
Abstract
The intensive use of synthetic nitrogen fertilizers over the last century has both increased agricultural productivity and modified biogeochemical cycles in terrestrial ecosystems, causing severe negative environmental impacts. Tea (Camellia sinensis L.) plantations usually receive high levels of synthetic fertilizers, which strongly affect plant and soil properties. However, there is no quantitative study to assess how synthetic N additions affect soil chemistry and the production and quality of tea shoots. Here, we conducted a comprehensive meta-analysis to evaluate the effects of experimental synthetic N fertilizers. Our main findings are (1) N additions in tea plantations acidify soils (− 0.41 pH unit in average) and produce soil nutrient imbalance. Soil acidification commonly exacerbates the accumulations of toxic aluminum ions. (2) Synthetic N fertilizer additions may strongly increase tea production by almost 70% but alter tea shoot quality by increasing the concentrations of free amino acids (+ 16%), caffeine (+ 14%), and water extracts (+ 5%) while decreasing those of soluble sugars (− 8%) in the tea shoots. The responses of soil chemistry, tea production, and quality to N additions can vary among experimental conditions, tea tree species, and N fertilizer forms. Because there is statistical limitation in this meta-analysis, our findings recommend performing additional field studies to explore the potential mechanisms of nutrient cycling and ecosystem functioning under synthetic N additions. The development of a sustainable N management strategy in tea plantations is also urgently needed to enhance N use efficiency and reduce environmental risks.
Similar content being viewed by others
1 Introduction
Tea (Camellia sinensis L.) is the most consumed manufactured drink worldwide and is widely planted in tropical and subtropical areas, especially in the corresponding areas of Asian, African, and Latin American countries (Chang 2015; Hilal 2015). As a leaf-plucked cash crop, tea trees require a high nitrogen (N) supply; the current levels of N inputs in tea plantations range from 450 to 1200 kg N ha−1 year−1 (Tokuda and Hayatu 2001; Tokuda and Hayatsu 2004; Li et al. 2013), which are much higher than those in other artificial ecosystems. Synthetic N additions have been proposed to alter soil chemical properties as well as tea production and quality (Owuor and Cheruiyot 1989; Venkatesan et al. 2004; Ruan et al. 2010). Concurrently, massive amounts of N not taken up by plants tend to be lost to adjacent systems, causing environmental problems in tea-planted areas (Tokuda and Hayatsu 2004; Zhu et al. 2014).
Wide applications of synthetic N fertilizers can cause soil acidification and the depletion of base metal cations in terrestrial ecosystems (Bolan et al. 1991; Barak et al. 1997; Bowman and Cleveland 2008). In most tea-planted areas, soils are highly acidic and well drained (Ruan et al. 2013). Evidence suggests that acidic soils would experience stronger acidification under excessive N additions (Alekseeva et al. 2011). The losses of soil base cations (e.g., Ca2+ and Mg2+) and the accumulation of toxic aluminum (Al3+) under N fertilization might reduce the activity of plant roots and inhibit the growth of tea trees (Ruan 2005). However, a systematic understanding of how synthetic N additions affect the chemistry of tea-planted soils and how these changes impact tea growth and tea quality formation are still lacking.
Global tea production reached approximately 5064 thousand tons in 2013, and tea consumption has also rapidly increased, particularly in China (Fig. 1a shows the green tea planting area in the southern region of Henan province) and India, among other countries (Chang 2015). The global black and green tea productions are projected to grow at 2.90 and 8.20%, respectively, by 2023 (Chang 2015; Hilal 2015). Tea bioactive ingredients are widely considered to have human health benefits, such as controlling blood pressure and glucose levels (Hodgson et al. 2013; Zheng et al. 2013; Weerawatanakorn et al. 2015), promoting fat oxidation for weight management (Dulloo et al. 1999) and causing relaxation effects (Juneja et al. 1999). An unbalanced soil nutrient status can limit plant growth and biochemical ingredient formation (Li and Xia 2005). N fertilizer addition has been shown to increase tea shoot yield by enhancing of soil N availability, while excessive N application may decrease tea quality. Free amino acids and polyphenols are key metabolites in plucked shoots, and the polyphenol/free amino acid ratio (PP/AA ratio) is generally used to evaluate the quality of prepared tea and the taste of tea infusions (Wang et al. 2010). Increasing the N supply enhances the formation and accumulation of free amino acids in plucked tea shoots (Okano et al. 1997; Ruan et al. 2007) while increasing or decreasing the polyphenol concentration (Venkatesan and Ganapathy 2004; Mudau et al. 2006), resulting in varying PP/AA ratios. Synthetic N additions might play an important role in modifying the biochemical properties of tea shoots and affecting the quality of specific manufactured tea products (Ruan 2005).

a The green tea plantations in mountain area of South Henan province, China (Photo credit Linxiang Jiang, Xinyang normal university). b Global distribution of the study sites in this meta-analysis. Green dots: green tea; red dots: black tea; light-green triangles: oolong tea; black rhombus: dark tea; gray rhombus: white tea. Black pins: N. A. (unavailable information)
Many simulated N-addition studies have been conducted to investigate the responses of soils, plants, or the N cycle in tea plantations. However, a knowledge gap still exists with respect to understanding the effects of synthetic N additions on tea-planted ecosystems. The objectives of this meta-analysis, therefore, were (1) to comprehensively and quantitatively evaluate the effects of experimental synthetic N additions on the chemistry of tea-planted soils and the yield and quality of tea shoots and (2) to explore how N fertilizer effects varied among different experimental and environmental conditions.
