dutilleul 1993) for seedlings, adult trees, and light incidence. For this comparison, P values < 0.025 indicated a significant spatial association, whereas P values > 0.975 were indicative of a significant spatial disassociation.
Finally, we visualized the observed patterns of spatial aggregation and association for beech seedlings across the two study sites using Surfer® v. 14 (Golden Software Inc., CO, USA) with the red-blue method (Perry et al. 1996; Perry and Dixon 2002).
3 Results
3.1 Spatial patterns of Mexican beech seedlings
Contour maps of the index of clustering visually indicate the spatial distribution of the main patches and gaps within the sampled plots. In the two studied sites, Mexican beech seedlings showed significant pattern of spatial aggregation (Fig. 2(1A)—La Mojonera: Ia > 2, Pa 0.0002; Fig. 2(2A)—Medio Monte: Ia > 1, Pa 0.063; Table 2). Overall, we also observed significant spatial association in the distribution of Fagus’ seedlings (Ia > 1; Pa 0.0002) and canopy openness (Ia > 1; Pa < 0.0184; Fig. 2).

Maps of interpolated clustering and spatial association of Mexican beech seedling (A), canopy openness (B), and average diameter at breast height for different tree species: beeches (C) and other tree species (D), in two Mexican beech forests (1 = La Mojonera and 2 = Medio Monte). Association is represented by two letters (i.e., A+B). Only significant indices were mapped: v i > 1.5 (patches) and v j < − 1.5 (gaps) for aggregation; X k > 0.5 (positive aggregation) or X k < 0.5 (disassociation)
3.2 Association of Mexican beech seedlings
The spatial distribution of beech seedlings showed a significant association with below-canopy light incidence, with a value ≥ 80% but lower than 100% (P < 0.025; Fig. 2(1), (2A+B)). We found significant associations between the spatial distribution of beech seedlings and the average diameter at breast height of conspecific trees in the two Mexican beech stands (Fig. 2(1A+B) and (2A+B); Table 2). However, we observed a negative correlation between the number of beech seedlings per subplot and the diameter at breast height of mature beeches in La Mojonera: r = − 0.08, P = 0.05; Fig. 2(1A+C). However, there was no evidence of a spatial association between beech seedlings and adult beeches in Medio Monte (X k = 0.016; P = 0.446; Fig. 2(2A+C)). We also found a spatial disassociation between the distribution of beech seedlings and the occurrence of canopy tree species other than Fagus. More specifically, at La Mojonera, we observed a spatial disassociation between beech seedlings and the diameter at breast height of trees excluding Fagus (X k = − 0.3; P = 0.997; Fig. 2(1A+D)). For Medio Monte, we also found a spatial disassociation between beech seedlings and mature trees of other species (X = − 0.3; P = 0.999; Fig. 2(2A+D).
The spatial patterns of association do not include cases of non-significant associations or disassociations. It also shows the distribution of significant clustering values (v i ) (> 1.5 = patches; < − 1.5 = gaps), and values obtained for the association index (X k ) (> 0.5 = association; < − 0.5 disassociation).
4 Discussion
This is the first work that study spatial seedling patterns after a masting event in Mexican beech forest. The use of SADIE allowed us to detect spatial patterns of Mexican beech seedlings at specific locations and determine its association/disassociation with light availability and the presence of adult trees within the forest. The spatial indices obtained with SADIE showed that the distribution of adult beech trees favors the establishment and development of beech seedlings. We suggest that beech mother trees offer sheltering effect to the seedlings and saplings as suggested by Gorzelak et al. (2015). The Janzen-Connell hypothesis explains the maintenance of tropical tree diversity through the interacting effects of parent-centered dispersal patterns and distance- and density-dependent seed survival (Connell et al. 1984). When a gap is opened by fall of a conspecific tree, the opportunity emerges for the sapling to develop. Notwithstanding, the availability of suitable habitat for beech seedlings appears to be affected by the degree of canopy openness and the presence of several oak species (i.e., Quercus delgadoana, Q. trinitatis, and Q. meavei) and Magnolia schiedeana (Fig. 2(1A+D) and (2A+D)).
