
Abstract
Intensification and homogenization of agricultural landscapes have led to a strong decline in European farmland birds. Agroforestry systems, which were widespread in the past, are regaining attention as they could return structural heterogeneity to agricultural landscapes. However, few studies focus on the effects of such systems on biodiversity and especially bird diversity. We hypothesized that agroforestry systems host a higher alpha and beta diversity of birds compared to open agriculture as well as distinct bird communities. Moreover, we expected that bird communities in temperate Europe and the Mediterranean are differently affected by agroforestry systems. In this study, we assessed breeding bird diversity via audio recordings in nineteen mature agroforestry plots, comprising both silvoarable and silvopastoral systems distributed across seven countries in temperate and Mediterranean Europe. For comparison, bird diversity was also assessed in nearby open agricultural land, forests, and orchards. Bird species richness in agroforestry was more than doubled compared to open agricultural land and similar to the diversity found in forests and orchards. Community composition and within-habitat beta diversity differed between the habitat types and between European regions. While temperate agroforestry systems hosted generalist and woody habitat species, bird communities in Mediterranean agroforestry were composed of species from both open and woody habitats. Beta diversity was significantly higher in agroforestry than in open agriculture in temperate systems but not in the Mediterranean. Our study demonstrates that agroforestry systems represent a valuable habitat for breeding birds in European agricultural landscapes. A wider adoption of these systems could thus contribute to halting and reversing the decline in bird diversity, especially in temperate agricultural landscapes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Across Europe, farmland biodiversity has been declining markedly over the last decades (Benton et al. 2002; Donald et al. 2001). The main reason for the decline is agricultural intensification, which started in the second half of the twentieth century, aiming to increase agricultural yields (Pain and Pienkowski 1997; Tscharntke et al. 2005). Consequences that arose from changes in farming practices are numerous, including an increase in field sizes, often associated with a reduction of semi-natural and edge habitats, higher stocking densities, a reduction of crop diversity, and an increased use of agrochemicals and pesticides (Newton 2004; Pain and Pienkowski 1997; Robinson and Sutherland 2002; Stoate et al. 2001). This led to a considerable loss of heterogeneity at both field and landscape scales resulting in habitat loss for many taxa including birds (Benton et al. 2002; Donald et al. 2006; Voříšek et al. 2010; Kamp et al. 2021). As the global demand for agricultural products will continue to increase during the twenty-first century (Tilman et al. 2011), there is an urgent need to identify and implement food production systems that maintain and restore biodiversity (Bommarco et al. 2013).
In contrast to intensified open farmland, agroforestry systems provide a high structural heterogeneity and incorporate woody plants. Agroforestry exists in a remarkable diversity all over Europe but was much more widespread in the past (Nerlich et al. 2013). In agroforestry, trees and shrubs are deliberately combined with crops and/or livestock on the same land (Fig. 1) (Nair 1993). Benefits can be both ecological and economic, resulting from interactions between the woody and non-woody components (Mosquera-Losada et al. 2009, http://apps.fao.org/). An example in temperate Europe are the so-called Streuobstwiesen, i.e., orchards combining fruit trees with either cropland, meadows, or pastures (Herzog 1998). In the Iberian Peninsula, there are the savannah-like dehesas (Spain) and montados (Portugal), in which oak-grassland ecosystems have been shaped by centuries of pastoralism and human management (Joffre et al. 1999). These systems support high biodiversity and are culturally significant (Moreno and Pulido 2009; Moreno et al. 2018). By (re-)integrating structural heterogeneity into agricultural landscapes, agroforestry systems have the potential to be part of a more resilient and sustainable agriculture. Providing a compromise between production demands and nature conservation needs, these systems have recently regained attention (Jose 2009; Quinkenstein et al. 2009; Nerlich et al. 2013).

a A silvopastoral site with holm oak and cattle-grazed pastures in the dehesas of Extremadura, Spain. b A silvoarable site with apple trees and strawberries near Sursee, Switzerland.
In agroforestry systems, productivity can be increased compared to arable cropland and woodland systems managed separately through the complementary use of light, nutrients, and water as well as the production of different goods, e.g., forage, wood products, fodder, crops, and livestock (Cannell et al. 1996; Graves et al. 2007). Additionally, agroforestry systems support a wide variety of ecosystem services (Fagerholm et al. 2016; Jose 2009) such as carbon storage (Nair et al. 2009), reduction of soil erosion and leaching of nutrients, and improvement of soil fertility and nutrient cycling (Nair 2007; Torralba et al. 2016) as well as water retention (Joffre and Rambal 1988). Due to their high internal heterogeneity, agroforestry systems offer a diversity of food resources and habitat structures (McArthur 1958) and have thus also been found to support biodiversity (Jose 2009; McAdam and McEvoy 2009; Torralba et al. 2016). However, only a few studies focused on the effects of European agroforestry systems on bird diversity (McAdam et al. 2007; Mupepele et al. 2021). Many bird species in farmland depend on the presence of trees and shrubs (Jakobsson and Lindborg 2017; Rösch et al. 2023). Thus, the benefits of agroforestry on birds can be expected to be strong. In contrast to annual crops, agroforestry systems are passing through different stages of tree maturity over decades. These stages may be preferred by different species of birds, similar to the successional stages of forests (Helle and Mönkkönen 1985). Thus, a higher turnover of bird species (β diversity) can be expected among agroforestry systems than among open farmland. Other studies have dealt with wood pastures (a type of silvopasture) and showed that bird diversity was higher in wood pastures compared to open pastures (Hartel et al. 2014; Morgan-Davies et al. 2008). However, the studied wood pastures served mostly for conservation management in forested areas and not for food production in agricultural landscapes, on which we focus in the current study.
Our objective was to compare breeding bird diversity of agroforestry with purely woody and agricultural reference areas. We used acoustic recording on eight agroforestry sites comprising nineteen mature agroforestry plots across seven countries in temperate and Mediterranean Europe.
Specifically, we tested the following hypotheses: (1) Due to a high structural heterogeneity provided by the presence of trees, bird species richness is higher in agroforestry systems compared to open agriculture and similar to bird richness in orchards and forests. (2) European agroforestry systems, with their combination of trees and open vegetation, support distinct bird communities composed of species from both open and woody habitats. (3) Due to the variety of agroforestry systems across Europe and their variation in tree maturity, their β diversity is higher than in open agriculture. (4) Due to differences in the structure of forests and open agricultural land, the effects of agroforestry on bird diversity differ between temperate Europe and the Mediterranean.