2 Methods
2.1 Data compilation
In this meta-analysis, the relevant peer-reviewed articles were retrieved using the Web of Sciences (1973–2016) and Google Scholar. The analyses were restricted to experiments meeting the following criteria: (1) field experiments conducted in tea plantations or tea tree-planted pot experiments, (2) minimum of two levels of synthetic N fertilizer additions, and (3) corresponding variables measured in the experiment. The effects of N additions on tea plantation ecosystems were assessed using two categories with 18 variables: (1) soil chemistry: soil pH; soil available phosphorus (P); concentrations of potassium ion (K+), calcium ion (Ca2+), magnesium ion (Mg2+), and aluminum ion (Al3+); soil organic carbon (SOC); total N (TN); soil dissolved inorganic nitrogen (DIN); ammonium (NH4+) concentration; and nitrate (NO3−) concentration and (2) the yield and quality of tea shoots: shoot yield, free amino acids, polyphenols, the polyphenol/free amino acid ratio (PP:AA ratio), caffeine, soluble sugars, and total water extracts.
The results of the first layer of soil sampling (mostly between 0 and 20 cm) were used to assess the effects of N addition on the chemistry of tea-planted soil. Tea production in our database was mainly presented as the fresh weight of tea shoots. In most N-addition experiments in our database, the quality properties were determined by the chemical analysis of dry tea shoot material that has not undergone tea manufacturing procedures. For different characteristics, plucked shoots from different tea tree species may be suitable for processing into only one type of tea (Sedaghathoor et al. 2009). We therefore used tea types referenced in the literature to define tea tree species, e.g., tea trees were categorized as oolong tea species if they were referenced as being suitable for processing into oolong tea. In addition, N fertilizer levels, experimental locations, tea tree species, and latitude were also included in the dataset. In the data source studies included in our database, all controls used had no N applied (the N amount was 0), except for the study of Hilton et al. 1973 (control N level: 40.48 kg ha−1, the net N amount of the treatment was used); therefore, we used “N addition/application levels” in the following text to represent the amounts of N addition in treatments compared to those in the controls. If data were presented only in graphical form, we quantified it by using Engauge Digitizer software (Free Software Foundation, Inc., Boston, MA, USA).
2.2 Meta-analysis
In total, information from 25 peer-reviewed publications and 464 datasets was extracted for the meta-analysis. Please find details of the database for the meta-analysis via doi: https://doi.org/10.5281/zenodo.1049182. The locations of all experimental sites are shown in Fig. 1b. For soil pH, SOC, and TN parameters, the average values across the entire tea growing season were used. The effect sizes of synthetic N additions on the parameters were assessed by different experimental conditions, N fertilizer forms, and tea tree species (black, green, oolong, white and dark tea trees) in corresponding studies. Observations were considered independent if the data were obtained from separate studies or from different treatments within a single experiment, e.g., treatments with more than N level, N form, or tea tree species were reported in a study (Liu and Greaver 2009; Qiao et al. 2015). If data were reported in a study for multiple experimental years under the same conditions, we used the average level across years as the independent observations.
The response ratio (RR) was used to estimate the effect of N fertilizer application on each parameter (Hedges et al. 1999). For each parameter, the natural log of RR was calculated by lnRR = ln(XT/XC), where XC is the control mean and XT is the treatment mean. The average RR was calculated using a mixed model and MetaWin 2.1 (Sinauer Associates, Inc. Sunderland, MA, USA). We also assessed the soil and tea responses to N additions under different experimental conditions and with different tea tree species and N forms. Due to the limited information available, the responses of soil chemistry were mainly discussed with respect to different experimental conditions. The total heterogeneity among categorical groups (Qt) was partitioned into within-group heterogeneity (Qw) and between-group heterogeneity (Qb). When the P value of Qb was lower than 0.05, the RRs were considered significantly different among subgroups. The effects of N addition on each parameter were considered significant if the percent change in the RR did not overlap 0.
3 Results and discussion
3.1 Effects of N additions on soil chemistry
3.1.1 Soil carbon and N
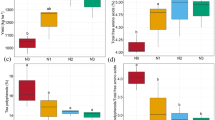
Anthropogenic N inputs can dramatically affect the biogeochemical cycles of carbon (C) and N in terrestrial ecosystems. Although detectable changes in C storage have rarely been reported before (Nadelhoffer et al. 1999; Giardina et al. 2004; Mack et al. 2004), currently, N additions can increase the SOC storage in terrestrial ecosystems (Pregitzer et al. 2008; Liu and Greaver 2010; Fornara and Tiliman 2012; Zak et al. 2017). In agricultural ecosystems, N additions have significantly increased the SOC by 3.5% (Lu et al. 2011). Tea plantation ecosystems make an important contribution to C sinks in tea planting regions (Zhang et al. 2017). However, in the present study, N additions showed no significant effects on the SOC of tea plantations (Fig. 2). For different tea-planted soils, N additions significantly increased the SOC of green tea-planted soils (mean + 15.13%, bootstrap CI + 6.91 to + 23.99%) but did not affect the SOC of oolong tea tree-planted soils. Most green tea products are made only from young tea shoots and leaves. After young shoots are plucked, the canopy is cut; more massive pruning materials under N fertilization would be returned back to soil rather than being processed into products. We thought this might be the main reason for the significant SOC increase in green tea-planted soils under N-addition conditions.