The possibility of changing spatial patterns (i.e., canopy openness) and its effects on the distribution of beech seedlings and saplings within forest stands requires further attention. Additional specific studies about beech seedlings, micro-environmental characteristics (i.e., pH, moisture, ectomycorrhizal, amount litter, stress, and allelopathy), adaptation of leaf morphology, and asymmetric competition for light resources are needed to understand recruitment and to clarify the influence of these features for the maintenance and dynamics of Mexican beech forests.
The distribution of seedlings may be a direct consequence of beechnut dispersal, which tends to be disseminated relatively close to the mother tree. Under the hypothesis, recruitment only occurs under a small “population recruitment curve” (Janzen 1970) that is centered at some distance from mother trees; this is possibly reflecting in the Mexican beech forests. Likewise, more field studies on the behavior of predatory animals (i.e., mammals, birds, snails, and insects) in relation to the spatial dispersal of beechnuts and Mexican beech seedlings are necessary.
Rodríguez-Ramírez et al. (2016) considered that Mexican beech is a species specialized in micro-environmental conditions within the Tropical Montane Cloud Forest. Besides, Akaji et al. (2016), Alvarez-Aquino et al. (2004), and Camacho-Cruz et al. (2000) showed that the natural recruitment and survival of beech seedlings were higher within structurally mature forests than in more disturbed forests. Thus, despite the high mortality of beech seedlings and presence of conspecific trees, the high production of beechnuts during masting events appears to be enough for the maintenance of Mexican beech populations (Godínez-Ibarra et al. 2007; Cleavitt et al. 2008; Meyer and Pendleton 2015).
The observed relationship between diameter at breast height of mature trees and the spatial distribution of beech seedlings hotspots indicates that the presence of similar structure and floristic composition between Mexican beech stands studied (Rodríguez-Ramírez et al. 2016) is crucial for the establishment and survival of juveniles, providing protection during the first developmental stages. Similarly, Čater and Kobler (2017), Collin et al. (2017), and Ohkubo et al. (1996) observed that the spatial distribution of individuals of Fagus grandifolia Ehrh., F. sylvatica L., and F. japonica Maxim. was positively associated with low levels of canopy openness.
We observed a similar association between the distribution of beech seedlings, canopy openness, and the conspecific trees within the two study sites. This suggests that the spatial distribution of beech seedlings is tightly linked to microsites with specific levels of light availability (Nelson and Wagner 2014; Peterson et al. 1990). In general, Fagus-dominated forests throughout the world have dense canopies (≈ 90%; Ishida et al. 2015; Yan et al. 2015), which favors the establishment and growth of beech seedlings. Several studies on the distribution and establishment of seedlings have shown similar results to ours by analyzing other beech species, namely Fagus sylvatica in southeastern Europe (Čater and Kobler 2017; Dusan et al. 2007), F. crenata Blume and F. japonica in Japan (Ariya et al. 2015; Ohkubo et al. 1996; Tomita et al. 2002), and F. engleriana Seemen in central China (Zhang et al. 2007). Similar patterns have also been observed in other tree species sharing environmental conditions with Mexican beech, such as Magnolia schiedeana and M. dealbata Zucc. in the Tropical Montane Cloud Forest of eastern Mexico (Dieringer and Espinosa 1994; Rivers et al. 2016; Vovides and Iglesias 1996). These shared patterns reinforce the importance of low levels of light availability to the germination and survival of non-light demanding tree species.
The present study advances our understanding of the processes involved in the natural recruitment of this highly endangered species with a relict distribution. However, the presence of oak species (i.e., Quercus delgadoana, Q. trinitatis, and Q. meavei) and Magnolia schiedeana in the surroundings and the differences in the tree structure and composition between forests determine light level and soil conditions in the understory. (Gazol and Ibáñez 2010; Nilsson 1985; Peña et al. 2011; Kitabatake and Wada 2001; Yuan et al. 2011).