2 Material and methods
2.1 Study sites
The study was carried out in eight different agroforestry sites with either silvoarable (combination of trees and crops) or silvopastoral systems (combination of trees and livestock) and nearby control plots. The sites were located in seven countries across temperate Europe (England, Switzerland, Central France, Germany; hereafter “temperate sites”) and Southern Europe (Southern France, Italy, Portugal, and Spain; hereafter “Mediterranean sites”).
In each site, one to three agroforestry plots with mature trees (at least 15 years old) were selected. Reference areas were chosen to contain the same woody and agricultural component as the agroforestry system. Thus, open pastures served as a reference for silvopastoral systems and cropland for silvoarable systems. For the woody component, we used forest as a reference area, and added orchards as a reference where the tree component of agroforestry was fruit or nut trees. Forests were selected according to the following definition “land with tree cover or equivalent stocking level of more than 10% and with an area of more than 0.5 hectares” (FAO, http://apps.fao.org/). Moreover, in this study, the control plots defined as forests contained two or more different tree species with part of the trees aged more than 30 years. Conversely, in orchards, i.e., fruit or nut-producing orchards, only trees of the same species and of similar age as in the respective agroforestry plot were present.
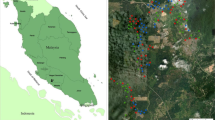
In the temperate sites, agroforestry plots, forests, and orchards were dominated by broad-leaved trees. In the Mediterranean sites, tree stands consisted of walnuts, oaks, or pines. The agroforestry sites in England, Switzerland, and Southern France were silvoarable systems with trees aged 24, 14, and 27 years, respectively. Silvopastoral sites contained trees aged 20 (Germany), 35 (Central France), 90 (Italy), 25 (Portugal), and 175 (Spain) years. The agroforestry and reference plots were always located in the same region (see Table S1), to minimize the variation of, e.g., climate, soil, and the surrounding landscape. A total of nineteen agroforestry plots (six silvoarable and thirteen silvopastoral plots), eighteen open agricultural plots, fifteen forests, and eight orchards were sampled. The geographic location of the study sites is shown in Fig. 2. Detailed characteristics of the sampled silvoarable and silvopastoral agroforestry plots as well as the associated control plots are summarized in Table S1.

Geographic location of the study sites. triangles = silvoarable systems, circles = silvopastoral systems, A = Wakelyns (UK), B = Sursee (CH), C = Restinclières (FR), D = Bannmühle (DE), E = Lamartine (FR), F = Tenuta di Paganico (IT), G = Dehesa de Majadas (ES), H = Moinhos de Vento (PO).
2.2 Data collection
Bird diversity was assessed using autonomous sound recorders (AudioMoth V1.1.0, Open Acoustic Devices, firmware version 1.7., configuration app version 1.6.0). They were set up three times over the course of the breeding season (between March and July) in 2021 (England, Germany, Switzerland, and Central France) and 2022 (Southern France, Italy, Portugal, and Spain, see Table S1 for the detailed sampling dates). On each sampling date, one AudioMoth was set up in each plot, at least 20 m from the plot’s edge, with a microphone oriented towards the plot’s center. AudioMoths were wrapped in a single layer of clingfilm to protect them from humidity and dust. They were attached to a pole of a smaller diameter than the AudioMoths, 1.5 m above the ground. Surveys were conducted only during calm and dry weather, as bird activity can be strongly reduced by wind or rain (Bibby et al 2000; Pijanowski et al. 2011). During each of the three samplings, the AudioMoths were programmed to record continuously from 1 h before sunrise until 2 h after sunrise. To reduce possible hardware bias, recorders were rotated between plots after each sampling. Devices were programmed to record with a sampling rate of 192 kHz, a medium gain, a recording duration of 59 min and 55 s, and a sleep duration of 5 s. Recordings were saved as .wav files and stored on memory cards (SanDisk Corporation, Milpitas, CA, USA). Information about plots, their management and characteristics, and environmental parameters as well as the sampling dates were collected through a sampling sheet.
2.3 Species identification
Prior to analyses, all audio recordings were resampled at 22050 Hz, the conventional sampling rate for audible sounds (Gibbs et al. 2016) in order to improve frequency resolution.
For each plot and each of the three sampling dates, we analyzed two 10-min recordings: the first one starting at sunrise, the second starting 1 h after sunrise, since this falls within the highest singing activity of breeding birds (Bibby et al. 2000). Thus, in total 60 min were analyzed per plot. All species present in the recordings were identified aurally and visually with the help of the software Audacity® (version 5.4.8) with a 1024-point Hann window spectrogram, showing frequency variations over time. The databases Xeno-canto (www.xeno-canto.org), e-bird (www.ebird.org), and Tierstimmenarchiv (www.tierstimmenarchiv.de) were used to verify species identifications. Species that cannot be safely distinguished from closely related ones based on their songs or calls were identified to genus level, in our case Passer sp. and Galerida sp.
In each recording, all species identified by their call or song (hereafter “vocalization”) were annotated using Audacity. For each bird individual, the duration in seconds of its vocalization (maximum of 600 s) was measured. In addition, the maximal relative sound level (RSL) measured in decibels (dB) and its associated frequency in Hertz were measured using the software Kaleidoscope Pro (version 5.4.8; Wildlife Acoustics Inc., Concord, MA, USA). The parameters used were an FFT (fast Fourier transform) size of 512 and a win size of 128 in an overlapping Hann window. The RSL was measured by selecting the area around the loudest vocalization in the recording and was used as an indicator for the distance of the vocalizing individuals from the recorder (Yip et al. 2017). In the case of two individuals from the same species singing simultaneously (in 8 cases or 2.22% of the −30dB vocalization dataset (see below)), both were assessed to better represent the total species abundance. All vocalizations shorter than 5 s were removed from the dataset (short calls, probably not being territorial vocalizations) as well as aquatic species that were not associated with the study plots but with ponds located nearby. Vocalizations were included only above −30 dB, since this is the loudness typically shown by species singing within 20 m of the recorder (Manon Edo, unpublished data). This way, we made sure that the analyzed species were mostly associated with the habitat type in which the AudioMoth was located. Birds can be detected with AudioMoths over several hundred meters distance, which could have been far outside the focal habitat in many cases. Thus, the use of a minimum loudness was an important step, although it reduced the number of analyzed bird vocalizations by 75%, eliminating some species from the dataset (e.g., skylark, swallows) that were singing or calling outside the study plot.
