
Abstract
New material of known roveacrinids from two Cretaceous sites in Texas is described. Upper Albian strata at Saginaw Quarry furnished rich material of Poecilocrinus latealatus and Roveacrinus pyramidalis; for these two forms cups can be combined with primibrachials and secundibrachials. Some cups of Orthogonocrinus apertus are also present. Juvenile cups of these three species demonstrate ontogenetic changes, which are most prominent in P. latealatus. At the adult stage this species has cups with dish-like lateral wings; the first primibrachials have an aboral bowl; axillary second primibrachials and proximal secundibrachials have a wide aboral bowl; and distal brachials carry spines. The arms lack pinnules. A pelagic lifestyle with a mouth-up position is assumed. In contrast, R. pyramidalis has rod-shaped or stick-like brachials without wings, and food grooves are lacking. The less rich lower Cenomanian strata at the Dottie Lynn site provided a number of cups and brachials of Roveacrinus spinosus. In addition, cups of R. peracutus and R. alatus are present. Styracocrinus Peck is considered synonymous with Roveacrinus Douglas. The double cavity in the cups of the various species is discussed; its proposed function as a hydrostatic organ seems conjectural. Two genera of Saccocominae of the Late Jurassic radiation are thought to be ancestors to similar genera of Roveacrinidae first appearing during the Albian radiation. Derivation of Poecilocrinus may be from Saccocoma, Discocrinus and Plotocrinus possibly from Crassicoma. The ancestry of species of Roveacrinus and Orthogonocrinus with conical cups and two distinct body chambers is unclear. The closely similar juvenile cups of Roveacrinus pyramidalis and Poecilocrinus latealatus suggest a common origin despite the widely diverging arm structure. A comparison of Roveacrinus pyramidalis with the Triassic somphocrinid Osteocrinus reveals similar rod-shaped, smooth and tall brachials lacking a distinct food groove. Based on their mass occurrence and wide distribution, species of Osteocrinus species are thought to have been pelagic. This is substantiated by the presence of cups and rod-shaped brachials in Ladinian black shales of southern China. While food of species of Poecilocrinus presumably consisted of coccolithophores and planktonic foraminifera, collection and transport of food particles by Roveacrinus pyramidalis is enigmatic, as is that of Osteocrinus.
Similar content being viewed by others

Introduction
Remains of small stalkless crinoids of the family Roveacrinidae occur in great profusion in Cretaceous sediments of Texas. They were first described and well illustrated by Peck (1943) who established five new genera and 22 species, all based on disarticulated material. Subsequently, Rasmussen (1961) revised Peck’s taxa and proposed additional genera (Rasmussen 1971). Scott et al. (1977) described intact material of the distinctive Poecilocrinus dispandus Peck and made a reconstruction of the animal. For Peck (1943, p. 453), “the Roveacrinidae were planktonic, floating passively, mouth down and arms pendent, driven along with the current with little if any power to determine their direction of movement”.
For descriptions of roveacrinids from sites outside the United States the reader is referred to Rasmussen’s monograph (1961). Later descriptions are listed in the following. Destombes (1985) established a new genus (Hyalocrinus) from the Albian of France, and Destombes and Breton (2001) another new genus (Platelicrinus) from the Campanian of the same country. Meanwhile, Jagt (1999) had established Veugelersia and Birgenelocrinus from upper Campanian-upper Maastrichtian strata in the Netherlands, the latter representing the youngest named roveacrinid found so far (Jagt 2005). Salamon et al. (2010) described indeterminate roveacrinid remains from the lower Palaeocene of Poland, indicating that the order survived the Cretaceous–Paleogene extinction event, if only for a short time. Subsequently, Subsequently, Gorzelak et al. (2011) described some roveacrinid remains from the middle Miocene of Poland, thus characterizing the order as a “Lazarus taxon”. Nekvasilová and Prokop (1963) described remains of Orthogonocrinus and Roveacrinus from the Cretaceous of Bohemia; and Pisera (1983) recorded cups and brachials of Styracocrinus peracutus from Poland. Descriptions based on thin sections include species of Roveacrinus from the Albian of the Congo Basin (Ferré and Granier 2001) and a new species of Roveacrinus from the Cenomanian of Turkey (Farinacci and Manni 2003). Data on Cretaceous roveacrinids are summarised in the Revised Treatise (Hess in Hess and Messing 2011).
Morphological variability of cups, in part including also growth changes, was discussed by Sieverts (1932), Peck (1943, 1955), Scott et al. (1977), Jagt (1999) and Schneider (1987, 1988, 1989, 1995). However, very young, postlarval stages are largely unknown. Isolated brachials from the Cretaceous of Texas were figured by Peck (1943) and Rasmussen (1961) but could not be assigned to any given species or genus in most cases (Peck 1943, p. 452). Only Sieverts’ 1932 paper contained detailed information on brachials of a roveacrinid (Drepanocrinus sessilis Jaekel = Roveacrinus communis Douglas), including their position along the arms. Peck (1943, p. 456) noted that, “The Duck Creek Formation and the Grayson Marl yielded more crinoids and a greater variety of crinoids than the other Lower Cretaceous formations. The Duck Creek assemblage is easily recognised and differentiated from other assemblages, but many individual forms are difficult to identify specifically. The genus Poecilocrinus is represented by three species that are easily identified when mature, well-developed, and well-preserved, but they intergrade with each other to considerable extent”. In contrast Peck (1943, figs. 1–22) did see developmental trends in his material. The number of genera and species, all of considerable variability, but also limited data on ontogenetic changes, make the Cretaceous roveacrinids a taxonomically difficult group; and this includes not only assignment of species to genera but also definition of genera. In the Revised Treatise (Hess and Messing 2011) Rasmussen’s (1978) diagnoses of the Cretaceous genera described previously by Peck were largely adopted. The problems of assigning brachials to cups in most cases and the lack of reconstructions have so far prevented a deepened discussion on the mode of life of the different forms.
For the present paper new material from the upper Albian Duck Creek Formation and the lower Cenomanian Del Rio Clay was available. It was collected and provided by Andrew S. Gale. The Duck Creek sample is much richer in roveacrinid remains than that from Del Rio and, thus, forms the bulk of the paper. Its main goal is to illustrate ontogenic changes and variability of cups as well as of brachials assignable to cups. These additional data contribute to a reappraisal of the systematics of some species and their synonymy. The samples contain very distinctive species; and the availability of brachials enables a discussion of functional morphology and mode of life of the Cretaceous Roveacrinidae in comparison with the Jurassic Saccocomidae (Hess and Etter 2011) and the Triassic Somphocrinidae (Wang et al. 2007, Wang et al. 2008; Hess in Hess and Messing 2011) (Fig. 1).

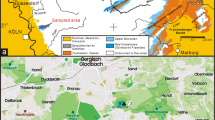
Geological setting and material (Figs. 2, 3)
The two sites are in the Fort Worth area, and about 50 kg were sampled at both places. The first sample was taken 0.5 m beneath the top of the upper Duck Creek Formation, Mortoniceras rostratum Zone [=Heterohelix reussi Zone (planktonic foraminifera)] at the large abandoned Saginaw Quarry (Fig. 2). Here the entire Duck Creek Formation and the base of the overlying Fort Worth Formation (both of late Albian age) are exposed. Important markers beneath the sampled mudstone include a triplet of thin limestones and a limestone containing abundant oysters, both of which are in the upper Duck Creek Formation and are correlatable regionally. The second sample was collected at the Dottie Lynn Locality (Fig. 3). The section exposes about 8 m of dark grey clays and micritic limestones, with thin silty lenses and carbonate concretions, and it belongs to the lower Cenomanian Del Rio Clay Formation. The fauna is dominated by large calcitic bivalves (oysters, Neithea), and rarer ammonites are present (Stoliczkaia, Plesioturrilites). The Del Rio Clay Formation here is overlain disconformably by the middle Cenomanian Woodbine Formation, formerly visible at the top of the section (F. Holterhoff, pers. comm., 2012). The sample was taken 1.5 m beneath the top of the Del Rio Clay Formation.