Spatial distribution patterns of plant species in Tropical Montane Cloud Forest may also be influenced by niche assembly and random dispersal assembly processes acting at both local and landscape scales (Peña et al. 2011). The establishment of beech seedlings appears to be mainly influenced by conspecific tree distribution, subsequently is also affected by the presence of another tree species that are co-dominant in the Mexican beech forest.
References
Akaji Y, Miyazaki Y, Hirobe M, Makimoto T, Sakamoto K (2016) The relationship between seedling survival rates and their genetic relatedness to neighboring conspecific adults. Plant Ecol 217:465–470. https://doi.org/10.1007/s11258-016-0591-z
Alvarez-Aquino C, Williams-Linera G (2002) Seedling bank dynamics of Fagus grandifolia var. mexicana before and after a mast year in a Mexican cloud forest. J Veg Sci 13:179–184. https://doi.org/10.1111/j.1654-1103.2002.tb02037.x
Alvarez-Aquino C, Williams-Linera G, Newton AC (2004) Experimental native tree seedling establishment for the restoration of a Mexican cloud forest. Restor Ecol 12:412–418. https://doi.org/10.1111/j.1061-2971.2004.00398.x
Ariya U, Hamano KY, Makimoto T, Kinoshita S, Akaji Y, Miyazaki Y, Hirobe M, Sakamoto K (2015) Temporal and spatial dynamics of an old-growth beech forest in western Japan. J For Res 21:73–83. https://doi.org/10.1007/s10310-015-0514-1
Barna M, Bosela M (2015) Tree species diversity change in natural regeneration of a beech forest under different management. For Ecol Manag 342:93–102. https://doi.org/10.1016/j.foreco.2015.01.017
Batjes NH (1997) A world dataset of derived soil properties by FAO/UNESCO soil unit for global modelling. Soil Use Manag 13:9–16. https://doi.org/10.1111/j.1475-2743.1997.tb00550.x
Beaudet M, Messier C (2008) Beech regeneration of seed and root sucker origin: a comparison morphology, growth, survival, and response to defoliation. For Ecol Manag 255:3659–3666. https://doi.org/10.1016/j.foreco.2008.03.010
Bell ED (1998) Spatio-temporal dynamics of UK moths. Ph. D. thesis, University of Leicester, UK
Camacho-Cruz A, González-Espinosa M, Wolf JHD, De Jong BHJ (2000) Germination and survival of tree species in disturbed forests of the highlands of Chiapas, Mexico. Can J Bot 78:1309–1318. https://doi.org/10.1139/b00-103
Camarero J, Gutiérrez E, Fortin MJ, Ribbens E (2005) Spatial patterns of tree recruitment in a relict population of Pinus uncinata: forest expansion through stratified diffusion. J Biogeogr 32:1979–1992. https://doi.org/10.1111/j.1365-2699.2005.01333.x
Čater M, Kobler A (2017) Light response of Fagus sylvatica L. and Abies alba Mill. in different categories of forest edge—vertical abundance in two silvicultural systems. For Ecol Manag 391:417–426. https://doi.org/10.1016/j.foreco.2017.02.027
Cleavitt NL, Fairbairn M, Fahey TJ (2008) Growth and survivorship of American beech (Fagus grandifolia Ehrh.) seedlings in a northern hardwood forest following a mast event. J Torrey Bot Soc 135:328–345. https://doi.org/10.3159/08-RA-025.1
Collet C, Chenost C (2006) Using competition and light estimates to predict diameter and height growth of naturally regenerated beech seedlings growing under changing canopy conditions. Forestry 79:489–502. https://doi.org/10.1093/forestry/cpl033