2.4 Statistical analysis
To compare the bird diversity between the four different habitat types, different diversity indices and statistical analyses were used. In order to analyze bird species richness, we fitted mixed-effects models (R package lme4, function lmer, Bates et al. 2015) with habitat type and climate zone (temperate/Mediterranean) as explanatory variables (equation: lmer (SpR ~ Habitat + Climatic Zone + (1|Site)). Due to the nested design of the study, “Site” was used as a random factor. A post hoc test was used to determine pairwise differences between habitats (R package emmeans, function emmeans, Lenth 2022). Due to strong correlations between bird species richness, the number of recorded individuals, and singing activity in seconds, all analyses are based on species richness alone. To optimize the sample size, silvoarable and silvopastoral plots were analyzed together. A separate analysis of the species richness between the different habitat types for silvoarable versus silvopastoral sites is found in Supplementary Fig. S3; species richness showed the same trend between habitats for both types of systems. Bird community composition was analyzed using partial redundancy analyses (RDAs) based on the number of recorded individuals per species over the season with habitat as an explanatory variable and country as a conditional variable (R package vegan, function rda, Oksanen et al. 2013). Only species that occurred on more than two plots were retained in the analysis. Prior to analysis, the community data matrices were Hellinger-transformed, thereby giving lower weights to rare species (Legendre and Gallagher 2001). Due to strong differences in species composition, the Mediterranean sites and the temperate sites were analyzed separately. A permutation test with 9999 permutations was used to determine the significance of the results (R package vegan, function permutest, Oksanen et al. 2013). Finally, species turnover (β diversity) among habitats of the same type was assessed based on Euclidean distances for Mediterranean and temperate sites, respectively (R package vegan, functions vegdist and betadisper, Oksanen et al. 2013). The similarity of composition was calculated as distance to centroid and was compared between habitat types. Statistical significance was assessed through a permutation test with 9999 permutations (R package vegan, function permutest, Oksanen et al. 2013).
For all statistical tests, p-values lower than 0.05 were considered significant. P-values, means, and standard errors of mean (± SE) are given in text and tables. All statistical analyses were conducted in R version 4.2.2 (R Core Team 2022). Information about species habitat specialization (Open, Forest, or Generalist) indicated in Table 1 were found at https://www.iucnredlist.org/en. Species categorized as “Open habitat species” are breeding in “arable land,” “grassland,” or “pastureland,” but not forest. “Forests species” are those breeding in “forest,” but none of the mentioned open habitat types. As an exception, species with forest as breeding habitat of “major importance” were classified as forest species even if open habitats were listed for breeding, as well (e.g., the common chaffinch, Fringilla coelebs). Species categorized as “Generalist” are those associated with “forest” AND “arable land,” “grassland,” or “pastureland.” One species with neither forest, arable, or pastureland as breeding habitat is categorized as “other.”
3 Results
3.1 Species richness
In total, 48 different bird species were recorded (Table 1). Fourteen species were recorded in England, 16 in Germany, 8 in Switzerland, 19 in Central France, 13 in Southern France, 7 in Italy, 22 in Spain, and 8 in Portugal. In the temperate sites, the most common species were blackcap Sylvia atricapilla, common blackbird Turdus merula, wren Troglodytes troglodytes, robin Erithacus rubecula, and common chaffinch Fringilla coelebs. In the Mediterranean sites, common chaffinch Fringilla coelebs, corn bunting Emberiza calandra, nightingale Luscinia megarhynchos, crested or thekla lark Galerida sp., and short-toed treecreeper Certhia brachydactyla were the most commonly found species. The total number of species varied between habitats, with 34 species recorded in agroforestry, 28 in forests, 15 in orchards, and 13 in open agriculture.
The highest bird species richness per site was recorded in forests (5.20 ± 0.51), followed by agroforestry (3.63 ± 0.53), orchards (2.38 ± 0.78), and open agriculture (1.06 ± 0.31) (Fig. 3). Bird species richness in forests and agroforestry was significantly higher than in open agriculture. Moreover, species richness in forests was significantly higher than in orchards (Fig. 3, Table S2).

Bird species richness in agroforestry (n = 19), forest (n = 15), orchard (n = 8), and open agriculture (cropland/pasture) (n = 18). Significance levels: *P < 0.05, **P < 0.01, ***P < 0.001. See Table S2 for the detailed results of species richness in the different habitats as well as the results of mixed-effect models and post hoc tests used to determine the pairwise differences between habitats.
3.2 Community composition
The four habitat types had distinct bird communities in both temperate and Mediterranean Europe. However, in the temperate sites, the bird communities of the different habitats were more clearly separated from each other than in the Mediterranean (Fig. 4a, b). Across both climate zones, generalist (occurring in a wide range of habitats, defined here as species breeding in forest and arable or pastureland) and forest species were associated with both agroforestry and forests, e.g., chaffinch Fringilla coelebs, blackbird Turdus merula, and great and blue tit Parus major and Cyanistes caeruleus. In temperate Europe, typical species of woodland areas such as wren Troglodytes troglodytes and blackcap Sylvia atricapilla as well as generalist species, e.g., green woodpecker Picus viridis and robin Erithacus rubecula were found in both forests and agroforestry. Only few species and individuals (blackbird Turdus merula, chaffinch Fringilla coelebs, blue tit Cyanistes caeruleus, and wood pigeon Columba palumbus) were observed in temperate orchards and open agriculture. In the Mediterranean, forest species such as golden oriole Oriolus oriolus and short-toed treecreeper Certhia brachydactyla were common in both agroforestry and forest. Species typical for open or semi-open areas such as corn-bunting Emberiza calandra, Galerida sp. larks, and spotless starling Sturnus unicolor were present in both open agriculture and agroforestry of the Iberian Peninsula. The corn-bunting Emberiza calandra was also present in the Spanish forests and larks Galerida sp. in the Portuguese pine orchards. The European serin Serinus serinus, a generalist species, was recorded in Mediterranean agroforestry and forest plots.