Profile of the Saginaw locality, raised by A. S. Gale, May 1993

Profile of the Dottie Lynn locality, raised by A.S. Gale, October 2011
The isolated and washed crinoid ossicles were cleaned in an ultrasonic bath. Photographs were taken at the Zentrum für Mikroskopie of the University of Basel by E. Bieler, using a Philips XL30 ESEM and gold-coated samples.
Repository. Naturhistorisches Museum Basel.
The Duck Creek roveacrinids
Specimens belong to four distinctive roveacrinid types. By far the most common and largest form is represented by cups and proximal brachials with spectacular horizontal dish-like wings; on distal brachials the wings are replaced by thorns or spines. The many thousands of remains are assigned to Poecilocrinus latealatus (Figs. 4, 5, 6, 7, 8). Less common is a small species with radial extensions on the cup but lacking horizontal wings; it is assigned to Roveacrinus pyramidalis (Fig. 9). A series of most peculiar rod-shaped or stick-like brachials are assigned to this species (Figs. 10, 11). The third form is represented by large, angular cups with radial flanges, but devoid of lateral wings or processes; it is referred to Orthogonocrinus apertus (Fig. 12). A few primibrachials may belong to this species (Fig. 13). Finally, a single, small and broken cup is assigned to Roveacrinus peracutus (Fig. 12h).

Poecilocrinus latealatus (Peck), upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. Ontogenetic series of cups in aboral view. a M11564, b M11561, c M11548, d M11556, e M11560, f M11544, g M11554, h M11539, i M11555, j M11533, k M11537, l M11570, m 11536, n M11538, o M11545, p M11552, q M11558, r M11543, s M11541, t M11577

Poecilocrinus latealatus (Peck), upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. Ontogenetic series of cups in lateral (a, b, i–l, q–r) and oblique-adoral view (c–h, m–p). a M11567, b M11583, c M11562, d M11578, e M11547, f M11559, g M11584, h M11574, i M11565, j M11540, k M11557, l M11581, m M11576, n M11527, o M11573, p M11550, q M11551, r M11553

Poecilocrinus latealatus (Peck), upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. a Cup with base of crown in lateral view, arrows indicate bowls on aboral side of first primibrachials; M11522. b Lateral view of broken cup, partition between upper and lower chamber weakly developed; M11523. c Broken cup with well-developed partition between the two chambers, M11525. d Oblique-adoral view of cup showing articular facets and moderate interfacet processes, flanges to partly broken dish-like wings well developed; M11569. e Adoral view of partly broken cup, lower chamber is visible on top, flanges connecting facets with dish weakly developed; M11528. f Oblique-adoral view of cup with well-developed facets, wings broken away; M11571. g Adoral view of small cup, M11530. h Oblique lateral-adoral view of cup, interfacet processes and flanges to broken dish-like wings well-developed; M11572. i Lateral view of cup with high interfacet processes, M 11535. j Lateral view of cup with comparatively small aboral part, lateral wings broken away, M11534. k Adoral view of cup with unequal lateral wings, M11529. l Adoral view of cup with weak interfacet processes but distinct flanges connecting facets to dish, M11532

Poecilocrinus latealatus (Peck), upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. Axillary second primibrachials (a–f) illustrating range of shape. a M11608; a1 aboral, a2 adoral. b Distal view with the two articular facets to first secundibrachials, M11654. c M11666. d M11650. e M11667. f M11651. First primibrachials illustrating size and shape of bowl on aboral side proximal to cryptosynarthrial facet (g–s). g Aboral-distal view, M11612. h Aboral-distal view, M11615. i Aboral-distal view with particularly wide bowl, M11613. j Aboral-distal view, M11611. k Aboral-distal view, M11614. l Aboral-distal view, M11645 (this ossicle is also figured in Fig. 13a under Roveacrinus peracutus to which it may alternatively belong). m Aboral-distal view with wide bowl, M11644. n Proximal-adoral view, M11610. o Proximal-adoral view with protruding bowl visible adorally, M11643. p Lateral view showing bowl and muscle fossa along edge of cryptosynarthrial facet at right, M11609. q Proximal facet showing wide axial canal and aboral ligament pit below, M11531. r Aboral-distal view of specimen with protruding elliptical bowl, M11647. s Aboral-distal view of specimen with wide protruding bowl, M11646

Poecilocrinus latealatus (Peck), upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. Secundibrachials of various morphologies, concavity of wings is directed aborally. a Brachial with shallow-concave aboral wing and two muscular facets, muscle fossae corrugated; M11661. b Brachial with deep-concave aboral wing, muscle fossae corrugated; M11655. c Brachial with deep, elliptical wing, M11665. d Proximal or distal-adoral view of brachial without wing, muscle fossae corrugated, other side synostosial; M11659. e Adoral view of wingless brachial, both facets muscular, muscle fossae corrugated; M11670. f Adoral view of winged brachial, M11663. g Adoral view of winged brachial, facet directed downwards cryptosynarthrial; M11662. h Adoral-proximal view of winged brachial with cryptosynarthrial facet, M11652. i Synostosial facet of wingless brachial, other facet muscular with corrugated fossae; M11660. j Synostosial facet of winged brachial, M11648. k Winged brachial with synostosial facet matching brachial, M11649. l Adoral-proximal view of winged brachial with cryptosynarthrial facet, M11653. m Lateral view of winged brachial with muscular (right) and cryptosynarthrial facet (top), wing at left; M11657. n Synostosial facet of wingless brachial, other side muscular with corrugated muscle fossae; M11658. o Muscular facet of brachial with corrugated fossae, other facet synostosial; M11656. p Brachial with spoon-like wing and corrugated muscle fossae; M11628. q Brachial with leaf-like wing, M11634. r High brachial with deep food groove and corrugated muscle fossae, both facets muscular, aboral wing with ridge; M11664. s Smaller brachial similar to r, M11672. t Brachial with irregular, nearly spine-like wing, both facets muscular; M11677. u Brachial with aboral spine and corrugated muscle fossae, both facets muscular; M11629. v Brachial with dagger-like spine, both facets muscular, muscle facets corrugated; M11671. w Brachial with spine, both facets muscular, muscle facets weakly corrugated; M11669

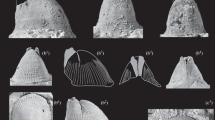
Roveacrinus pyramidalis Peck, upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. Ontogenetic series of cups. a Aboral view, apex partly damaged, M11596. b Aboral view with two weak radial spines, apex not preserved, M11594. c Aboral view of cup with one radial spine, apex not preserved, M11585. d Aboral view of cup with long radial spine, apex not preserved, M11595. e Aboral view of cup with intact apex and five unequal radial spines, M11586. f Lateral view, radial processes moderately produced, M11590. g Lateral view of conical cup with one prominent radial process, interfacet processes unequal; 11591. h Lateral view of cup with high interfacet processes, radial processes moderately produced; M11592. i. Lateral view of cup with low interfacet processes, radial processes well-produced; M11589. j Lateral view of cup with high interfacet processes, radial processes unequal; M11587. k Lateral view of cup with single high interfacet process, radial processes projecting wide cup; M11593

Primibrachials of Roveacrinus pyramidalis Peck, upper Duck Creek Formation, M. rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. a Distal facet of non-axillary second primibrachial, M11696. b Aboral view with distal facets of axillary second primibrachial, shaft broken; M11620. c Distal view of axillary second primibrachial, M11617. d Lateral view of high second primibrachial, note small cryptosynarthrial facet to first primibrachial (q); M11673. e Oblique-adoral view of slender second primibrachial with small facets to first secundibrachials, note row of pits on shaft; M11632. f Aboral view of keeled second primibrachial with facets to first secundibrachials, shaft broken; M11686. g Adoral view of broken second primibrachial with high cryptosynarthrial facet to first primibrachial, M11625. h Adoral view of intact second primibrachial with regular row of weak pits, M11684. i Lateral view of second primibrachial with large facet to first primibrachial, M11683. j Aboral view of first primibrachial with groove and cryptosynarthrial facet, M11675. k Aboral view of first primibrachial with cryptosynarthrial facet, note reticulate structure in proximal part; M11641. l Aboral view of slender first primibrachial, M11676. m Lateral view of compressed first primibrachial, M11690. n Adoral view of first primibrachial, M11616. o Proximal-adoral view of first primibrachial, arrow on enlarged ossicle indicates muscle fossae; M11679. p Oblique proximal-lateral view of laterally compressed first primibrachial, M11681. q Lateral view of first primibrachial, distal facet matches proximal facet of large second primibrachial (d); M11674. r Lateral view of cup (Fig. 9i), M11589