Collin A, Messier C, Kembel SW, Bélanger N (2017) Low light availability associated with American beech is the main factor for reduced sugar maple seedling survival and growth rates in a hardwood forest of southern Quebec. Forests 8:1–13. https://doi.org/10.3390/f8110413
Connell JH, Tracey JG, Webb LJ (1984) Compensatory recruitment, growth, and mortality as factors maintaining rain-forest tree diversity. Ecol Monogr 54:141–164. https://doi.org/10.2307/1942659
Dieringer G, Espinosa SJE (1994) Reproductive ecology of Magnolia schiedeana (Magnoliaceae), a threatened cloud forest tree species in Veracruz, Mexico. J Torrey Bot Soc 121:154–159. https://doi.org/10.2307/2997167
Dusan R, Stjepan M, Igor A, Jurij D (2007) Gap regeneration patterns in relationship to light heterogeneity in two old-growth beech-fir forest reserves in South East Europe. Forestry 80:431–443. https://doi.org/10.1093/forestry/cpm037
Dutilleul P (1993) Modifying the t-test for assessing correlation between two spatial processes. Biometrics 49:305–314
E Silva D, Rezende-Mazzella P, Legay M, Corcket E, Dupouey JL (2012) Does natural regeneration determine the limit of European beech distribution under climatic stress? For Ecol Manag 266:262–272. https://doi.org/10.1016/j.foreco.2011.11.031
Ehnis DE (1981) Fagus mexicana Martínez: su ecología e importancia. B.Sc. thesis, Facultad de Ciencias, Universidad Nacional Autónoma de México. Mexico City
Fiala ACS, Garman SL, Gray AN (2006) Comparison of five canopy cover estimation techniques in the western Oregon Cascades. For Ecol Manag 232:188–197. https://doi.org/10.1016/j.foreco.2006.05.069
García E (1988) Modificaciones al Sistema de Clasificación Climática de Köppen, 5th edn. Instituto de Geografía. Universidad Nacional Autónoma de México, Mexico City
Gazol A, Ibáñez R (2010) Scale-specific determinants of a mixed beech and oak seedling-sapling bank under different environmental and biotic conditions. Plant Ecol 211:37–48. https://doi.org/10.1007/s11258-010-9770-5
Godínez-Ibarra O, Ángeles-Pérez G, López-Mata L, García-Moya E, Valdez-Hernández JI, Santos-Posadas HD, Trinidad-Santos A (2007) Lluvia de semillas y emergencia de plántulas de Fagus grandifolia subsp. mexicana en La Mojonera, Hidalgo, México. Rev Mex Biodivers 78:117–128
González-Espinosa M, Meave JA, Lorea-Hernández FG, Ibarra-Manríquez G, Newton AC (2011) The red list of Mexican cloud forest trees. Fauna & Flora International Cambridge, UK
Gorzelak MA, Asay AK, Pickles BJ, Simard SW (2015) Inter-plant communication through mycorrhizal networks mediates complex adaptive behavior in plant communities. AoB Plants 7:1–13. https://doi.org/10.1093/aobpla/plv050
Hardiman BS, Bohrer G, Gough CM, Vogel CS, Curtis PS (2011) The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest. Ecology 92:1818–1827. https://doi.org/10.1890/10-2192.1
Ishida B, Masaki T, Miyamoto A, Tanaka H, Nakashizuka T (2015) Projection of the probability of local extinction of canopy tree species in forest landscapes. J For Res 20:337–346. https://doi.org/10.1007/s10310-015-0483-4
Janzen DH (1970) Herbivores and the number of tree species in tropical forests. Am Nat 104:501–528. https://doi.org/10.1086/282687
Jennings SB, Brown ND, Sheil D (1999) Assessing forest canopies and understorey illumination: canopy closure, canopy cover and other measures. Forestry 72:59–74. https://doi.org/10.1093/forestry/72.1.59