Redundancy analysis (RDA) plots of the bird species communities in different habitats: agroforestry, forests, orchards, and open agriculture (cropland or pastures) in a temperate climate (England, Germany, Switzerland, and Central France) and b Mediterranean climate (South of France, Italy, Spain, and Portugal). Dashed lines represent minimum convex polygons around habitat types.
3.3 β diversity within habitat types
In temperate Europe and the Mediterranean, β diversity (average distance to centroid) was significantly higher among forest plots than among open agriculture and orchards (Fig. 5a, b). Moreover, in temperate Europe, β diversity was significantly higher among agroforestry than among open agriculture. The low distances to centroid in temperate open agriculture and to a lesser extent in orchards are probably largely due to the low species richness (Fig. 3), with those habitats having hardly any birds recorded.

β diversity within habitat types (agroforestry, forest, orchard, open agriculture (cropland/pasture)) in a temperate sites (England, Germany, Switzerland, and Central France) and b Mediterranean sites (South of France, Italy, Spain, and Portugal). The y-axis shows the Euclidean distance to the centroid of the respective land-use type.
In contrast, in the Mediterranean, the β diversity in agroforestry was not significantly different from the other habitats (Fig. 5a). Mediterranean open agriculture showed a higher distance to centroid compared to the temperate plots.
4 Discussion
This study clearly underpins the potential of mature agroforestry systems as a habitat of high value for birds in agricultural landscapes. As hypothesized, bird species richness in agroforestry systems was higher than in open agriculture and similar to the species richness found in orchards and forests. In the Mediterranean, bird communities in agroforestry included both open and woody habitat species. However, in temperate Europe, no open habitat species were present in agroforestry plots which hosted only generalist and woody habitat bird species. As expected, in the temperate region, β diversity was significantly higher in agroforestry and forest than in open agriculture. Contrary to our expectations, β diversity within Mediterranean agroforestry did not differ significantly from other habitats.
4.1 Species richness
Our results are in line with other studies which demonstrated that silvopastoral systems are hosting a greater bird diversity than open pastures (Hartel et al. 2014; McAdam et al. 2007; Morgan-Davies et al. 2008). The high bird diversity we found in both agroforestry and forest plots, and to a lesser extent in orchards, can be explained by the heterogeneity of these habitats. Structural diversity increases niche availability, providing more foraging, shelter, and nesting sites for birds over the season (Morelli 2013; Söderström et al. 2001). Old and dead trees with hollows and crevices, e.g., under the bark (Godinho and Rabaça 2011) represent habitats of high value for birds, especially for cavity-nesting species. Poulsen (2002) showed that the number of old trees, tree species, and tree size classes in Danish forests is correlated with a higher number of bird species and individuals. Moreover, a well-developed understory plays an important role for many shrub-dependent species (Fuller et al. 2007; Hartel et al. 2014). This diversity of niches is not only beneficial for birds, but for other taxa as well. McAdam et al. (2007) demonstrated that arthropods, an important food resource for birds (Holland et al. 2006), were more abundant in agroforestry and forest than in conventional agricultural fields. Pardon et al. (2019) and Boinot et al. (2019) highlighted the value of understory vegetation and grass strips for overwintering arthropods in temperate arable agroforestry. Söderström et al. (2001) showed that an increased heterogeneity and diversity in the shrub and tree layer within pastures lead to a higher species richness of insects and birds.
The lower diversity we found in orchards compared to forests is probably due to their higher disturbance and lower structural complexity, including a lower understory heterogeneity (Bohada-Murillo et al. 2019) compared to multi-layer and long-established forest ecosystems. As opposed to agricultural areas, woodlands show positive effects on the abundance of non-migratory birds in Germany (Schulze et al. 2019), which is confirmed by the high diversity we found in all forest plots.
Hence, higher ecosystem complexity and higher food availability in woody habitats, and particularly in forests and agroforestry sites, probably explain the higher bird species richness we found in those habitats in comparison with open agricultural land, in which the habitat spectrum for birds and other taxa is reduced. Our results are in line with other studies; however, due to differences in study design, site characteristics, and historical and geographical context, caution is needed with regard to comparability between results (Mupepele et al. 2021). The surrounding landscape is known to highly affect bird communities (Broughton et al. 2021; Söderström and Pärt 2000; Söderström et al. 2001). Söderström et al. (2001) found that pastures surrounded by arable land hosted a lower bird richness than pastures surrounded by forests. In the same way, isolated orchards showed a negative effect on birds whereas habitat connectedness, i.e., proximity to forests, led to a higher bird species richness (Bailey et al. 2010). Moreover, land use history strongly affects biodiversity, which thus varies between older and newly established agroforestry systems (Mupepele et al. 2021). The value of tree-associated habitats increases with tree age (Poulsen 2002). We therefore expect the value of agroforestry systems for biodiversity to increase with time as well. The lack of biodiversity-related studies conducted in silvoarable systems, and especially alley-cropping systems, underlines that research in this area should be prioritized. Finally, several studies demonstrate the high value of birds as predators of invertebrates that support pest control services in agroecosystems (Barbaro et al. 2017; García et al. 2018). For example, great and blue tits, two species associated with agroforestry systems in our study, were identified as complementary predators of apple pest insects (García et al. 2021). In the present context of global change and the drastic decline in bird diversity and abundance, agroforestry sites could thus represent refuges for birds that support biological pest control in agricultural landscapes (Tscharntke et al. 2005).