Secundibrachials, upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. Most of the brachials are assignable to Roveacrinus pyramidalis Peck. a Muscular facet of distal brachial with adoral processes, M11626. b Lateral view of distal brachial with adoral processes on both muscular facets, M11622. c Proximal-adoral view of distal brachial with row of pits on shaft, M11623. d Proximal-adoral view of medial brachial with muscular facets, note food groove dissolved into pits along shaft, M11619. e Aboral-distal view of medial brachial with keeled shaft, M11621. f Lateral view of medial brachial with pinnule socket, M11635. g Lateral view of medial brachial with pathological (?) process, M11695. h Lateral view of compressed brachial, brachial has sharp keel at left (aborally) and furrow at lower right (adorally), proximal muscular facet bent, distal muscular facet with processes for muscle fossae; M11688. i Brachial similar to h but with pinnule socket, M11692. J–l Aboral view of keeled elongate brachials, muscular facets at both ends; M11689 (j), M11688 (k), 11668 (l). m–p Presumed first secundibrachials, proximal facets muscular, distal facets cryptosynarthrial to synostosial; adoral view (m), aboral view (n–o), lateral view (p); M11633 (m), M11693 (n), M11627 (o), M11624 (p)

Cups of Orthogonocrinus apertus Peck (a–g, i–l) and Roveacrinus peracutus Peck (h), upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. a Lateral view of cup with unequal interfacet processes and partly broken aboral part, M11606. b Adoral view, M11599. c Oblique-adoral view of cup broken below facets, M11600. d Cup broken vertically and aborally, with partition separating small upper and large lower body chamber, M11597. e Aboral view of broken cup, partition with five peripheral radial openings and a central opening connecting the two chambers; M11607. f Small, nearly intact cup, interfacet processes low; M11639. g Smallest cup composed of bulging radials with intact base, suggesting presence of basals; M11691. h Broken cup of Roveacrinus peracutus, lateral view; M11605. i Lateral view of cup with equal interradial processes, M11601. j Lateral view of broken cup with unequal interfacet processes, M11603. k Lateral view of broken cup with additional vertical ridges and unequal interfacet processes, M11598. l Lateral view of large, broken cup with unequal processes, note right angle between aboral ligament pit and adoral part of articular facet; M11602

Primibrachials tentatively assigned to Orthogonocrinus apertus Peck (b–c, e–h) and Roveacrinus peracutus (Peck) (a, d), upper Duck Creek Formation, Mortoniceras rostratum Zone (upper Albian); Saginaw Quarry, Fort Worth, Texas. a Aboral-distal view of first primibrachial lacking distinct bowl, M11645 (this specimen is also figured in Fig. 7l under Poecilocrinus latealatus). b Adoral-proximal view of first primibrachial with facet to radial, M11638. c Adoral-proximal view of first primibrachial with facet to radial, M11640. d Adoral-proximal view of first primibrachial with rows of pits, M11642. e Aboral-distal view of first primibrachial with corrugated edges, M11637. f Adoral view of second primibrachial, M11630. g Aboral view of second primibrachial lacking bowl, M11680. g Aboral view of second primibrachial lacking bowl, M11631

Cups of Roveacrinus alatus Douglas (d, p, t) and Roveacrinus spinosus Peck (a–c, e–o, q–s). Del Rio Formation (lower Cenomanian), Dottie Lynn Locality, Fort Worth, Texas. a Lateral view of smallest cup, M11706. b Lateral view of cup with prominent radial flanges, M11700. c Aboral view of broken cup showing partition with central opening and smaller radial openings at periphery, M11702. d Aboral view of cup of R. alatus, M11708. e Lateral view of cup with prominent radial flanges and bulbous aboral part, M11723. f Adoral-lateral view of cup with equal radial processes, M11711. g Adoral-lateral view of cup with unequal, broad radial processes; M11712. h Adoral view of cup with only two narrow radial processes, M11714. i Adoral view of cup with equal, broad radial processes; M11710. j. Lateral view of cup with short processes and irregular radial flanges, M11701. k Lateral view of cup well-developed radial processes and flanges reaching beyond bulbous aboral apex, M11704. l Lateral view of cup with irregular, short processes and moderately produced radial flanges, M11727. m Lateral view of cup with duckbill-shaped radial processes, M11713. n Part of cup with strongly bent concave process and wide articular facet, M11709. o Adoral-lateral view of cup with single radial process, M11703. p Adoral view of cup of R. alatus with unequal radial extensions, articular facets badly preserved; M11705. q Lateral view of cup with prominent radial extensions and flanges, one high interradial process; M11715. r Lateral view of cup with widely extended processes and flanges, M11716. s Cup with one wide and strong radial process, M11699. t Lateral view of cup of R. alatus with thin radial flanges supported by lateral wing; M11707