Impacts of synthetic N fertilizer additions on the chemistry of soils in which tea trees were planted. SOC soil organic carbon, DIN dissolved inorganic nitrogen. Values are the means ± bootstrap CIs. Numbers in parentheses are the sample sizes of variables
Synthetic N additions increased the soil TN (+ 18.29, + 10.22 to + 26.66%, Fig. 2). When the data between experimental conditions were examined, N additions increased the soil TN of studies conducted under field conditions (Table 1) but did not affect that in pot studies. However, the findings in pot studies tend to over- or underestimate the effects of treatments due to the unnatural environment and relatively short experimental period. Further, the relatively small sample size might also limit the power of the results from pot studies. On average, N fertilizer additions largely increased the soil DIN concentration in tea plantations (+ 73.94, + 36.22 to + 139.68%, Fig. 2). When the data were subdivided by experimental conditions, N additions significantly increased the soil DIN in field studies but did not affect that in the pots. N additions consistently increased the soil DIN of green and oolong tea-planted soils. It has been demonstrated that tea plants prefer ammonium nutrition and are well-adapted to NH4+-rich environments (Ruan et al. 2007). N additions significantly increased soil NH4+ concentrations (+ 103.25, + 41.65 to + 212.01%, Fig. 2) when averaged across all studies, and the effects were significant among all subgroups (Table 1). N additions also clearly increased the soil NO3− concentration (+ 177.78, + 86.28 to + 308.91%, Fig. 2). Differences in experimental conditions and planted tea tree species altered the responses of soil NO3− (Tables 1). Due to the existence of the nitrification process (Li et al. 2013), the NO3− increase in tea-planted soils in response to N fertilizer was greater than the NH4+ increase (Fig. 2); this phenomenon poses a high risk of soluble reactive N losses to underground and surface water systems.
3.1.2 Soil acidification, aluminum toxicity, and nutrient imbalance
Acidic soils with a pH below 5.5 limit agricultural production (Kochian et al. 2004). Nevertheless, the optimal pH for the growth of tea tree, an acid-tolerant plant, is 4.5 to 6.0, with 5.5 being the most suitable pH (USEPA 2008). The acidification of tea-planted soils, affected by various environmental and anthropogenic factors, has threatened the sustainability of tea-planting areas (Li et al. 2016). Synthetic N fertilizer additions significantly reduced the pH of tea-planted soils by 0.45 (− 8.74, − 10.49 to − 7.03%, Fig. 2) when averaged across all studies, leading to an extremely acidified environment for tea tree growth. The 0.45 unit reduction in tea-planted soil pH is similar to the reduction of 0.50 reported for Chinese agricultural systems (Guo et al. 2010) but is higher than the reported 0.26 in natural terrestrial ecosystems (Tian and Niu 2015). The high acidification of tea-planted soils under N additions can reduce the growth of tea plants (Ruan et al. 2007) and potentially damage the ecosystem health of tea plantations. The negative responses of soil pH to N additions in tea plantations were consistent among the various experimental conditions and tea trees (Table 1. Furthermore, the decrease in soil pH was inversely related to initial soil pH, find by doi: https://doi.org/10.5281/zenodo.1049182, the same below) and N addition level. The acidification of soils under N fertilization was a gradual process. In the early stage of N fertilization, soils tend to lose massive amounts of base cations by leaching, weakening the soil buffering capacity and leading to acidification. During long-term N inputs, neutral soils with a relatively high initial pH would experience a relative larger extent of decrease in pH before reaching the lowest threshold of soils that had a relatively lower initial pH. In addition, the extent of soil acidification was also determined by the levels of N fertilizer inputs. These results would provide some insights into land use changes in tropical and subtropical mountain areas. Forest ecosystems with neutral soil pH might suffer sharp and severe soil acidification when being transformed into tea plantations with excessive N inputs.
The soil base cations K+, Mg2+, and Ca2+ play great roles in buffering against soil acidification during early storage (Tachibana et al. 1995; Bowman and Cleveland 2008; Tian and Niu 2015). Indeed, we found that synthetic N additions significantly depleted soil K+ by 18.20% (− 32.07 to − 1.49%), Ca2+ by 23.28% (− 30.10 to − 16.00%), and Mg2+ by 37.11% (− 48.92 to −24.99%, Fig. 2). Soils of forest and grassland ecosystems worldwide undergo buffering transitions of base cations to non-base cations (Tian and Niu 2015). Soil Al3+ plays a critical role in maintaining the buffering capacity of soil systems when the soil pH decreases below 4.5 (Bowman and Cleveland 2008, Li et al. 2013). In addition, soil acidification can exacerbate aluminum toxicity effects on plants (Dong et al. 1999). In the present study, in acidified tea-planted soils, soil Al3+ accumulation dramatically increased by 73.73% (+ 56.23 to + 93.63%, Fig. 2) under N additions. The Al3+ response in black tea tree-planted soils was greater than that in green tea-planted soils (Qb = 0.31, P < 0.05, The planting regions of black tea trees were generally at lower latitudes than were those of green tea trees. In most areas, the soils were deeply weathered and retained higher levels of Al3+. The stronger soil acidification under N addition might accelerate Al3+ release to maintain the buffering capacity (Dong et al. 1999). The bioavailability of soil Al3+ can affect Al concentrations in most plant organs (Fang et al. 2014). Tea trees are typically aluminum-tolerant plants; they have physiological mechanisms to detoxify Al3+ internally by secreting oxalic acid or by altering either the activity of enzymes in the leaf shoots or the accumulation of Al3+ in the cell walls of old leaves (Carr et al. 2003, Yu et al. 2012). Although soil Al3+ accumulation can stimulate tea tree growth (Carr et al. 2003), the limitations of toxic Al3+ on plant growth and its inhibitory effects on tea tree growth always occur (Ma et al. 2001, Ruan 2005, Zheng 2010). Since tea is a major dietary source of Al for humans (De Silva et al. 2016), it is necessary to conduct future studies focused on the accumulation and transfer of Al in soil-plant systems under N fertilization.