Kelly D (1994) The evolutionary ecology of mast seeding. Trends Ecol Evol 9:465–470. https://doi.org/10.1016/0169-5347(94)90310-7
Kelly D, Sork VL (2002) Mast seeding in perennial plants: why, how, where? Annu Rev Ecol Evol Syst 33:427–447. https://doi.org/10.1146/annurev.ecolsys.33.020602.095433
Kiers ET, Duhamel M, Beesetty Y, Mesah JA, Franken O, Verbruggen E, Fellbaum CR, Kowalchuk GA, Hart MM, Bago A, Palmer TM, West SA, Vandenkoornhuyse P, Jansa J, Bücking H (2011) Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 333:880–882. https://doi.org/10.1126/science.1208473
Kindt R, Coe R (2005) Tree diversity analysis. A manual and software for common statistical methods for ecological and biodiversity studies. Nairobi: World Agroforestry Centre (ICRAF)
Kitabatake T, Wada N (2001) Notes on beech (Fagus crenata Blume) seed and seedling mortality due to rodent herbivory in a northernmost beech forest, Utasai, Hokkaido. J For Res 6:111–115. https://doi.org/10.1007/BF02762496
Kunstler G, Curt T, Lepart J (2004) Spatial pattern of beech (Fagus sylvatica L.) and oak (Quercus pubescens Mill.) seedlings in natural pine (Pinus sylvestris L.) woodlands. Eur J For Res 123:331–337. https://doi.org/10.1007/s10342-004-0048-0
Leibold MA, McPeek MA (2006) Coexistence of the niche and neutral perspectives in community ecology. Ecology 87:1399–1410. https://doi.org/10.1890/0012-9658(2006)87[1399:COTNAN]2.0.CO;2
Lemmon PE (1956) A spherical densiometer for estimating forest overstory density. For Sci 2:314–320. https://doi.org/10.1371/journal.pone.0089437
Maestre FT, Cortina J (2002) Spatial patterns of surface soil properties and vegetation in a Mediterranean semi-arid steppe. Plant Soil 241:279–291. https://doi.org/10.1023/A:1016172308462
Maestre FT, Cortina J, Bautista S, Bellot J, Vallejo R (2003) Small-scale environmental heterogeneity and spatiotemporal dynamics of seedling establishment in a semiarid degraded ecosystem. Ecosystems 6:630–643. https://doi.org/10.1007/s10021-002-0222-5
Meier ES, Kienast F, Pearman PB, Svenning JC, Thuiller W, Araújo MB, Guisan A, Zimmermann NE (2010) Biotic and abiotic variables show little redundancy in explaining tree species distributions. Ecography 33:1038–1048. https://doi.org/10.1111/j.1600-0587.2010.06229.x
Messaoud Y, Houle G (2006) Spatial patterns of tree seedling establishment and their relationship to environmental variables in a cold-temperate deciduous forest of eastern North America. Plant Ecol 185:319–331. https://doi.org/10.1111/j.1600-0587.2010.06229.x
Meyer SE, Pendleton BK (2015) Seedling establishment in a masting desert shrub parallels the pattern for forest trees. Acta Oecol 65–66:1–10. https://doi.org/10.1016/j.actao.2015.03.001
Monks A, Kelly D (2006) Testing the resource-matching hypothesis in the mast seeding tree Nothofagus truncata (Fagaceae). Austral Ecol 31:366–375. https://doi.org/10.1111/j.1442-9993.2006.01565.x
Montiel-Oscura D, Ramírez-Herrera C, Ángeles-Pérez G, López-Upton J, Antonio-López P (2013) Allozyme variation and population size of haya mexicana (Fagus grandifolia subsp. mexicana) in the Sierra Madre Oriental. Rev Fitotec Mex 36:413–420
Nathan R, Muller-Landau HC (2000) Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol Evol 15:278–285. https://doi.org/10.1016/S0169-5347(00)01874-7