Brachials. Del Rio Formation (lower Cenomanian), Dottie Lynn Locality, Fort Worth, Texas. a, b First primibrachials, possibly belonging to Roveacrinus alatus Douglas; a aboral-distal view, M11734; b adoral-proximal view, M11739. c–w Brachials assigned to Roveacrinus spinosus Peck. c Aboral-distal view of first primibrachial, M11738. d Adoral-proximal view of first primibrachial, M11737. e Proximal-aboral view of first primibrachial, M11742. f Aboral view of strongly sculptured second primibrachial, M11732. g Adoral-distal view of second primibrachial, M11740. h Lateral view of second primibrachial; proximal, cryptosynarthrial facet at left; M11741. i Aboral-distal view of second primibrachial with two spines, M11746. j Aboral view of second primibrachial with one lateral spine, M11747. k Aboral view of second primibrachial without spine, M11743. l Aboral-distal view of sculptured second primibrachial, M11735. m Adoral view of secundibrachial with pinnule socket (arrow), M11745. n Distal view of secundibrachial with aboral spine and corrugated muscle fossa on higher side, M11733. o Secundibrachial with corrugated muscle fossae, M11724. p Asymmetric secundibrachial with aboral spine, high side with corrugated muscle fossae; M11748. q Synostosial facet of secundibrachial with strong spine broken at end, M11730. r Synostosial facet of secundibrachial with sculptured aboral surface, other side muscular with corrugated muscle fossae; M11729. s Secundibrachial with deep food groove and long spine, muscle fossae corrugated; M11731. t Aboral-proximal view of secundibrachial with muscular facet and moderate corrugated muscle fossae, M11744. u Lateral view of secundibrachial with pinnule socket (arrow), M11728. v Oblique lateral view of secundibrachial with pinnule socket (arrow), M11726. w Aboral view of secundibrachial with pinnule socket at left (arrow) and spine at right, M11725
Peck (1943, fig. 23) noted the following genera and species from the Duck Creek Formation: Drepanocrinus peracutus Peck, Orthogonocrinus apertus Peck, Roveacrinus pyramidalis Peck, Plotocrinus hemisphericus Peck, Pl. modulatus Peck, Pl. inornatus Peck, Poecilocrinus porcatus Peck, Po. pendulus Peck, Po. spiculatus Peck, and Discocrinus catastomus Peck. Thus, from this list, only Orthogonocrinus apertus, Roveacrinus peracutus and R. pyramidalis are represented in our material.
Poecilocrinus latealatus (Peck, 1943), Figs. 4–8.
1943 Roveacrinus latealatus Peck, Peck, p. 468; pl. 73, figs. 9–12, 14; pl. 76, fig. 1.
1943 Plotocrinus hemisphericus Peck, Peck, pl. 71, fig. 4 (first primibrachial).
1961 Poecilocrinus latealatus (Peck), Rasmussen, p. 380; pl. 55, figs. 4–5.
Cups Available cups have a wide size range, from a diameter of about half a millimetre to 3 mm. Approximately 6000 cups were examined, roughly half of them juvenile or subadult with lateral extensions or wings not yet fused into dishes. The smallest cups are low, open bowls; the aboral apex, which may be flattened, carries interradially five knobs, presumably representing vestigial basals (Fig. 4a, b). The radial articular facets slightly protrude from the upper rim so that the cups achieve a pentagonal outline at this stage (Fig. 4a, b). During growth the area below the facets is prolonged downwards and outwards developing into spoon-shaped extensions that are adorally concave (Fig. 5d–j). The outline of the extensions may vary from rounded (Figs. 4c, d; 5k, l) to spearhead shaped (Fig. 4f, h, j). During further growth the extensions increase in size until they finally merge into a wide, thin wing that is flat aborally (Fig. 4q–t) and slightly concave adorally (Fig. 6d–e). The outline of the lateral wing is mostly circular in aboral view (Fig. 4q), but may also be pentagonal (Fig. 4t). In rare cases the dish comprised more than five pieces (Fig. 4m); cups in which individual parts are lobed are somewhat more common (Fig. 4k, o). Aborally, the wings carry a medial rib or flange that extends to the apex in most cases. However, size and shape of the ribs are quite variable, especially in adult specimens. They may be curved (Fig. 4n) or branched (Fig. 4k, l). Alternatively, they may be strongly developed and thick on the aboral, globular part of the cup (Fig. 4p, q, t), they may be absent in part (Fig. 4m) of the aboral apex or altogether (Fig. 4j, l, s). In a number of specimens the aboral apex is fused with the lateral dish (Fig. 4s). In larger (adult) specimens the ribs commonly are quite thick (Fig. 4p, q, t).
Morphological changes during ontogeny are also demonstrated in lateral view (Fig. 5). The smallest cups of about half a millimetre in diameter are bowl shaped, with a flat base and radial articular facets on the upper rim (Fig. 5a, b). Growth of the processes beneath the facets starts at cup diameters approaching 1 mm (Fig. 5c, d). The downward-directed processes are first pointed (Fig. 3e–g), but then broaden and become spoon-shaped (Fig. 5h–l). However, pointed, lanceolate extensions may persist to a later stage (Fig. 5m). The spoons are supported by a rib or flange that may be quite prominent, extending beyond the aboral apex (Fig. 5p, q). At this stage the spoons are in lateral contact and eventually the lateral wings merge into a bowl that is slightly concave adorally. Cups of similar size may have widely separated (Fig. 5p) or nearly merged spoons (Fig. 5q, r; see also Fig. 4d–i). On the adoral side of post-juvenile to adult cups the radial articular facets and their interfacet processes are arranged in a ring lying on the shallow bowl of the dish formed by the lateral wings. In juvenile specimens the interfacet processes, directed upwards, are poorly developed, although present in some specimens (Fig. 5d–f). On the surface of the dish are deep clefts below the interfacet processes, but the clefts do not extend into the cavity (Fig. 6). In some specimens the interfacet processes may be of similar height (Fig. 6j), but they may also be quite variable (Fig. 6b, i).The sutures of the radials have a double ridge adorally (Figs. 5e, f, 6c, d, f, h). Broken cups commonly display a double body cavity (Fig. 6b, c, e). The upper or radial cavity is shallow and moderately wide; the lower or aboral cavity forms the globular part of the cup. A thin, mostly broken, wall may separate the two cavities (Fig. 6e), but such a wall is not present in all specimens. The radial articular facets have a deep aboral ligament pit separated from the somewhat wider axial canal by a narrow transverse ridge; the interarticular ligament fossae are indistinct, and the muscle fossae extend onto the interfacet processes (Fig. 6h–j). The variable size of the processes begs the question on muscle activity between the cup and the first primibrachials whose muscle fossae are tall and uniform (Fig. 7n–p).