4.2 Community composition and β diversity
Bird community composition differed between the four habitat types. In the Mediterranean sites, the results confirmed that bird communities in agroforestry comprised species of both open and woody habitats. However, in temperate sites, the bird communities of the agroforestry plots were mainly composed of generalist and woody habitat species while open agriculture and orchards were associated with a low diversity. In temperate Europe, β diversity was higher in agroforestry and forests compared to open agriculture. Similarly, in the Mediterranean, β diversity was higher in forests compared to open agriculture but no significant difference in β diversity between agroforestry and open agriculture was found. Temperate open agriculture showed a low β diversity compared to the same habitat in the Mediterranean. The differences observed between temperate and Mediterranean sites, especially concerning the open and semi-open habitats, can partly be explained by differences in agricultural intensity. The highly intensive farming in north-western Europe, associated with high crop yields and fertilizer applications, has led to a sharp decline in birds of open farmland habitats (Reif and Hanzelka 2020). This explains the low bird species richness as well as β diversity in temperate open agriculture. Almost no bird species of open and semi-open habitat were found in the temperate zone—neither in agroforestry nor in open agriculture. Birds of open and semi-open agricultural landscapes such as common whitethroat Curruca communis, cirl bunting Emberiza cirlus, or yellowhammer Emberiza citrinella have generally become rare in temperate Europe. In contrast, in the Mediterranean, due to more extensive land-use systems, open farmland habitats in many areas still host a high biodiversity of birds and other taxa (Kleijn et al. 2009).
Given the higher species richness and β diversity of birds that we found in temperate agroforestry compared to open agriculture, this habitat is of high value for birds in temperate farmland. The presence of species relying on trees/shrubs for nesting and/or foraging in the agroforestry sites, such as green woodpecker Picus viridis, wren Troglodytes troglodytes, and great and blue tit Parus major and Cyanistes caeruleus confirm this. In temperate Europe, agroforestry systems may be particularly beneficial in the transition zone between forest and open farmland. In the intensively managed and highly fragmented European farmland, agroforestry could act as stepping stones (habitat islands) or ecological corridors and could thus contribute to the facilitation of animal movement and plant dispersion (Fischer et al. 2006; Hidalgo et al. 2021; Jongman et al. 2004).
In accordance with other studies (Tellería 2001), our results show that Mediterranean agroforestry hosts both woodland and open habitat bird species. Open habitat species, such as larks Galerida sp. and corn bunting Emberiza calandra, relying on low vegetation for nesting and foraging, were present in agroforestry as well as in open agriculture. Moreover, generalist/semi-open habitat species like European serin Serinus serinus and great tit Parus major were also found in agroforestry sites, as well as in forests. Finally, the presence of species such as short-toed treecreeper Certhia brachydactyla, blue tit Cyanistes caeruleus, or golden oriole Oriolus oriolus in both forest and agroforestry confirms the value of agroforestry for woody habitat species that rely on dense woody vegetation for nesting and/or foraging (Godinho and Rabaça 2011). Those results are in line with studies showing that the Spanish dehesas, whose semi-open physiognomy and habitat diversity are the result of centuries of extensive grazing management, act as an ecotone habitat, hosting a high bird diversity composed of species relying on open and border habitat as well as forest species (Tellería 2001).
In the Mediterranean, the lower differences in species composition and β diversity that we measured between habitats (distance to centroid) could be due to a higher habitat and landscape heterogeneity compared to the temperate sites. For example, in the open pasture plots in Spain, scattered trees or shrubs provided valuable structures for birds. In the same way, woody control plots often showed gaps with low vegetation or with a lower tree density. Therefore, numerous tree/shrub-dependent species, e.g., nightingale Luscinia megarhynchos or chaffinch Fringilla coelebs, occurred in Mediterranean forests and agroforestry as well as open agriculture.
Our findings suggest that agroforestry, through increasing heterogeneity at field and landscape scale, favors mostly generalist bird species as also supported by other authors (Batáry et al. 2011; Filippi-Codaccioni et al. 2010; Söderström et al. 2001; Pickett and Siriwardena 2011). In contrast, as shown by numerous studies (Báldi and Batáry 2011; Batáry et al. 2011; Hagist and Schürmann 2021; Reif and Hanzelka 2020), some specialist species, such as birds of semi-natural grassland or arable land such as the skylark Alauda arvensis, are not favored or can even be deterred by the establishment of woody structures. Hence, the implementation of new agroforestry systems needs to be adapted to regional conditions and the geographical as well as historical context (Reisner et al. 2007).
Our results show that agroforestry can support a high diversity of breeding birds in both temperate and Mediterranean Europe; however, the added value of those systems differs between the regions. As demonstrated within this study, agroforestry can act as an important refuge for generalist and forest species in intensively managed, tree-poor agricultural areas in temperate Europe. However, they do not necessarily bring back the farmland birds that have declined so drastically over the past decades (Voříšek et al. 2010). In southern Europe where open semi-natural habitats and low-intensity agricultural systems with high bird species richness are still relatively common but threatened by both agricultural intensification and abandonment, agroforestry systems can represent an important intermediate habitat for both open/semi-open habitat and forest bird species.
5 Conclusion
To our knowledge, this is the first study examining the bird diversity in mature European agroforestry systems across multiple countries. Confirming our hypothesis, we found that agroforestry systems supported bird diversity as they hosted a higher species richness than open agricultural systems in temperate as well as in Mediterranean countries. However, the effect of agroforestry systems on bird diversity was different in temperate compared to Mediterranean sites and hence highly dependent on the regional context. Through providing heterogeneity in agricultural landscapes, agroforestry systems may help to restore farmland bird diversity in temperate regions of Europe and to halt biodiversity loss in the Mediterranean. In general, agroforestry systems are a promising example of agricultural production that is compatible with biodiversity conservation. Positive effects of agroforestry on ecosystem functions and resilience could contribute to ecological intensification, e.g., via improved pest regulation also in surrounding crops.