Long-term tea planting causes nutrient imbalance in soil-plant systems (Dang 2005). It has been demonstrated that N additions can greatly affect soil P availability in terrestrial ecosystems. A meta-analysis showed that N enrichment could aggravate P limitations on plant biomass production in various terrestrial ecosystems (Li et al. 2016). However, our result showed that N fertilizer additions did not impact the soil available P supply in tea plantations. We believed that, in most tea-planted areas, the soils are heavily weathered and severely lack available P. Therefore, small-magnitude soil P changes under N addition may not have been easily detected. In addition, the sample size of soil P (n = 4) would also limit the power of this result. The base cations K+, Mg2+, and Ca2+ are also vital mineral nutrients for tea tree growth (Sedaghathoor et al. 2009). Potassium is an important nutrient for tea (Ranganatha and Natesan 1985), and our result indicated that N additions led to deficient K+ supplies (− 18.20, − 1.49 to − 32.07%) in the tea-planted soils. The smaller K+ availability could also affect tea yield as well as the biochemical properties and leaf nutrient status of tea plants (Ruan et al. 1999; Venkatesan and Ganapathy 2004).
Soil acidification caused by N fertilization exacerbates the severity of aluminum toxicity and nutrient imbalance for tea plants and threatens tea production and quality. To alleviate the impairment of ecosystem services in tea-planted areas, further field research should be conducted to explore the underlying mechanisms and effective nutrient management strategies in tea plantations. Furthermore, intense public attention and effective governmental policies are also vital for developing sustainable tea planting ecosystems.
3.2 Effects of N additions on tea yield and quality
3.2.1 Tea yield
Nitrogen additions induced a significant increase in tea yield across all studies (+ 68.74, + 53.98 to + 85.72%, Fig. 3). The impact of N addition varied among tea tree species and experimental conditions but not among N forms. The increase in tea yield in response to N addition in pot studies was higher than that in field studies (Qb = 0.96, P < 0.01). Compared with black tea trees, green tea trees planted in the soil under N fertilization exhibited larger increases in tea yield (Qb = 1.10, P < 0.05). Nitrogen is a vital essential element for plant growth, and adequate N fertilizer can promote plant growth and production. As an NH4+-preferring plant, the absorption of NH4+ by tea trees was faster than that of NO3− during the cultivation process (Hoshina 1985; Morita et al. 1998; Ruan et al. 2007). With more than twofold increases in soil NH4+ (+ 103.25%, Fig. 2) and tea shoot N (+127.62%, n = 24, unpublished data), the positive responses of tea yield might be due mainly to the increased soil N availability and the enhanced NH4+-N uptake from soils. The increase in tea yield was expected to be higher under conditions of NH4+ addition than under those of NO3− addition; however, the statistical result of N form on tea yield was not significant in this study

Impacts of synthetic N additions on tea shoot yield and the quality properties of tea shoots. SSs soluble sugars, WEs water extracts. Values are the means ± bootstrap CIs. Numbers in brackets are the sample sizes of variables
3.2.2 Tea quality
On average, N addition greatly increased the free amino acid concentration in tea shoots (+ 16.46, + 10.10 to + 24.08%, Fig. 3). This increase in free amino acids was found to be proportional to the amount of N fertilizer applied (R2 = 0.39, P < 0.01,). The impact of N additions on tea free amino acids was not affected by experimental conditions or tea tree species. However, the effects were affected by the chemical forms of N applied. The NH4+-type fertilizers, not NO3− or NH4NO3, significantly increased the free amino acids in tea shoots (Qb = 0.14, P < 0.05,); this increase might be attributed to the preference for NH4+ and the fast incorporation of absorbed NH4+ into amino acids in tea plants (Ruan et al. 2007; OH et al. 2008). The overall response of tea shoot polyphenol concentrations was not significant (Fig. 3). However, N additions significantly decreased the PP/AA ratio across all studies (− 7.99, − 14.00 to − 1.92%, Fig. 3). The response of the PP/AA ratio consisted of a nonlinear decrease as the N addition level increased (R2 = 0.30, P < 0.05). The responses of the shoot PP/AA ratio differed among different tea tree species and N forms (Table 2). Synthetic N additions affected tea quality by altering the relative content of chemical constituents. Shoot free amino acid contents can affect the taste of prepared tea, especially the slightly sweet taste of green tea products (Lvova et al. 2003). The current result indicated that the quality of manufactured tea products might be improved by concentrating more free amino acids. Tea polyphenols and their oxidized derivatives are responsible for the color and unique aroma of tea infusions (Ruan 2005). The relative decrease in the PP/AA ratio of tea shoots induced by N additions might also alter the oxidization and fermentation during the manufacturing process and alter the quality of tea products.