Primibrachials Primibrachials number in the thousands. One specimen is preserved with cup and proximal arms still attached (Fig. 6a). The first primibrachials have a circular bowl on the aboral surface (arrows; the second primibrachial follows just above the bowl). Articulation with the second primibrachials and the following secundibrachials is not exposed because the ossicles are tightly connected, but all have an aborally concave wing. The first primibrachials have an aboral surface with a bowl proximally and an elliptical cryptosynarthry distally (Fig. 7g–s). The bowl varies from barely apparent (Fig. 7l; alternatively, this ossicle may belong to Roveacrinus peracutus, see Fig. 13a) to large circular (Fig. 7i, m) and wide elliptical (Fig. 7o, r, s). The rim may be thickened in small bowls (Fig. 7g, h, j, k). In side view the first primibrachials are wedge shaped, with the aboral bowl conspicuously standing out (Fig. 7p). The proximal, muscular facet (Fig. 7q) matches the radial articular facet of the cup; in proximal-adoral view the facet extends into two lateral muscle fossae characterised by dense stereom (Fig. 7n–q). The second primibrachials (Fig. 7a–f) are axillary. The proximal side has a cryptosynarthrial facet, matching the distal facet of the first primibrachial. The two distal facets are muscular and are also present adorally. The body of the ossicle with the proximal and the two distal facets is on the lower side of the wing that extends laterally and distally beyond the facets; and a knob may separate the distal facets (Fig. 7a2, b, d–f). The aboral side is concave, commonly with a central rib (Fig. 7a1). Ribs may also be developed on the adoral side (Fig. 7b, d).
Secundibrachials There are many thousands of these ossicles. The arms beyond the second primibrachials do not bifurcate again. There is no indication of the presence of pinnules because corresponding sockets are lacking. In the proximal part of the crown most brachials have a wing (Fig. 8a–c, f–h, g, j–l), similar to that of the second primibrachials. The muscle facets commonly are corrugated to various degrees (Fig. 8a–d), and this is also true of brachials without wings (Fig. 8n–p, r–w). A number of brachials are connected by synostoses (Fig. 8i–o), and these occur on brachials with or without (Fig. 8i, n, o) wings. In more distal brachials the wing becomes elongate and keeled, assuming various shapes (Fig. 8p–t). Most of the distal brachials carry a conspicuous spine (Fig. 8u–w) and have only muscular facets.
Remarks Scott et al. (1977) ascribed more or less intact specimens from the Weno Formation of Fort Worth to Poecilocrinus dispandus (their reconstruction is here reproduced in Fig. 16). They are similar to the present material in certain characters, such as proximal brachials with aborally concave wings, distal brachials with spines, and the presence of synostosial arm articulations. However, specimens of P. dispandus differ in the following features. (1) The aboral part of the cup has strong ribs connected ring-like around the periphery of the apex (see Peck 1943, pl. 75, fig. 6). (2) The first primibrachial lacks a bowl or dish on the aboral surface (see Scott et al. 1977, pl. 2, fig. 4); and (3) the terminal part of the arm is pinnulate (see Scott et al. 1977, pl. 1, fig. 5). These differences suggest that the two forms do not belong to the same species. Peck (1943) described a number of species with lateral wings on the cup that he assigned to the genera Roveacrinus and Poecilocrinus. They include Poecilocrinus dispandus (with the four subspecies dispandus, explicatus, elongatus, and molestus), Po. pendulus, Po. spiculatus, Roveacrinus latealatus and R. signatus. The two last-named species were transferred by Rasmussen (1961) to Poecilocrinus. However, only Po. pendulus and Po. spiculatus are mentioned by Peck (1943) from the Duck Creek Formation. Poecilocrinus dispandus, the type species of the genus, and its subspecies are from sites of the upper Albian Fort Worth to Main Street formations. Poecilocrinus pendulus differs from the present specimens by the downward-concave, circular wing on the radial, developed as a deep bowl (Peck 1943, pl. 75, figs. 5, 10, 13). This character is also present in Poecilocrinus spiculatus; see Peck (1943, pl. 73, fig. 6) and Rasmussen (1961, pl. 55, fig. 1a, b), who suggested that the specimens referred to this species are actually young individuals of P. pendulus. The present cups are comparable to cups of two forms described by Peck (1943). These are Roveacrinus latealatus from the Fort Worth Formation and Roveacrinus signatus which is confined to the Main Street and Grayson formations. According to Peck (1943) and Rasmussen (1961) the two species are distinguished by the development of the radial ridge or flange which leads from the aboral apex to near the edge of the wing. Rasmussen (1961, p. 380) diagnosed P. latealatus as follows: “A Poecilocrinus with a prominent, narrow, radial ridge from the dorsal point (=aboral apex) to the horizontal wing just below the articular face. The horizontal wing is very large, confluent in the interradius with the neighbouring wings around the theca (=cup), but with distinct sutures. The side-branches from the radial ridge to support the horizontal wing along the sutures are rudimentary”. Poecilocrinus signatus was diagnosed by Rasmussen (1961, p. 381) as follows: “A Poecilocrinus with a prominent radial flange from the dorsal point to the horizontal wing just below the articular face. Side-branches from each radial flange are angularly bent and are continued in the two halves of the horizontal wing, forming together a bowl around the top of the radial ridge. The radial bowls and horizontal wings are greatly enlarged during growth, and in large specimens they are more or less confluent in the interradius”. In his discussion of P. latealatus, Peck (1943, p. 468) mentioned that, “superficially, R. latealatus closely resembles P. dispandus…The two species cannot be differentiated from an oral view. From an aboral or side view it is evident that on R. latealatus the main radial flange continues all the way from the aboral pole to the proximal portion of the radial facet. On representatives of P. dispandus the flanges fork about mid-height from the calyx (=cup)”. Peck also mentioned that, “R. latealatus is probably the ancestor of R. signatus of the Main Street limestone and Grayson marl, although representatives of this line of development have not been found in the intervening formations”. Peck’s holotype of R. latealatus (his pl. 73, fig. 14), compares well with our specimen in Fig. 4q, and Peck’s lateral view of specimen (his pl. 73, fig. 9) is comparable to Fig. 6j in the present paper. Adoral views are also similar (Peck 1943, pl. 73, fig. 12 and Fig. 6d and l here). Peck’s figures of R. signatus show specimens with forked aboral flanges (Peck 1943, pl. 74, figs. 1, 3) that are quite similar to specimens in the present material (Fig. 4k, l). In addition, Peck’s pl. 74, fig. 11 and 13 are comparable to our Fig. 4d, f, although the bowl is downward-concave in Peck’s specimens. Thus, our Duck Creek specimens have features of two of Peck’s species that are stratigraphically younger. A first primibrachial figured by Peck (1943, pl. 71, fig. 4) has an aboral bowl beneath the cryptosynarthrial facet, a distinctive character of our Duck Creek specimens. This ossicle was assigned by Peck to Plotocrinus hemisphericus, “a characteristic Duck Creek species”. Peck also figured some brachials with serrated processes comparable to some of our specimens. However, no cup assignable to P. hemisphericus is in the present material. The morphological changes during development as well as the large variation in adult specimens prevent classification into more than one species. Because the majority of cups in our material are closer to the holotype of P. latealatus than to that of Poecilocrinus signatus I assign them to the former. The Duck Creek specimens with forked ribs (Fig. 4k, l, n) suggest a line of development leading to P. signatus. Such development was already suggested by Peck (1943, figs. 1–22).