Data availability
The datasets as well as the codes generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable
References
Bailey D, Schmidt-Entling MH, Eberhart P, Herrmann JD, Hofer G, Kormann U, Herzog F (2010) Effects of habitat amount and isolation on biodiversity in fragmented traditional orchards: orchard fragmentation effects on biodiversity. J Appl Ecol 47(5):1003–1013. https://doi.org/10.1111/j.1365-2664.2010.01858.x
Báldi A, Batáry P (2011) Spatial heterogeneity and farmland birds: different perspectives in Western and Eastern Europe. Ibis 153(4):875–876. https://doi.org/10.1111/j.1474-919X.2011.01169.x
Barbaro L, Rusch A, Muiruri EW, Gravellier B, Thiery D, Castagneyrol B (2017) Avian pest control in vineyards is driven by interactions between bird functional diversity and landscape heterogeneity. J Appl Ecol 54(2):500–508. https://doi.org/10.1111/1365-2664.12740
Batáry P, Fischer J, Báldi A, Crist TO, Tscharntke T (2011) Does habitat heterogeneity increase farmland biodiversity? Front Ecol Environ 9(3):152–153. https://doi.org/10.1890/11.WB.006
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Benton TG, Bryant DM, Cole L, Crick HQP (2002) Linking agricultural practice to insect and bird populations: a historical study over three decades: farming, insect and bird populations. J Appl Ecol 39(4):673–687. https://doi.org/10.1046/j.1365-2664.2002.00745.x
Bibby CJ, Burgess ND, Hill DA, Mustoe SH (2000) Bird census techniques, 2nd edn. Academic Press, London. ISBN: 9780120958313
Bohada-Murillo M, Castaño-Villa GJ, Fonturbel FE (2019) The effects of forestry and agroforestry plantations on bird diversity: a global synthesis. Land Degrad Dev 31(5):646–654. https://doi.org/10.1002/ldr.3478
Boinot S, Poulmarc’h J, Mézière D, Lauri PE, Sarthou JP (2019) Distribution of overwintering invertebrates in temperate agroforestry systems: implications for biodiversity conservation and biological control of crop pests. Agr Ecosyst Environ 285:106630. https://doi.org/10.1016/j.agee.2019.106630
Bommarco R, Kleijn D, Potts SG (2013) Ecological intensification: harnessing ecosystem services for food security. Trends Ecol Evol 28(4):230–238. https://doi.org/10.1016/j.tree.2012.10.012
Broughton RK, Chetcuti J, Burgess MD, Gerard FF, Pywell RF (2021) A regional-scale study of associations between farmland birds and linear woody networks of hedgerows and trees. Agr Ecosyst Environ 310:107300. https://doi.org/10.1016/j.agee.2021.107300
Cannell MGR, Van Noordwijk M, Ong CK (1996) The central agroforestry hypothesis: the trees must acquire resources that the crop would not otherwise acquire. Agrofor Syst 34(1):27–31. https://doi.org/10.1007/BF00129630
Donald PF, Green RE, Heath MF (2001) Agricultural intensification and the collapse of Europe’s farmland bird populations. P Roy Soc Lond B Bio 268(1462):25–29. https://doi.org/10.1098/rspb.2000.1325
Donald PF, Sanderson FJ, Burfield IJ, van Bommel FPJ (2006) Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agr Ecosyst Environ 116(3–4):189–196. https://doi.org/10.1016/j.agee.2006.02.007
Fagerholm N, Torralba M, Burgess PJ, Plieninger T (2016) A systematic map of ecosystem services assessments around European agroforestry. Ecol Indic 62:47–65. https://doi.org/10.1016/j.ecolind.2015.11.016
Filippi-Codaccioni O, Devictor V, Bas Y, Clobert J, Julliard R (2010) Specialist response to proportion of arable land and pesticide input in agricultural landscapes. Biol Conserv 143(4):883–890. https://doi.org/10.1016/j.biocon.2009.12.035
Fischer J, Lindenmayer DB, Manning AD (2006) Biodiversity, ecosystem function, and resilience: ten guiding principles for commodity production landscapes. Front Ecol Environt 4 (2):80–86. https://doi.org/10.1890/15409295(2006)004[0080:BEFART]2.0.CO;2
Fuller RJ, Smith KW, Grice PV, Currie FA, Quine CP (2007) Habitat change and woodland birds in Britain: implications for management and future research: habitat change and woodland birds. Ibis 149:261–268. https://doi.org/10.1111/j.1474-919X.2007.00775.x
García D, Miñarro M, Martínez-Sastre R (2018) Birds as suppliers of pest control in cider apple orchards: avian biodiversity drivers and insectivory effect. Agr Ecosyst Environ 254:233–243. https://doi.org/10.1016/j.agee.2017.11.034
García D, Miñarro M, Martínez-Sastre R (2021) Enhancing ecosystem services in apple orchards: nest boxes increase pest control by insectivorous birds. J Appl Ecol 58(3):465–475. https://doi.org/10.1111/1365-2664.13823
Gibbs S, Koblents H, Coleman B, Gordon A, Thevathasan N, Wiliams P (2016) Avian diversity in a temperate tree-based intercropping system from inception to now. Agrofor Syst 90(5):905–916. https://doi.org/10.1007/s10457-016-9901-7
Godinho C, Rabaça JE (2011) Birds like it Corky: the influence of habitat features and management of ‘montados’ in breeding bird communities. Agrofor Syst 82(2):183–195. https://doi.org/10.1007/s10457-010-9345-4
Graves AR, Burgess PJ, Palma JHN, Herzog F, Moreno G, Bertomeu M, Dupraz C, Liagre F, Keesman K, van der Werf W, de Nooy AK, van den Briel JP (2007) Development and application of bio-economic modelling to compare silvoarable, arable, and forestry systems in three European countries. Ecol Eng 29(4):434–449. https://doi.org/10.1016/j.ecoleng.2006.09.018
Hagist D, Schürmann H (2021) Agroforesterie et oiseaux nicheurs – Opportunités et conflits. Fiche Info. Station ornithologique suisse, Sempach. https://www.vogelwarte.ch/modx/assets/files/vogelwarte/news/avinews/Faktenblatt%20Agroforst_fr.pdf. Accessed 17 January 2023
Hartel T, Hanspach J, Abson DJ, Máthé O, Moga CI, Fischer J (2014) Bird communities in traditional wood-pastures with changing management in Eastern Europe. Basic Appl Ecol 15(5):385–395. https://doi.org/10.1016/j.baae.2014.06.007
Helle P, Mönkkönen M (1985) Measuring turnover rates in secondary succession in European forest bird communities. Ornis Scand: 173–180. https://doi.org/10.2307/3676628