Nelson AS, Wagner RG (2014) Spatial coexistence of American beech and sugar maple regeneration in post-harvest northern hardwood forests. Ann For Sci 71:781–789. https://doi.org/10.1007/s13595-014-0376-2
Nilsson SG (1985) Ecological and evolutionary interactions between reproduction of beech Fagus sylvatica and seed eating animals. Oikos 44:157–164. https://doi.org/10.2307/3544057
Offord CA, Meagher PF, Zimmer HC (2014) Growing up or growing out? How soil pH and light affect seedling growth of a relict rainforest tree. AoB Plants 6:1–9. https://doi.org/10.1093/aobpla/plu011
Ohkubo T, Tanimoto T, Peters R (1996) Response of Japanese beech (Fagus japonica Maxim.) sprouts to canopy gaps. Vegetatio 124:1–8. https://doi.org/10.1007/BF00045139
Övergaard R, Gemmel P, Karlsson M (2007) Effects of weather conditions on mast year frequency in beech (Fagus sylvatica L.) in Sweden. Forestry 80:555–565. https://doi.org/10.1093/forestry/cpm020
Pedraza-Pérez RA, Williams-Linera G (2005) Condiciones de microhábitat para la germinación y establecimiento de dos especies de árboles del bosque mesófilo de montaña en México. Agrociencia 39:457–467
Peña L, Amezaga I, Onaindia M (2011) At which spatial scale are plant species composition and diversity affected in beech forests? Ann For Sci 68:1351–1362. https://doi.org/10.1007/s13595-011-0147-2
Pérez-Rodríguez PM (1999) Las hayas de México, monografía de Fagus grandifolia spp. mexicana, 1st edn. Universidad Autónoma Chapingo, Montecillo, Estado de México, Mexico
Perry JN (1998) Measures of spatial pattern for counts. Ecology 79:1008–1017. https://doi.org/10.1890/0012-9658(1998)079[1008:MOSPFC]2.0.CO;2
Perry JN, Dixon PM (2002) A new method to measure spatial association for ecological count data. Ecoscience 9:133–141. https://doi.org/10.1080/11956860.2002.11682699
Perry JN, Bell ED, Smith RH, Woiwod IP (1996) SADIE: software to measure and model spatial pattern. Asp Appl Biol 46:95–102. https://doi.org/10.1890/0012-9658(1998)079[1008:MOSPFC]2.0.CO;2
Perry JN, Winder L, Holland JM, Alston RD (1999) Red-blue plots for detecting clusters in count data. Ecol Lett 2(2):106–113. https://doi.org/10.1046/j.1461-0248.1999.22057.x
Perry GL, Miller BP, Enright NJ (2006) A comparison of methods for the statistical analysis of spatial point pattern in plant ecology. Plant Ecol 187:59 82–59 82. https://doi.org/10.1007/s11258-006-9133-4
Peters R (1992) Ecology of beech forests in the Northern Hemisphere. Ph. D. thesis, Wageningen Agricultural University, Wageningen, The Netherlands
Peters R (1995) Architecture and development of Mexican beech forest. In: Box EO et al (eds) Vegetation science in forestry. Holland, Kluwer Academic Publishers, Dordrecht, pp 325–343
Peters R (1997) Beech forests. Dordrecht, The Netherlands: Kluwer Academic Publishers, https://doi.org/10.1007/978-94-015-8794-5
Peterson CJ, Carson WP, Mccarthy BC, Pickett STA, Peterson CJ, Carson WP, Mccarthy BC, Pickett STA, Microsite STA (1990) Microsite variation and soil dynamics within newly created treefall pits and mounds. Oikos 58:39–46. https://doi.org/10.2307/3565358
Piovensan G, Adams JM (2001) Masting behaviour in beech: linking reproduction and climatic variation. Can J Bot 79:1039–1047. https://doi.org/10.1139/b01-089
Piovensan G, Adams JM (2005) The evolutionary ecology of masting: does the environmental prediction hypothesis also have a role in mesic temperate forests? Ecol Res 20:739–743. https://doi.org/10.1007/s11284-005-0096-z