Reconstruction of Poecilocrinus dispandus Peck from the Weno Formation (upper Albian). Approximately ×10 (from Scott et al. 1977)
The genus Poecilocrinus was established by Peck (1943, p. 471) with the following diagnosis: “Roveacrininae in which the radial ornamentation consists of a single flange that bifurcates at mid-height or below, each branch connecting with the outside proximal portion of the arm facet to form an enclosed, more or less circular depression under the arm facet, the depression on most specimens remaining hollow but being occasionally bisected by a vertical flange of secondary origin”. This diagnosis is difficult to follow and was modified by Rasmussen (1961, p. 378) to read, “Roveacrinidae in which the theca (=cup) is provided with a prominent, horizontal wing on each radial, sometimes fused around the theca, or each radial has a curved, bowl-shaped wing with the opening of the concavity outward or downward. There is a radial ridge from the wings to the dorsal point (=aboral apex). The articular facets are fairly large, outward-sloping, and connected with the wings by short lateral ridges. The interfacet processes are rather short and stout. The radial cavity is large.” This diagnosis, adopted in the Revised Treatise (Hess and Messing 2011), may be enlarged as follows: “Proximal brachials with aborally concave wing, distal secundibrachials with spine; corrugated adoral processes common, a number of secundibrachials articulated by flat synostosis.”
Roveacrinus pyramidalis Peck, 1943, Figs. 9–11.
1943 Roveacrinus pyramidalis Peck, Peck, p. 468; pl. 72, figs. 24–29.
1943 isolated brachials of unknown affinities, Peck, p. 473; pl. 76, figs. 23, 27, 42, 44.
?1943 Plotocrinus primitivus Peck, Peck, p. 470; pl. 71, figs. 1–3.
1961 Roveacrinus pyramidalis Peck, Rasmussen, p. 371; pl. 53, figs. 10–11.
1961 brachials of Roveacrinidae, Rasmussen, pl. 56, figs. 9–11.
Cups (Fig. 9) The material contains 375 cups. The diameter ranges from about 0.5 to 1 mm. The outline is roughly pentagonal, and the profile more or less pyramidal. Some small cups are broken above the aboral apex, exposing a lower chamber with spherical bodies, presumably basals (Fig. 9a–d). The surface is coarsely reticulate. The articular facets are roughly triangular and prolonged downwards into prominent processes of irregular shape, giving the cups a ragged appearance. The processes also occur in smaller individuals where they are produced into spines (Fig. 9b–e); these may not be developed on all sides. The articular facets have a deep aboral ligament pit separated from the small central canal by a narrow transverse ridge. The interarticular ligament fossae are indistinct, and the muscle fossae extend onto interfacet processes of variable height; some cups may have processes of different height (Fig. 9h, k).
Primibrachials (Fig. 10) The material includes a significant number (373, many of them broken) of tall second primibrachials that were attached by a sloping cryptosynarthrial facet to the smaller first primibrachials (Fig. 10d, q). There are only 17 first primibrachials that can be combined with the axillaries. They have a roughly triangular proximal muscular facet (Fig. 10n, o) matching the facet of the cups (Fig. 10r). The muscle fossae are prolonged adorally, matching the corresponding processes on the cup (Fig. 10o, arrow). Their proximal part is slightly thickened and has a reticulate structure comparable to that of the cup (Fig. 10k, l, q). The shaft is smooth and composed of dense stereom. A series of small pits may be developed along the adoral–aboral margin (Fig. 10o). The distal facet, seen aborally, is a strongly sloping cryptosynarthry of somewhat variable height. However, most of the available ossicles have a facet that is lower than the proximal facet of the following second primibrachial. These axillary ossicles are unusually tall. The proximal facet is cryptosynarthrial and strongly sloping. The facet may sit on a pedestal (Fig. 10d) but mostly reaches the lower (proximal) end (Fig. 10e, g–i). In large ossicles this part resembles the sole of a shoe in side view (Fig. 10i). In section, the second primibrachials are elliptical to laterally compressed; exceptionally, they are adorally keeled (Fig. 10f). The shaft commonly bears a row of shallow pits on the adoral side (Fig. 10e, g, h); occasionally there is a narrow, weak continuous furrow. At the distal end are two muscular facets for articulation with the first secundibrachials (Fig. 10b, c, f, h, i), they are directed outwards (aborally) at an angle of about 60°; exceptionally there is only one facet so that the arm did not branch in this case (Fig. 10a). The facets have a strong relief, with pronounced aboral ligament fossa and axial canal, flanked by similarly produced interarticular ligament fossae; the muscle fossae extend onto narrow processes. Such articulation suggests considerable power for arm movement at this place.
Secundibrachials (Fig. 11) The secundibrachials number in the thousands. They vary greatly in shape (Fig. 11), and many cannot be assigned to a definite position in the crown. Similar to the primibrachials, none has a distinct food groove. No further axillary brachials have been found so that the arms must have branched only at the second primibrachials. Brachials illustrated in Fig. 11m–p appear to be first secundibrachials. The proximal facet is muscular, the distal synostosial and outward-sloping. The aboral side of the shaft has a median ridge and lateral flanges. The ossicles vary in height. First secundibrachials of similar shape were figured by Rasmussen (1961; pl. 56, fig. 3) in a cup of R. peracutus, from the Cenomanian of Cambridge, with attached primibrachials. The second primibrachials (primaxils) of this specimen are also high and slender, but they differ from the present material by a narrow median ridge and thin lateral flanges. Some brachials (Fig. 11h–i) are laterally compressed, similar to Fig. 11p; yet distally they have muscular facets with long processes for muscle fossae. Other brachials (Fig. 11d–e) have muscular facets at both ends matching the distal facets of the second primibrachials (Fig. 10a–c). However, they cannot be first secundibrachials because the distal facet is also muscular, not synostosial. Second secundibrachials with proximal synostosial facet and distal muscular facet have not been found. A number of brachials appear to have a pinnule socket (Fig. 11f, i), although in other brachials of similar morphology such a structure is weak (Fig. 11h). A single brachial carries a worm-like process, perhaps an epizoan (Fig. 11g). The material includes quite a number of tall brachials with an aboral keel and muscular facets at both ends (Fig. 11j–l), they are presumably from median arm portions. Somewhat smaller brachials are strongly compressed laterally and, except for a keel, almost transparent (not figured). The smallest brachials from distal parts of the arms are common. They have muscular articulations at both ends and adoral processes for muscle attachment (Fig. 11a–c). As in the other secundibrachials distinct food grooves along the body are lacking, but small impressions similar to the axillary primibrachials (Fig. 10g–h) may be seen on the adoral side (Fig. 11c).
Remarks Peck (1943, pl. 76) figured a number of brachials from the Duck Creek Formation which he could not assign to any given species. In this he was followed by Rasmussen (1961; pl. 56, figs. 9–11) who did not discuss either the particularly tall, rod-shaped second primibrachials or their possible function. Similar brachials with a height of 1 mm or so, including axils, were figured by Kristan-Tollmann (1970, fig. 5) from the Cassian Formation (upper Ladinian/Carnian) of the Dolomites (see also Hess in Hess and Messing 2011). She assigned these remains to the somphocrinid Osteocrinus rectus rectus (Frizzell & Exline 1955). As in the present material the Triassic form has brachials articulated by both muscular and cryptosynarthrial facets. Food grooves (“Ventralfurchen”) may extend all along the ossicle, or they may be developed only near muscular articulations; alternatively, they may be dissolved into small pits or be absent altogether. In the present material, proper food grooves are either lacking or are dissolved into a series of small pits on the adoral side with its cryptosynarthrial facet (Fig. 10e, g, h). The axillary facets for the secundibrachials are directed outwards (aborally). The combination of the peculiar tall axils with the cups of R. pyramidalis is supported by matching facets between the different elements (cup and primibrachials); one cup has a first primibrachial attached, but displaced into the cavity. The cups are coarsely reticulate, with irregular vertical flanges or processes beneath the facets. The primibrachials are connected by a sloping cryptosynarthry. It may be particularly tall in the second primibrachials (Fig. 10g–i). Such ossicles match only with part of the corresponding facet of the mostly small first primibrachials. However, there are pairs that match well (Fig. 10q, e), even in cases where the second primibrachial is much larger (Fig. 10q, d). Some of the axils have a downward prolongation of the shaft beyond the facet (Fig. 10d). The position of the axial canal near the upper end of tall facets may indicate that articulation with the first primibrachial was restricted to the upper part (Fig. 10g); such an assumption is supported by a slight angle of the facet in the large ossicle in Fig. 10i, with the upper third serving for articulation.
Orthogonocrinus apertus Peck, 1943, Figs. 12–13.
1943 Orthogonocrinus apertus Peck, Peck, p. 464; pl. 76, figs. 2–8.
1955 Orthogonocrinus apertus Peck, Peck, p. 1022; pl. 106, figs. 7–9.
1961 Orthogonocrinus apertus Peck, Rasmussen, p. 385; pl. 56, figs. 4–5.
Cups (Fig. 12) The 240 cups correspond well to the material described by Peck (1943). The size of the cups ranges from less than 1 mm in height to about 3 mm. The smallest cup (Fig. 12g) has convex radials with only a faint aboral ridge; it is open at the lower end where the unbroken radials suggest that a ring of basals was originally present. Adult cups are more or less conical. Each radial has a vertical ridge extending from the facet to the aboral apex and beyond, as demonstrated by one of Peck’s specimens (1943, pl. 76, fig. 3). However, most of the cups are broken in the lower part exposing an aboral cavity that is of much greater height than the upper one. The cavities are separated by a horizontal partition (Fig. 12d). The partition has five peripheral and a central opening connecting the two chambers (Fig. 12e). The articular facets are angularly bent at the transverse ridge. The aboral ligament fossa is vertical and prominent on the side of the cup; it has a deep pit below the transverse ridge. The interarticular ligament fossae and the axial canal are situated on the upper, nearly horizontal or slightly inclined part of the facet. The interfacet processes are of unequal height and shape, even in a single individual. The muscle fossae extend to the lower part of the processes; they are separated from the radial cavity by a thin wall forming a V-shaped notch (Fig. 12j). In adoral view the interfacet processes project into the radial cavity, giving the cup a characteristic appearance (Fig. 12b).
Primibrachials (Fig. 13) There is a limited number of wingless primibrachials that may belong to this species. Some of the first primibrachials have corrugated muscle facets on the adoral side and are somewhat crooked (Fig. 13b, c, e). The aboral side lacks a bowl as developed in Poecilocrinus latealatus. The asymmetric profile may be the result of unequal interfacet processes to the base of which the muscles attached. Second primibrachials are aborally concave and smooth. The proximal facet is a cryptosynarthry matching the distal facet of the first primibrachial; the two distal facets are muscular and separated by a process.
Remarks Most of the cups of this distinct species are broken at the lower (distal) end, exposing the aboral cavity (hence the species name). However, the cups probably were closed at the bottom during life, as demonstrated by practically intact specimens (e.g. Peck 1943, pl. 76, fig. 3). In the present material, the specimen in Fig. 12f is largely intact. It seems possible that the smallest individuals originally were closed at the apex by a basal circlet that was later resorbed (Fig. 12g). In any case, the lower part of the cups of this species is prone to breakage as a result of the thin wall (Fig. 12e). Rasmussen (1961, p. 386) mentioned second primibrachials with broad lateral wings found together with cups of the present species. These remains are from the Santonian of Corneville (France) and are housed at the Geneva Museum. They are closely similar to those of Roveacrinus communis Douglas 1908 from the upper Cenomanian of Germany as figured by Rasmussen (1961, pl. 53, figs. 4 and 6); in this specimen the first primibrachials are covered by the second ones. The first primibrachials tentatively assigned to the present species lack distinct lateral wings or aboral bowls.
Roveacrinus peracutus (Peck 1943), Figs. 12h, ?13a, d.
1943 Drepanocrinus peracutus Peck, Peck, p. 463; pl. 76, figs. 9–22, 26, 28.
1943 isolated brachials of unknown affinities, Peck, p. 473; pl. 76, figs. 30, 33–35.
1955 Styracocrinus peracutus (Peck), Peck, p. 1022; pl. 106, figs. 10–12.
1955 unidentified primaxils, Peck, p. 1025; pl. 106, figs. 14–15.
1961 Styracocrinus peracutus (Peck), Rasmussen, p. 383; pl. 56, figs. 1–3.
1983 Styracocrinus peracutus (Peck), Pisera, p. 387; pl. 11, figs. 1–8; pl. 12, figs. 1–6.
The cup assigned to this species is small and broken below the facets. These are rather indistinct, only slightly angularly bent at the transverse ridge and prolonged downwards into flanges. The greatest diameter of the cup is just below the facets where the flanges are accompanied by lateral depressions. The interfacet processes are low. From below the aboral cavity has a partition with a distinct central opening but only small peripheral openings (see Fig. 17e for a specimen from Del Rio). First primibrachials with straight, non-corrugated adoral sides may belong to the present species (Fig. 13a, d), although the specimen in Fig. 13a may also belong to Poecilocrinus latealatus where much reduced bowls rarely occur (the specimen is also illustrated in Fig. 7l).