Herzog F (1998) Streuobst: a traditional agroforestry system as a model for agroforestry development in temperate Europe. Agrofor Syst 42(1998):61–80. https://doi.org/10.1023/A:1006152127824
Hidalgo PJ, Hernández H, Sánchez-Almendro AJ, López-Tirado J, Vessella F, Porras R (2021) Fragmentation and connectivity of island forests in agricultural Mediterranean environments: a comparative study between the Guadalquivir Valley (Spain) and the Apulia Region (Italy). Forests 12(9):1201. https://doi.org/10.3390/f12091201
Holland JM, Hutchison MAS, Smith B, Aebischer NJ (2006) A review of invertebrates and seed-bearing plants as food for farmland birds in Europe. Ann Appl Biol 148(1):49–71. https://doi.org/10.1111/j.1744-7348.2006.00039.x
Jakobsson S, Lindborg R (2017) The importance of trees for woody pasture bird diversity and effects of the European Union’s tree density policy. J Appl Ecol 54(6):1638–1647. https://doi.org/10.1111/1365-2664.12871
Joffre R, Rambal S, Ratte JP (1999) The dehesa system of southern Spain and Portugal as a natural ecosystem mimic. Agrofor Syst 45:57–79. https://doi.org/10.1023/A:1006259402496
Joffre R, Rambal S (1988) Soil water improvement by trees in the rangelands of southern Spain. Acta Ooecol - Oec Plant 9 (4):405–422. ISBN: 1146-609X
Jongman RHG, Külvik M, Kristiansen I (2004) European ecological networks and greenways. Landsc Urban Plan 68(2–3):305–319. https://doi.org/10.1016/S0169-2046(03)00163-4
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor Syst 76(1):1–10. https://doi.org/10.1007/s10457-009-9229-7
Kamp J, Frank C, Trautmann S, Busch M, Dröschmeister R, Flade M et al (2021) Population trends of common breeding birds in Germany 1990–2018. J Ornithol 162:1–15. https://doi.org/10.1007/s10336-020-01830-4
Kleijn D, Kohler F, Báldi A, Batáry P, Concepción ED, Clough Y, Díaz M, Gabriel D, Holzschuh A, Knop E, Kovács A, Marshall EJP, Tscharntke T, Verhulst J (2009) On the relationship between farmland biodiversity and land-use intensity in Europe. P R Soc B: Biol Sci 276(1658):903–909. https://doi.org/10.1098/rspb.2008.1509
Legendre P, Gallagher E (2001) Ecologically meaningful transformations for ordination of species data. Oecologia 129:271–280. https://doi.org/10.1007/s004420100716
Lenth R (2022) Emmeans: estimated marginal means, aka least-squares means. R package version 1.8.2. https://CRAN.R-project.org/package=emmeans
McAdam JH, Sibbald AR, Teklehaimanot Z, Eason WR (2007) Developing silvopastoral systems and their effects on diversity of fauna. Agrofor Syst 70(1):81–89. https://doi.org/10.1007/s10457-007-9047-8
McAdam JH, McEvoy PM (2009) The potential for silvopastoralism to enhance biodiversity on grassland farms in Ireland. In: A Rigueiro-Rodríguez J, McAdam J, MR Mosquera-Losada (eds) Agroforestry in Europe: current status and future prospects. Springer, Netherlands, Vol. 6, pp 343–356. ISBN:9781402082719
McArthur RH (1958) Population ecology of some warblers of northeastern coniferous forests. Ecology 39(4):599–619. https://doi.org/10.2307/1931600
Morelli F (2013) Relative importance of marginal vegetation (shrubs, hedgerows, isolated trees) surrogate of HNV farmland for bird species distribution in Central Italy. Ecol Eng 57:261–266. https://doi.org/10.1016/j.ecoleng.2013.04.043
Moreno G, Aviron S, Berg S, Crous-Duran J, Franca A, de Jalón SG, Hartel T, Mirck J, Pantera A, Palma JHN, Paulo JA, Re GA, Sanna F, Thenail C, Varga A, Viaud V, Burgess PJ (2018) Agroforestry systems of high nature and cultural value in Europe: provision of commercial goods and other ecosystem services. Agrofor Syst 92(4):877–891. https://doi.org/10.1007/s10457-017-0126-1
Moreno G, Pulido FJ (2009) The functioning, management and persistence of dehesas. In: A Rigueiro-Rodríguez J, McAdam J, MR Mosquera-Losada (eds) Agroforestry in Europe: current status and future prospects. Springer, Netherlands, vol. 6, p 127–160. ISBN:9781402082719
Morgan-Davies C, Waterhouse A, Pollock ML, Holland JP (2008) Integrating hill sheep production and newly established native woodland: achieving sustainability through multiple land use in Scotland. Int J Agr Sustain 6(2):133–147. https://doi.org/10.3763/ijas.2007.0248
Mosquera-Losada MR, McAdam JH, Romero-Franco R, Santiago-Freijanes JJ, Rigueiro-Rodríguez, A (2009) Definitions and components of agroforestry practices in Europe. In: A Rigueiro-Rodríguez J, McAdam J, MR Mosquera-Losada (eds) Agroforestry in Europe: current status and future prospects. Springer, Netherlands, Vol. 6, p 3–20. https://doi.org/10.1007/978-1-4020-8272-6_1. ISBN:9781402082719
Mupepele A-C, Keller M, Dormann CF (2021) European agroforestry has no unequivocal effect on biodiversity: a time-cumulative meta-analysis. BMC Ecol Evol 21(1):193. https://doi.org/10.1186/s12862-021-01911-9
Nair PR (2007) The coming of age of agroforestry. J Sci Food Agr 87(9):1613–1619. https://doi.org/10.1002/jsfa.2897
Nair PR, Mohan Kumar B, Nair VD (2009) Agroforestry as a strategy for carbon sequestration. J Plant Nutr Soil Sci 172(1):10–23. https://doi.org/10.1002/jpln.200800030
Nair PR (1993) An introduction to agroforestry. Kluwer, Dordrecht
Nerlich K, Graeff-Hönninger S, Claupein W (2013) Agroforestry in Europe: a review of the disappearance of traditional systems and development of modern agroforestry practices, with emphasis on experiences in Germany. Agrofor Syst 87(2):475–492. https://doi.org/10.1007/s10457-012-9560-2
Newton I (2004) The recent declines of farmland bird populations in Britain: an appraisal of causal factors and conservation actions: recent declines of farmland bird populations in Britain. Ibis 146(4):579–600. https://doi.org/10.1111/j.1474-919X.2004.00375.x
Oksanen J, Blanchet FG, Kindt R, Minchin PR, Hara RBO, Simpson GL, Sólymos P, Stevens MHH, Wagner H (2013) Vegan: Community Ecology Package. R Package Version 2.0-9. http://cran.r-project.org/package=vegan%20