Our results indicated that N additions significantly increased the caffeine concentration in tea shoots (+ 14.00, + 10.68 to + 17.09%, Fig. 3), and the responses were almost significant within subgroups (Table 2). When the data were divided by tea tree species, the increase in caffeine of dark tea tree species was significantly higher than that of black and green tea tree species (Qb = 0.09, P < 0.01). The chemical N form also affected the responses of shoot caffeine concentrations (Qb = 0.06, P < 0.05,). In general, N additions significantly reduced the concentration of soluble sugars in tea shoots (− 7.83, − 13.67 to − 1.60%, Fig. 3). Reductions in soluble sugar concentrations under N fertilization have also been reported in other plants, e.g., birch, apple trees, and rice (Lavola and Julkunen-Tiitto 1994; Cheng et al. 2004; Wang et al. 2006). N supply can regulate plant photosynthesis and product formation. As N supplies increase, more C in the leaves would be incorporated into amino acids and proteins rather than stored within carbohydrates (Cheng et al. 2004). The responses of soluble sugars to N addition were affected by experimental conditions, tea tree species, and N form (Table 2). Nitrogen fertilizers increased the water extracts of tea shoots across all studies, with a proportion of 5.35% (+ 3.67 to + 6.97%, Fig. 3); however, the responses were not affected by tea tree species (Qb = 0.01, P = 0.14,).
3.3 Meta-analysis limitations
By conducting a meta-analysis, we quantitatively assessed the impact of synthetic N fertilizer additions on the chemistry of tea-planted soils and on the production and quality of tea shoots. However, flaws in the data still existed in the meta-analysis. Findings in pot studies tended to over- or underestimate the effects of experimental N additions. In our dataset, the results of pot studies dominated the overall results, which might affect the accuracy of the results. Therefore, we suggest that further studies focusing on the soil nutrient cycle and balance of tea plantations should be carried out in field. Furthermore, the small study and sample sizes of variables (e.g., soil available P) might limit the statistical power of the results. Due to the lack of standard deviations or standard errors of experimental and control groups, we could not weigh RRs by effect sizes and could not assess the publication bias of each variable based on our current database. In the meta-analysis, observations within a single experiment were considered independent if they were from treatments with differentially manipulated levels (Koricheva et al. 2013), such as N levels, N forms, tea tree species, and so on. If the data sources for a meta-analysis are sufficient, we recommend that data from multiple treatments within a single experiment sharing a common control should be used, e.g., by using their average values, to keep datasets fully independent. Although we adequately considered the controlled factors, including the type of fertilizer, N application level, tea tree age, soil sampling time, and shoot harvest time, we could not fully assess how these factors affected the effects of synthetic N on tea-planted soils and on tea plants for the limited studies conducted in tea plantations.
4 Conclusions
Tea has high socio-economic importance in areas where it is planted. Our results indicated that, by reducing the soil pH and altering the composition of the soil buffering system, synthetic N fertilizers could significantly alter the chemistry of tea-planted soils, leading to soil nutrient imbalance. Experimental N additions could significantly increase tea production and alter tea biochemical constituents. The responses of soil chemistry and tea production and quality to N additions varied under different experimental conditions and with different tea tree species and N forms.
Given the widespread planting of tea plants and the worldwide popularity of tea drinking, it is critically important to conduct field research to explore the underlying mechanisms of nutrient cycling and ecosystem function under N addition and to improve N fertilizer management strategies in tea plantations. In future field studies, the manipulative and sampling variability, such as fertilization method, soil sampling depth, and tea sampling season and stage, should be fully considered. Optimal N application and scientific management could cause economic and ecological benefits for tea-planted areas and could further substantially benefit the health and welfare of human beings. Well-managed combinations of chemical and organic fertilizers could also overcome the disadvantages of single-source fertilizer applications (Miao et al. 2010). Therefore, another area that might benefit from future N-addition research in tea plantations involves the interactive effects of N chemical forms (or N gradients, etc.) and effective methods, including combined applications of nitrification inhibitors (Akiyama et al. 2010; Abalos et al. 2014; Qiao et al. 2015) and organic fertilizers. Improving nitrogen-use efficiency during crop production is critical for addressing the triple challenges of food security, environmental degradation, and climate change (Zhang et al. 2015). The exploration of sustainable N-management strategies with various affecting factors is particularly urgently needed to improve N efficiency and reduce environmental risks in tea plantations.