Queenborough SA, Burslem DFRP, Garwood NC, Burslem P, Garwood C (2007) Neighborhood and community interactions determine the spatial pattern of tropical tree seedling survival. Ecology 88:2248–2258. https://doi.org/10.1890/06-0737.1
Rivers M, Beech E, Murphy L, Oldfield S (2016) Magnoliaceae revised and extended. UK: Botanic Gardens Conservation International
Roberts-Pichette P, Gillespie L (1999) Terrestrial vegetation biodiversity monitoring protocols. EMAN Occasional Paper Series, Report No. 9. Burlington, Ontario, Canada: Ecological Monitoring Coordinating Office
Rodríguez-Ramírez EC, Sánchez-González A, Ángeles-Pérez G (2013) Current distribution and coverage of Mexican beech forests Fagus grandifolia subsp. mexicana in Mexico. Endanger Species Res 20:205–216. https://doi.org/10.3354/esr00498
Rodríguez-Ramírez EC, Sánchez-González A, Ángeles-Pérez G (2016) Relationship between vegetation structure and microenvironment in Fagus grandifolia subsp. mexicana forest relicts in Mexico. J Plant Ecol 12:rtw138. https://doi.org/10.1093/jpe/rtw138
Rowden A, Robertson A, Allnutt T, Heredia S, Williams-Linera G, Newton AC (2004) Conservation genetics of Mexican beech, Fagus grandifolia var. mexicana. Conserv Genet 5:475–484. https://doi.org/10.1023/B:COGE.0000041028.02423.c0
SEMARNAT (Secretaría del Medio Ambiente y Recursos Naturales) (2010) Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección ambiental. Especies nativas de México de flora y fauna silvestres, Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio. Lista de especies en riesgo, Diario Oficial de la Federación. Mexico, Distrito Federal. Available at http://biblioteca.semarnat.gob.mx/janium/Documentos/Ciga/agenda/DOFsr/DO2454.pdf Accessed 22 June 2016
Shibata M, Tanaka H, Iida S, Abe S, Masaki T, Niitama K, Nakashizuka T (2002) Synchronized annual seed production by 16 principal tree species in a temperate deciduous forest, Japan. Ecology 83:1727–1742. https://doi.org/10.1890/0012-9658(2002)083[1727:SASPBP]2.0.CO;2
Snell RS, Huth A, Nabel JEMS, Bocedi G, Travis JMJ, Gravel D, Bugmann H, Gutiérrez AG, Hickler T, Higgins SI, Reineking B, Scherstjanoi M, Zurbriggen N, Lischke H (2014) Using dynamic vegetation models to simulate plant range shifts. Ecography 37(12):1184–1197. https://doi.org/10.1111/ecog.00580
Sork VL, Bramble J, Sexton O (1993) Ecology of mast fruiting in three species of Missouri oaks, Quercus alba, Quercus rubra & Quercus velutina (Fagaceae). Ecology 74:528–541. https://doi.org/10.2307/1939313
Szymura TH, Szymurz M, Maciol A (2015) The effect of ecological niche and spatial pattern on the diversity of oak forest vegetation. Plant Ecol Divers 8:505–518. https://doi.org/10.1080/17550874.2015.1010186
Taugourdeau O, Sabatier S (2010) Limited plasticity of shoot preformation in response to light by understorey saplings of common walnut (Juglans regia). AoB Plants 1:1–8. https://doi.org/10.1080/17550874.2015.1010186
Téllez-Valdés O, Dávila-Aranda P, Lira-Saade R (2006) The effects of climate change on the long-term conservation of Fagus grandifolia var. mexicana, an important species of the Cloud Forest in Eastern Mexico. Biodivers Conserv 15:1095–1107. https://doi.org/10.1007/978-1-4020-5208-8_3