Cups of Roveacrinus peracutus Peck, Del Rio Formation (lower Cenomanian), Dottie Lynn Locality, Fort Worth, Texas. a Lateral view of intact cup with radial flange accompanied by narrow interradial flanges, M11718. b Small cup lacking interradial flanges, M11719. c Lateral-adoral view of low cup without interradial flanges, interradial sutures distinct; 11717. d Lateral view of broken cup with well-developed interfacet processes, M11722. e Aboral view of broken cup with largely intact partition at the centre and peripheral radial openings, M11721. f Adoral view of cup, M11720


Remarks This cup is distinguished from Orthogonocrinus apertus by its articular facets which narrow aborally into radial flanges. It corresponds to the material described by Peck (1943) as Drepanocrinus peracutus, a species later transferred by the same author (Peck 1955) to Styracocrinus. This genus is here considered a synonym of Roveacrinus (see below).
The Del Rio roveacrinids
The bulk of the Del Rio specimens illustrate well the difficulties that exist in assignment to any roveacrinid species, especially if juvenile and subadult individuals are also considered (Fig. 14). Peck (1943, fig. 23) noted the following genera and species from the Grayson Formation (all taxa established by him): Drepanocrinus peracutus, Orthogonocrinus apertus, Roveacrinus pyramidalis, R. euglypheus, R. signatus, R. pentagonus, R. spinalatus, R. multisinuatus, and R. spinosus. From this list, only Drepanocrinus (=Roveacrinus) peracutus, Roveacrinus pentagonus (=Roveacrinus alatus Douglas) and R. spinosus are represented in our material. According to Rasmussen (1961, p. 373) R. spinosus and spinalatus seem to be conspecific. The primibrachials and secundibrachials in the collection have a number of processes, spines or thorns and may be classified with R. spinosus, but others are of uncertain affinity. The brachials are discussed under R. spinosus.
Roveacrinus alatus Douglas, 1908, Figs. 14d, p, t; ? Fig. 15a, b.
1908 Roveacrinus alata Douglas, p. 358; pl. 17, figs. 1–2.
pars 1932 Drepanocrinus sessilis Jaekel, Sieverts, p. 599, fig. 6.
1943 Roveacrinus pentagonus Peck, Peck, p. 467; pl. 72, fig. 12; pl. 73, figs. 1–5, 7.
1955 Roveacrinus alatus Douglas, Peck, p. 1023; pl. 105, figs. 15–16.
1961 Roveacrinus alatus Douglas, Rasmussen, p. 364; pl. 54, figs. 1–4.
1988 Roveacrinus alatus Douglas, Schneider, p. 277, figs. 1–2.
2011 Roveacrinus alatus Douglas, Hess in Hess & Messing, p. 207; fig. 103,1a-b.
Cups The five cups available have large, thin vertical wings or flanges supporting the facets and extending beyond the aboral apex. The specimen in Fig. 14t has small secondary flanges connecting the upper part of the vertical flanges. In the specimen in Fig. 14p the flanges are not developed equally and are thickened in the upper part. The juvenile specimen (Fig. 14d) has main flanges giving off short branches. The radial facets are rather small in the specimen in Fig. 14t.
Remarks As demonstrated by Peck (1943, p. 467), Rasmussen (1961, p. 365) and Hess and Thiel (2015), R. alatus is a variable species, and our material is not an exception. The specimen in Fig. 14t resembles Peck’s (1943) specimens in pl. 72, fig. 12 and pl. 73, fig. 1; and the specimen in Fig. 14p resembles Peck’s pl. 73, fig. 7, although two flanges are rudimentary in our specimen.
Roveacrinus spinosus Peck, 1943, Figs. 14a–c, e–o, q–s; 15c–w.
1943 Roveacrinus spinosus Peck, p. 467; pl. 74, figs. 6, 7, 9.
1943 Roveacrinus spinalatus Peck, p. 467; pl. 74, figs. 8, 10.
1961 Roveacrinus spinosus Peck, Rasmussen, p. 373; pl. 54, figs. 5–6.
Cups The majority of cups in the Del Rio sample belongs to this species; 86 are larger than about 1 mm, and about 300 smaller (Fig. 14a). The radial articular facets are prolonged spine-like aborally, and the processes are connected to the bulbous aboral part of the cup by thin flanges. Such development is already present in juvenile cups. The processes are adorally concave and thickened on top, commonly with a pit (Fig. 14q, r). The wider ones are duckbill-shaped. In some cups the processes are very prominent (Fig. 14s), and they may not be developed equally around the cup (Fig. 14g, h, o, s). A broken cup shows a large central opening and smaller radial excavations between the two body chambers (Fig. 14c).
Brachials Most of the primibrachials and secundibrachials in the sample have processes or spines and appear to belong to Roveacrinus spinosus. Muscle facets are commonly more or less corrugated (Fig. 15c–d, n–t), and some secundibrachials have a synostosial facet (Fig. 15q, r). Facets for pinnules are present in a few brachials (Fig. 15m, u–w: arrows), an occurrence not otherwise present in the Del Rio and Duck Creek samples. Two relatively tall first primibrachials (Fig. 15a, b) may belong to R. alatus; their adoral side is moderately concave, with narrow lateral muscle fossae (Fig. 15b).
Remarks The holotypes of R. spinosus and R. spinalatus are from the same locality and horizon, and Peck (1943) regarded the forms to be closely related. Following Rasmussen (1961, p. 365), the forms are here regarded as conspecific, with R. spinosus the preferred name; this species is documented by more extensive material and has page priority.
Roveacrinus peracutus (Peck, 1943), Fig. 17.
1943 Drepanocrinus peracutus Peck, p. 463; pl. 76, figs. 9–22, 26, 28.
1955 Styracocrinus peracutus (Peck), Peck, p. 1022; pl. 106, figs. 10–12.
1961 Styracocrinus peracutus (Peck), Rasmussen, p. 383; pl. 56, figs. 1–3.
1971 Styracocrinus peracutus (Peck), Schmid (1971), p. 71, pl. 1, figs. 9–15.
Cups There are 70 cups in the sample. Variability is low; and adult specimens do not basically differ from small, subadult ones. The cups are slender and conical. The radial articular facets are prolonged into a narrowing ridge to the aboral apex. The sutures between the radials are distinct. The facets are outward-sloping and slightly angularly bent at the transverse ridge. They are separated by fairly strong, blunt interfacet processes of somewhat variable height, with distinct muscle fossae at their base (Fig. 17f). Broken cups have a partition at about a fourth of cup height from the top (Fig. 17e, f). The partition may be broken by a central opening (Fig. 17f), and smaller radial openings occur around the periphery. The radials are much thicker above the partition than below.
Remarks As pointed out by Rasmussen (1961, p. 383) the genus and a new subfamily (Styracocrininae) were established by Peck (1955) on erroneous assumptions. The slender, spike-like shape of the cups is a common character of species of Roveacrinus, such as R. communis (Douglas 1908) (see Peck 1955, pl. 105, figs. 1–14). In his key to the genera of Roveacrinida, Rasmussen (1961, p. 363) mentioned that Orthogonocrinus and Styracocrinus had “articular faces distinctly angulate about the articular ridge. The ventral part is almost horizontal; the large, dorsal ligament fossa is almost vertical.” In contrast, Styracocrinus was diagnosed by Rasmussen (1961, p. 383) as follows: “The articular faces of the radials are slightly angularly bent, strongly outward-sloping.” Styracocrinus is here regarded as a synonym of Roveacrinus.
Ontogenetic and phylogenetic trends
Previous discussions The oldest member of the family Roveacrinidae, Roveacrinus berthoui, was described by Ferré and Granier (2000) from the lower Hauterivian of Spain, based on a single cup identified from an oblique medial section. Due to its state of preservation even an approximate reconstruction of the cup’s morphology is conjectural, making this oldest recorded species unsuitable for phylogenetic studies. During Albian times the family radiated into widely different forms, as illustrated by Peck (1943) and herein. In his basic paper Peck (1943) discussed developmental trends but did not take ontogentic changes and brachials into consideration. He distinguished essentially three lines of development, starting from an unknown ancestor (Peck 1943, figs. 1–22). His scheme suffers from the problem that two of the basic species, Roveacrinus pyramidalis and Plotocrinus primitivus, probably are conspecific (Rasmussen 1961, p. 371). Species of Poecilocrinus certainly are the most spectacular roveacrinids. According to Peck the main character is a radial ridge bifurcating at mid-height and branching into a bowl-like depression under the arm facets. However, as shown above, branching of flanges on the aboral part of the cup is quite variable, making this character unreliable. More distinctive are horizontal wings fused into a dish below the articular facets. In one of his lineages Peck combined Poecilocrinus latealatus with Roveacrinus pyramidalis (as the basic form) and added Roveacrinus spinosus and R. alatus. In another lineage the basic form is Plotocrinus primitivus (=Roveacrinus pyramidalis). This lineage also includes Plotocrinus hemisphericus and Discocrinus catastomus, both with bowl-shaped cups, and also Poecilocrinus dispandus, the type species of the genus. A more logical grouping is discussed below.
Ontogenetic series in the present material Juvenile specimens that markedly differ from adults are present in most species. Differences between juveniles and adults are largest in Poecilocrinus latealatus and smallest in Roveacrinus peracutus. At times, the smallest cups of P. latealatus and Roveacrinus pyramidalis are difficult to distinguish. In contrast, brachials of the two species differ widely. The smallest cups of Poecilocrinus latealatus have a diameter of 0.6 mm. They are bowl shaped and roughly pentagonal, with a nearly flat upper rim and flat aboral apex (Figs. 4a, b, 5a, b). At the apex interradial knobs suggest the presence of basals in the process of resorption. Articular facets in the smallest cups rest on patches of denser stereom in an otherwise coarsely reticulate structure (Fig. 5a, b). During later stages processes grow downwards and outwards below the facets. The processes are first spine-like (Fig. 5c–h), they then become spoon- or duckbill-shaped (Fig. 5i–p) and subsequently wing-like (Fig. 5q–r) until they finally merge into a dish (Fig. 4k–t). The facets are supported by flanges that eventually grow to the aboral apex and beyond (Figs. 4, 6a). On the adoral side interfacet processes grow to variable heights (Fig. 6). The smallest cups of Roveacrinus pyramidalis are about 0.5 mm in diameter, and they differ markedly from larger cups. However, a continuous series is available so that assignment to a single species can be made with confidence. Small cups with a diameter of 0.5–1 mm are pentagonal. The aboral apex is nearly flat and, when broken, has spherical granules in a seemingly developing lower chamber (Fig. 9a–d). Spine-like radial processes are either lacking (Fig. 9a) or are present below one or the other of the facets (Fig. 9b–d). Small cups of Roveacrinus pyramidalis and Poecilocrinus latealatus are quite similar (e.g. Figs. 5c, 9f). At the earliest stages spine-like processes are formed below some of the facets. At later stages thin radial processes grow outwards in irregular fashion, but they do not extend beyond the aboral apex. Interfacet processes may also be variable. In contrast to P. latealatus, the aboral apex is more or less conical and the stereom is coarsely reticulate. The largest cups reach only about half the size of P. latealatus. The smallest cup of Orthogonocrinus apertus (Fig. 12g) has bulging radials; it is 1 mm in height at a diameter half that size. Such a ratio is also present in large cups (Fig. 12a). Larger cups are more conical and have flanges below the articular facets. The aboral part of the facet with the deep ligament pit is vertical, and the adoral part with the central canal is weakly sloped to nearly horizontal; such geometry is maintained throughout growth. The smallest cup of Roveacrinus spinosus has a diameter and height of about 0.5 mm (Fig. 14a). The vertical flanges extending from the inclined articular facets are already well developed. During growth this basic morphology is largely maintained, although the flanges become thickened and are prolonged in irregular fashion below the facets. The smallest cup of Roveacrinus peracutus has a diameter of 0.5 mm at a height of 0.75 mm, a ratio more or less maintained during growth (Fig. 17a, b). The conical shape with a major flange that extends from the articular facet to the aboral apex is also largely unchanged, as are the outward-sloping articular facets.
The Late Albian–Early Cenomanian radiation produced several lines of cup morphotypes (Fig. 20). (1) A first type has conical cups with modest vertical flanges, wings or spines are absent. This type has essentially isometric growth. The lineage includes Roveacrinus peracutus and Orthogonocrinus apertus. (2) A second type has wide and low, bowl-shaped cups lacking wings or spines; articular facets are large and nearly vertical. It includes Discocrinus integer, D. catastomus and Plotocrinus hemisphericus with a series of small vertical flanges. (3) A third type has cups that are greatly modified during growth, resulting in cups with spectacular dish-like horizontal wings below the articular facets. Characteristic species in this lineage are Poecilocrinus latealatus and P. dispandus. (4) In a fourth type the cups have prominent vertical flanges that may be thickened below the outward-sloping articular facets. The shape tends to be asymmetric and variable. Included forms are Roveacrinus pyramidalis, R. peracutus, R. alatus and R. spinosus. In the first and second groups the cup hardly changes in shape during ontogeny. The third and fourth groups may have similar juvenile cups. However, they are greatly modified during growth, and arm morphology of adults is also different.