Abbreviations
- Al3+ :
-
Aluminum ion
- Ca2+ :
-
Calcium ion
- DIN:
-
Soil dissolved inorganic nitrogen
- K+ :
-
Potassium ion
- Mg2+ :
-
Magnesium ion
- NH4 + :
-
Ammonium
- NO3 − :
-
Nitrate
- P:
-
Phosphorus
- PP/AA ratio:
-
Polyphenol/free amino acid ratio
- Q t :
-
Total heterogeneity among categorical group
- Q w :
-
Within-group heterogeneity
- Q b :
-
Between-group heterogeneity
- RR:
-
Response ratio
- SOC:
-
Soil organic carbon
- TN:
-
Total N
References
Abalos D, Jeffery S, Sanz-Cobena A, Guardia G, Vallejo A (2014) Meta-analysis of the effect of urease and nitrification inhibitors on crop productivity and nitrogen use efficiency. Agr Ecosyst Environ 189(0):136–144. https://doi.org/10.1016/j.agee.2014.03.036
Akiyama H, Yan X, Yagi K (2010) Evaluation of effectiveness of enhanced-efficiency fertilizers as mitigation options for N2O and NO emissions from agricultural soils: meta-analysis. Glob Chang Biol 16(6):1837–1846. https://doi.org/10.1111/j.1365-2486.2009.02031.x
Alekseeva T, Alekseev A, RK X, Zhao AZ, Kalinin P (2011) Effect of soil acidification induced by a tea plantation on chemical and mineralogical properties of Alfisols in eastern China. Environ Geochem Health 33(2):137–148. https://doi.org/10.1007/s10653-010-9327-5
Barak P, Jobe BO, Krueger AR, Peterson LA, Laird DA (1997) Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant Soil 197(1):61–69. https://doi.org/10.1023/A:1004297607070
Bolan NS, Hedley MJ, White RE (1991) Processes of soil acidification during nitrogen cycling with emphasis on legume based pastures. Plant Soil 134(1):53–63. https://doi.org/10.1007/BF00010717
Bowman WD, Cleveland CC (2008) Negative impact of nitrogen deposition on soil buffering capacity. Nat Geosci 1(11):767–770. https://doi.org/10.1038/ngeo339
Carr H, Lombi E, Küpper H, Mcgrath S, Wong M (2003) Accumulation and distribution of aluminium and other elements in tea (Camellia sinensis) leaves. Agronomie 23(8):705–710. https://doi.org/10.1051/agro:2003045
Chang K (2015) World tea production and trade: current and future development. A publication by the Food and Agricultural Organization of the United Nations, Rome. Available online at www fao org. Accessed 29 May 2016
Cheng L, Ma F, Ranwala D (2004) Nitrogen storage and its interaction with carbohydrates of young apple trees in response to nitrogen supply. Tree Physiol 24:91–98
Dang MV (2005) Soil-plant nutrient balance of tea crops in the northern mountainous region, Vietnam. Agric Ecosyst Environ 1(105):413–418. https://doi.org/10.1016/j.agee.2004.05.004
De Silva J, Tuwei G, Zhao FJ (2016) Environmental factors influencing aluminium accumulation in tea (Camellia sinensis L.) Plant Soil 400(1):223–230. https://doi.org/10.1007/s11104-015-2729-5
Dong D, Xie Z, Du Y, Liu C, Wang S (1999) Influence of soil ph on aluminum availability in the soil and aluminum in tea leaves. Commun Soil Sci Plan 30(5–6):873–883. https://doi.org/10.1080/00103629909370252
Dulloo AG, Duret C, Rohrer D, Girardier L, Mensi N, Fathi M, Chantre P, Vandermander J (1999) Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. Am J Clin Nutr 70(6):1040–1045
Fang X, Chen F, Hu X, Yuan P, Li J, Chen X (2014) Aluminum and nutrient interplay across an age-chronosequence of tea plantations within a hilly red soil farm of subtropical China. Soil Sci Plant Nutr 60(4):448–459. https://doi.org/10.1080/00380768.2014.912950
Fornara DA, Tilman D (2012) Soil carbon sequestration in prairie grasslands increased by chronic nitrogen addition. Ecology 93(9):2030–2036. https://doi.org/10.1890/12-0292.1
Giardina C, Binkley D, Ryan M, Fownes J, Senock R (2004) Belowground carbon cycling in a humid tropical forest decreases with fertilization. Oecologia 139(4):545–550. https://doi.org/10.1007/s00442-004-1552-0
Guo J, Liu X, Zhang Y, Shen J, Han W, Zhang W, Christie P, Goulding K, Vitousek P, Zhang F (2010) Significant acidification in major Chinese croplands. Science 327(5968):1008–1010. https://doi.org/10.1126/science.1182570
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecology 80(4):1150–1156.
Hilal MM (2015) The dynamics of production. Consumption and Prices: A Study on Global Tea Industry Volume IV 99
Hilton P, Palmer-Jones R, Ellis R (1973) Effects of season and nitrogen fertiliser upon the flavanol composition and tea making quality of fresh shoots of tea (Camellia sinensis L.) in central Africa. J Sci Food Agr 24(7):819–826. https://doi.org/10.1002/jsfa.2740240710
Hodgson JM, Woodman RJ, Puddey IB, Mulder T, Fuchs D, Croft KD (2013) Short-term effects of polyphenol-rich black tea on blood pressure in men and women. Food Funct 4(1):111–115. https://doi.org/10.1039/C2FO30186E
Hoshina T (1985) Studies on absorption and utilization of fertilizer nitrogen by tea plants. BullNatl Res Inst Tea
Juneja LR, Chu DC, Okubo T, Nagato Y, Yokogoshi H (1999) L-theanine—a unique amino acid of green tea and its relaxation effect in humans. Trends Food Sci Tech 10(6-7):199–204. https://doi.org/10.1016/S0924-2244(99)00044-8