Tinoco-Rueda JA (2009) Clima y variabilidad climática en los municipios de Hidalgo con presencia de bosque mesófilo de montaña. In: Monterroso-Rivas AI (ed) El bosque mesófilo de montaña en el estado de Hidalgo: perspectiva ecológica frente al cambio climático, Vol. 1. Texcoco, Mexico, Universidad Autónoma Chapingo, pp 71–98
Tomita M, Hirabuki Y, Seiwa K (2002) Post-dispersal changes in the spatial distribution of Fagus crenata seeds. Ecology 83:1560–1565
Tuomisto H, Ruokolainen L, Ruokolainen K (2012) Modelling niche and neutral dynamics: on the ecological interpretation of variation partitioning results. Ecography 35:961–971. https://doi.org/10.1111/j.1600-0587.2012.07339.x
Vovides PA, Iglesias CG (1996) Seed germination of Magnolia dealbata Zucc. (Magnoliaceae), an endangered species from Mexico. Hortic Sci 31:877
Wang Y, Camarero JJ, Luo T, Liang E (2012) Spatial patterns of Smith fir alpine treeline on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns. Plant Ecol Divers 5:311–321. https://doi.org/10.1080/17550874.2012.704647
Williams-Linera G (2007) El bosque de niebla del centro de Veracruz: ecología, historia y destino en tiempos de fragmentación y cambio climático. CONABIO–Instituto de Ecología, A.C, Xalapa, Veracruz, Mexico
Williams-Linera G, Rowden A, Newton AC (2003) Distribution and stand characteristics of relict populations of Mexican beech (Fagus grandifolia var. mexicana). Biol Conserv 109:27–36. https://doi.org/10.1016/S0006-3207(02)00129-5
Winder L, Alexander CJ, Holland JM, Woolley C, Perry JN (2001) Modelling the dynamic spatio-temporal response of predators to transient prey patches in the field. Ecol Lett 4:568–576. https://doi.org/10.1046/j.1461-0248.2001.00269.x
Yan Y, Zhang C, Wang Y, Zhao X, von Gadow K (2015) Drivers of seedling survival in a temperate forest and their relative importance at three stages of succession. Ecol Evol 5:4287–4299. https://doi.org/10.1002/ece3.1688
Yuan Z, Gazol A, Wang X, Xing D, Lin F, Bai X, Zhao Y, Li B, Hao Z (2011) What happens below the canopy? Oikos 000:001–009. https://doi.org/10.1111/j.1600-0706.2011.19757x
Zhang Q, Jiang M, Chen F (2007) Canopy recruitment in the beech (Fagus engleriana) forest of Mt. Shennongjia, Central China. J For Res 12:63–67. https://doi.org/10.1007/s10310-006-0249-0
Acknowledgements
We thank Brenda Muñoz-Vazquez, Guadalupe Pérez-Paredes, Rodrigo Ortega, and Othón Álcantara-Ayala for their support during fieldwork. The first author thanks the postdoctoral fellowship DGAPA-UNAM 2015-2016.
Funding
This research was financed by the projects PAPIIT IV201015 and IN223218.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Laurent Bergès
Contribution of the co-authors
ECHRR contributed in the research designing the experiment, writing the paper, and running the data analyses; APMF helped writing the paper and running the data analyses; and ILV supervised and revised all the project stages, including the manuscript writing.
Electronic supplementary material
ESM 1
(DOCX 19 kb)
Rights and permissions
About this article

Cite this article
Rodríguez-Ramírez, E.C., Martínez-Falcón, A.P. & Luna-Vega, I. Spatial patterns of Mexican beech seedlings (Fagus grandifolia subsp. mexicana (Martínez) A.E. Murray): influence of canopy openness and conspecific trees on recruitment mechanisms. Annals of Forest Science 75, 27 (2018). https://doi.org/10.1007/s13595-018-0698-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-018-0698-6