Pain DJ, Pienkowski MW (1997) Farming and birds in Europe: the Common Agricultural Policy and its implications for bird conservation. Academic Press, UK. ISBN:9780125442800
Pardon P, Reheul D, Mertens J, Reubens B, De Frenne P, De Smedt P et al (2019) Gradients in abundance and diversity of ground dwelling arthropods as a function of distance to tree rows in temperate arable agroforestry systems. Agr Ecosyst Environ 270:114–128
Pickett SR, Siriwardena GM (2011) The relationship between multi-scale habitat heterogeneity and farmland bird abundance. Ecography 34(6):955–969
Pijanowski BC, Villanueva-Rivera LJ, Dumyahn SL, Farina A, Krause BL, Napoletano BM, Gage SH, Pieretti N (2011) Soundscape ecology: the science of sound in the landscape. BioScience 61(3):203–216. https://doi.org/10.1525/bio.2011.61.3.6
Poulsen BO (2002) Avian richness and abundance in temperate Danish forests: tree variables important to birds and their conservation. Biodivers Conserv 11:1551–1566. https://doi.org/10.1023/A:1016839518172
Quinkenstein A, Woellecke J, Böhm C, Grünewald H, Freese D, Schneider BU, Hüttl RF (2009) Ecological benefits of the alley cropping agroforestry system in sensitive regions of Europe. Environ Sci Policy 12(8):1112–1121. https://doi.org/10.1016/j.envsci.2009.08.008
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Reif J, Hanzelka J (2020) Continent-wide gradients in open-habitat insectivorous bird declines track spatial patterns in agricultural intensity across Europe. Global Ecol Biogeogr 29(11):1988–2013. https://doi.org/10.1111/geb.13170
Reisner Y, de Filippi R, Herzog F, Palma J (2007) Target regions for silvoarable agroforestry in Europe. Ecol Eng 29:401–418. https://doi.org/10.1016/j.ecoleng.2006.09.020
Robinson RA, Sutherland WJ (2002) Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 39(1):157–176. https://doi.org/10.1046/j.1365-2664.2002.00695.x
Rösch V, Hafner G, Reiff JM, Entling MH (2023) Increase in breeding bird abundance and diversity with semi-natural habitat in vineyard landscapes. PloS ONE 18:e0284254. https://doi.org/10.1371/journal.pone.0284254
Schulze ED, Craven D, Durso AM, Reif J, Guderle M, Kroiher F, Hennig P, Weiserbs A, Schall P, Ammer C, Eisenhauer N (2019) Positive association between forest management, environmental change, and forest bird abundance. For Ecosyst 6(1):3. https://doi.org/10.1186/s40663-019-0160-8
Söderström B, Pärt T (2000) Influence of landscape scale on farmland birds breeding in semi-natural pastures. Conserv Biol 14(2):522–533. https://doi.org/10.1046/j.1523-1739.2000.98564.x
Söderström B, Svensson B, Vessby K, Glimskär A (2001) Plants, insects and birds in semi-natural pastures in relation to local habitat and landscape factors. Biodivers Cons 10:1839–1863. https://doi.org/10.1023/A:1013153427422
Stoate C, Boatman ND, Borralho RJ, Carvalho CR, Snoo GR, Eden P (2001) Ecological impacts of arable intensification in Europe. J Environ Manage 63(4):337–365. https://doi.org/10.1006/jema.2001.0473
Tellería JL (2001) Passerine bird communities of Iberian dehesas: a review. Anim Biodiv Conserv 24(2):67–78
Tilman D, Balzer C, Hill J, Befort BL (2011) Global food demand and the sustainable intensification of agriculture. P Natl A Sci USA 108(50):20260–20264. https://doi.org/10.1073/pnas.1116437108
Torralba M, Fagerholm N, Burgess PJ, Moreno G, Plieninger T (2016) Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agr Ecosyst Environ 230:150–161. https://doi.org/10.1016/j.agee.2016.06.002
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity – ecosystem service management. Ecol Lett 8(8):857–874. https://doi.org/10.1111/j.1461-0248.2005.00782.x
Voříšek P, Jiguet F, van Strien A, Škorpilová J, Klvaňová A, Gregory RD (2010) Trends in abundance and bio-mass of widespread European farmland birds: how much have we lost? BOUProceedings–Lowland Farmland Birds III. http://www.bou.org.uk/bouproc-net/ccb/vorisek-etal.pdf Accessed 8 September 2023
Yip DA, Leston L, Bayne EM, Sólymos P, Grover A (2017) Experimentally derived detection distances from audio recordings and human observers enable integrated analysis of point count data. Avian Conserv Ecol 12(1):art11. https://doi.org/10.5751/ACE-00997-120111
Acknowledgements
We would like to thank our colleagues from the AGROMIX project (Mickael Bernard, María Blanco Gutiérrez, Samuele Ciacci, Lydie Dufour, Marco Gentili, Samuel Hepner, Gerardo Moreno, João Palma, Alice Ripamonti, Robin Russias, Will Simonson, Jo Smith, Sophie Tournier, and Lorenzo Gabriele Tramacere) who helped with data collection as well as the land owners for allowing us to sample their farms. Thanks to the AGROMIX colleagues Felix Herzog, Rachel Ramsey, Rodrigo Olave, and Katharina Dehnen-Schmutz for the discussions about the study concept. A special thanks to Fabian Bindrich and Alexandre Leitão for sharing their expertise on bird identification. Finally, we would like to thank Felix Herzog and Jo Smith for their manuscript proofreading as well as the Editors and Reviewers for their valuable comments on earlier versions of the manuscript.
Funding
Open Access funding enabled and organized by Projekt DEAL. This research was part of the AGROMIX Project and received funding from the European Union’s Horizon 2020 research and innovation program under grant agreement 862993.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by M. E. The first draft of the manuscript was written by M. E. and all authors commented on previous versions of the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval
Not applicable
Consent to participate and for publication
Not applicable
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Edo, M., Entling, M.H. & Rösch, V. Agroforestry supports high bird diversity in European farmland. Agron. Sustain. Dev. 44, 1 (2024). https://doi.org/10.1007/s13593-023-00936-2
Accepted:
Published:
DOI: https://doi.org/10.1007/s13593-023-00936-2