Kochian LV, Hoekenga OA, Pineros MA (2004) How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu Rev Plant Biol 55(1):459–493. https://doi.org/10.1146/annurev.arplant.55.031903.141655
Koricheva J, Gurevitch J, Mengersen K (2013) Handbook of meta-analysis in ecology and evolution. Princeton University Press. https://doi.org/10.1515/9781400846184
Lavola A, Julkunen-Tiitto R (1994) The effect of elevated carbon dioxide and fertilization on primary and secondary metabolites in birch, Betula pendula (Roth). Oecologia 99(3):315–321. https://doi.org/10.1007/bf00627744
Li J, Xia J (2005) Summary on nitrogen (N), phosphorus (P), potassium (K) and tea quality. Chinese Agricul Sci Bull 21:62–65
Li SX, Wang ZH, Stewart BA (2013) Responses of crop plants to ammonium and nitrate N. In: Donald L (ed) Adv Agron. Academic Press, New York. https://doi.org/10.1016/B978-0-12-405942-9.00005-0
Liu LL, Greaver TL (2009) A review of nitrogen enrichment effects on three biogenic GHGs: the CO2 sink may be largely offset by stimulated N2O and CH4 emission. Ecol Lett 12(10):1103–1117. https://doi.org/10.1111/j.1461-0248.2009.01351.x
Liu LL, Greaver TL (2010) A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol Lett 13(7):819–828. https://doi.org/10.1111/j.1461-0248.2010.01482.x
Lu M, Zhou X, Luo Y, Yang Y, Fang C, Chen J, Li B (2011) Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis. Agric Ecosyst Environ 140(1–2):234–244. https://doi.org/10.1016/j.agee.2010.12.010
Li S, Li H, Yang C, Wang Y, Xue H, Niu Y (2016) Rates of soil acidification in tea plantations and possible causes. Agr Ecost Environ 233:60–66. https://doi.org/10.1016/j.agee.2016.08.036
Lvova L, Legin A, Vlasov Y, Cha GS, Nam H (2003) Multicomponent analysis of Korean green tea by means of disposable all-solid-state potentiometric electronic tongue microsystem. Sensors Actuat B: Chem 95(1-3):391–399. https://doi.org/10.1016/S0925-4005(03)00445-3
Ma JF, Ryan PR, Delhaize E (2001) Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci 6(6):273–278. https://doi.org/10.1016/S1360-1385(01)01961-6
Mack MC, Schuur EAG, Bret-Harte MS, Shaver GR, Chapin Iii FS (2004) Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 431(7007):440–443. https://doi.org/10.1038/nature02887
Miao Y, Stewart BA, Zhang F (2010) Long-term experiments for sustainable nutrient management in China. A review Agron Sustain Dev 31(2):397–414. https://doi.org/10.1051/agro/2010034
Morita A, Ohta M, Yoneyama T (1998) Uptake, transport, and assimilation of 15N-nitrate and 15N-ammonium in tea (Camellia sinensis L.) plants. Soil Sci Plant Nutr 44(4):647–654. https://doi.org/10.1080/00380768.1998.10414488
Mudau FN, Soundy P, Du Toit ES, Olivier J (2006) Variation in polyphenolic content of Athrixia phylicoides (L.) (bush tea) leaves with season and nitrogen application. S Afr J Bot 72(3):398–402. https://doi.org/10.1016/j.sajb.2005.10.007
Nadelhoffer KJ, Emmett BA, Gundersen P, Kjønaas OJ, Koopmans CJ, Schleppi P, Tietema A, Wright RF (1999) Nitrogen deposition makes a minor contribution to carbon sequestration in temperate forests. Nature 398(6723):145–148. https://doi.org/10.1038/18205
Okano K, Chutani K, Matsuo K (1997) Suitable level of nitrogen fertilizer for tea (Camellia sinensis L.) plants in relation to growth, photosynthesis, nitrogen uptake and accumulation of free amino acids. Japanese J Crop Sci. https://doi.org/10.1626/jcs.66.279
Oh K, Kato T, Xu HL (2008) Transport of nitrogen assimilation in xylem vessels of green tea plants fed with NH4-N and NO3-N. Pedosphere 18(2):222–226. https://doi.org/10.1016/S1002-0160(08)60010-7
Owuor PO, Cheruiyot DK (1989) Effects of nitrogen fertilizers on the aluminium contents of mature tea leaf and extractable aluminium in the soil. Plant Soil 119(2):342–345. https://doi.org/10.1007/BF02370429
Pregitzer KS, Burton AJ, Zak DR, Talhelm AF (2008) Simulated chronic nitrogen deposition increases carbon storage in northern temperate forests. Glob Chang Biol 14(0):142–153. https://doi.org/10.1111/j.1365-2486.2007.01465.x
Qiao C, Liu L, Hu S, Compton JE, Greaver TL, Li Q (2015) How inhibiting nitrification affects nitrogen cycle and reduces environmental impacts of anthropogenic nitrogen input. Glob Chang Biol 21(3):1249–1257. https://doi.org/10.1111/gcb.12802
Ranganatha V, Natesan S (1985) Potassium nutrition of tea. Madison, USA, pp 984–985
Ruan J (2005) Quality-related constituents in tea (Camellia sinensis (L.) O. Kuntze) as affected by the form and concentration of nitrogen and the supply of chloride. Christian-Albrechts Universität Kiel
Ruan J, Gerendás J, Härdter R, Sattelmacher B (2007) Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Annu Bot 99(2):301–310. https://doi.org/10.1093/aob/mcl258
Ruan J, Haerdter R, Gerendás J (2010) Impact of nitrogen supply on carbon/nitrogen allocation: a case study on amino acids and catechins in green tea [Camellia sinensis (L.) O. Kuntze] plants. Plant Biol 12(5):724–734. https://doi.org/10.1111/j.1438-8677.2009.00288.x
Ruan J, Ma L, Shi Y (2013) Potassium management in tea plantations: its uptake by field plants, status in soils, and efficacy on yields and quality of teas in China. J Plant Nutr Soil Sci 176(3):450–459. https://doi.org/10.1002/jpln.201200175
Ruan J, Wu X, Härdter R (1999) Effects of potassium and magnesium nutrition on the quality components of different types of tea. J Sci Food Agr 79(1):47–52. https://doi.org/10.1002/(SICI)1097-0010(199901)79:1<47::AID-JSFA172>3.0.CO;2-A
Sedaghathoor S, Torkashv AM, Hashemabadi D, Kaviani B (2009) Yield and quality response of tea plant to fertilizers. Afr J Agr Res 4:568–570
Tachibana N, Yoshikawa S, Ikeda K (1995) Influences of heavy application of nitrogen on soil acidification and root growth in tea [Camellia sinensis] fields. Japanese J Crop Sci
Tian D, Niu S (2015) A global analysis of soil acidification caused by nitrogen addition. Environ Res Lett 10(2):024019. https://doi.org/10.1088/1748-9326/10/2/024019
Tokuda S, Hayatsu M (2001) Nitrous oxide emission potential of 21 acidic tea field soils in Japan. Soil Sci Plant Nutr 47(3):637–642. https://doi.org/10.1080/00380768.2004.10408490
Tokuda S, Hayatsu M (2004) Nitrous oxide flux from a tea field amended with a large amount of nitrogen fertilizer and soil environmental factors controlling the flux. Soil Sci Plant Nutr 50(3):365–374. https://doi.org/10.1080/00380768.2004.10408490
USEPA (2008) Registering pesticides. Washington, D.C., pp 11–26
Venkatesan S, Ganapathy M (2004) Impact of nitrogen and potassium fertiliser application on quality of CTC teas. Food Chem 84(3):325–328. https://doi.org/10.1016/S0308-8146(03)00215-2
Venkatesan S, Murugesan S, Ganapathy MNK, Verma DP (2004) Long-term impact of nitrogen and potassium fertilizers on yield, soil nutrients and biochemical parameters of tea. J Sci Food Agr 84(14):1939–1944. https://doi.org/10.1002/jsfa.1897
Wang S, Zhu Y, Jiang H, Cao W (2006) Positional differences in nitrogen and sugar concentrations of upper leaves relate to plant N status in rice under different N rates. Field Crop Res 96(2):224–234. https://doi.org/10.1016/j.fcr.2005.07.008
Wang K, Liu F, Liu Z, Huang J, Xu Z, Li Y, Chen J, Gong Y, Yang X (2010) Analysis of chemical components in oolong tea in relation to perceived quality. Int J Food Sci Tech 45(5):913–920. https://doi.org/10.1111/j.1365-2621.2010.02224.x
Weerawatanakorn M, Hung WL, Pan MH, Li S, Li D, Wan X, Ho CT (2015) Chemistry and health beneficial effects of oolong tea and theasinensins. Food Sci Human Wellness 4(4):133–146. https://doi.org/10.1016/j.fshw.2015.10.002
Yu CP, Pan ZQ, Chen J, Fan DM, Wang XC (2012) Effects of Al3+ on growth and physiological characteristics of tea plant (Camellia sinensis). Plant Nutr Ferti Sci 1:182–187 (in Chinese)
Zak DR, Freedman ZB, Upchurch RA, Steffens M, Kögel-Knabner I (2017) Anthropogenic N deposition increases soil organic matter accumulation without altering its biochemical composition. Glob Change Biol 23(2):933–944. https://doi.org/10.1111/gcb.13480
Zhang M, Chen Y, ad F, Zhu Q, Pan Z, Fan K, Wang X (2017) Temporal evolution of carbon storage in Chinese tea plantations from 1950 to 2010. Pedosphere 27(1):121–128. https://doi.org/10.1016/S1002-0160(15)60098-4
Zhang X, Davidson EA, Mauzerall DL, Searchinger TD, Patrice D, Shen Y (2015) Managing nitrogen for sustainable development. Nature 528(7580):51. https://doi.org/10.1038/nature15743
Zheng SJ (2010) Crop production on acidic soils: overcoming aluminium toxicity and phosphorus deficiency. Ann bota-London 106(1):183–184. https://doi.org/10.1093/aob/mcq134
Zheng X, Xu Y, Li S, Hui R, Wu Y, Huang X (2013) Effects of green tea catechins with or without caffeine on glycemic control in adults: a meta-analysis of randomized controlled trials Am J Clin Nutr 97: 750-762. Doi: ajcn.111.032573v1
Zhu T, Zhang J, Meng T, Zhang Y, Yang J, Müller C, Cai Z (2014) Tea plantation destroys soil retention of NO3 − and increases N2O emissions in subtropical China. Soil Biol Biochem 73:106–114. https://doi.org/10.1016/j.soilbio.2014.02.016
Acknowledgments
The authors would like to gratefully acknowledge the anonymous reviewers for their comments on this manuscript.
Funding
This study was financially supported by the China Postdoctoral Science Foundation (2017M612390), the Science and Technology Project of Henan Province (172102110254), and the Youth Backbone Teachers Plan (2017GGJS-12) and Nanhu Scholars Program of XYNU.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
This article is part of the Topical Collection on Fertilisation
About this article

Cite this article
Qiao, C., Xu, B., Han, Y. et al. Synthetic nitrogen fertilizers alter the soil chemistry, production and quality of tea. A meta-analysis. Agron. Sustain. Dev. 38, 10 (2018). https://doi.org/10.1007/s13593-017-0485-z
Accepted:
Published:
DOI: https://doi.org/10.1007/s13593-017-0485-z