Diversification of roveacrinids during lifetime of the order. Selected examples are from Jaekel (1892), Jagt (1999), Hess (2002), Hess and Gale (2010), Hess and Messing (2011), and the present paper; figures of the specimens not to scale. IBr1 first primibrachial, IBr2 second primibrachial, IIBr secundibrachial, prox proximal facet, di distal facet
Phylogeny In addition to the small size and lack of a column strongly sloping cryptosynarthries between the primibrachials and the first and second secundibrachials are the prominent diagnostic characters of all Roveacrinida, from the Triassic to the Cretaceous (Hess in Hess and Messing 2011). It is astonishing that this synapomorphy was successfully maintained throughout the history of the order, including forms with widely different arms such as Poecilocrinus latealatus, Roveacrinus pyramidalis and Saccocoma tenella, to name but a few (Fig. 20). Phylogenetic trends must be recognised within the frame of this synapomorphy, namely in cups and their ontogeny. For example, the smallest cups of the cyrtocrinid Eugeniacrinites cariophilites (see Hess 2014) are surprisingly similar to equally sized cups of Poecilocrinus latealatus. Basals are lacking in the former and are vestigial in the latter; further growth mainly affects the height of the interfacet processes in both species. The facet to the column is indistinct in the smallest E. cariophilites. The distinctive morphology of the two species is only reached subsequently, in P. latealatus with its floating lifestyle, and in the fixed E. cariophilites with its protection against predators. Articulation between the primibrachials of E. cariophilites is straight synostosial, as in other cyrtocrinids, making it unsuitable for the study of possible relationships between Cyrtocrinida and Roveacrinida.
The oldest well-defined representative of the Roveacrinidae is Discocrinus integer Hess (Hess and Gale 2010) from the lower Albian (Fig. 20). It is characterised by a low hemispherical cup with interfacet extensions of similar height but devoid of wings, flanges or spines. The articular facets for the arms are large and nearly vertical. The cup has two cavities, the upper one separated from the lower one by a thin plate comprising the fused basals. The surface of the cup is finely rugose, resembling that of Saccocoma tenella (Goldfuss 1831) from the Tithonian. This is a highly specialised form (Hess and Etter 2011) and, thus, not likely to be an ancestor to Discocrinus. However, other Saccocominae have characters also present in various Albian species (see Hess 2002). Such characters are scalloped or corrugated muscle fossae (Crassicoma schattenbergi Sieverts-Doreck & Hess in Hess 2002) and nearly vertical, large articular facets on low cups (Crassicoma feifeli Sieverts-Doreck & Hess in Hess 2002) (see Fig. 20). The early Late Jurassic radiation produced a number of quite different species, six of Crassicoma and one or two of Saccocoma (see Brodacki 2006). Because Crassicoma schattenbergi has thick-walled cups and lacks wings or other floating aids it were considered by Hess (2002; p. 27, fig. 14) to have been essentially benthic. On the other hand, the presence of articulations with strongly developed muscle fossae and the loss of the column indicate that it was also an active swimmer at times. The cups of the stratigraphically younger Crassicoma feifeli, C. praeschattenbergi and C. subornata are much smaller than that of C. schattenbergi, hardly exceeding 1 mm in diameter. Such miniaturisation suggests an increased facility for permanent floating. In the larger Saccocoma species the skeleton is lightened and wings, flanges or spines on cups and arms indicate a permanently pelagic lifestyle. This was demonstrated by Hess and Etter (2011) for S. tenella, by far the largest member of the genus. Crassicoma and Saccocoma resulting from the Late Jurassic radiation were followed during the Albian radiation by similar genera to the Roveacrinidae. Poecilocrinus may be derived from Saccocoma, while Crassicoma may have been ancestral to Discocrinus and Plotocrinus. The ancestor of species of Roveacrinus and Orthogonocrinus with their conical cups is unclear, although one is reminded of the Triassic Somphocrinidae with their spine-like aboral processes (also called centrodorsals in the literature; see Peck 1948, and Kristan-Tollmann 1970). While this may be a convergence resulting from the floating lifestyle the similarity between the arms of R. pyramidalis and the Triassic Osteocrinus discussed below is utterly astonishing. In contrast, the closely similar juvenile cups of Roveacrinus pyramidalis and Poecilocrinus latealatus suggest a common origin despite the widely different arm structure.
Functional morphology and lifestyle of the Duck Creek and Del Rio roveacrinids
Role of the double cavity in cups Roveacrinidae basically have a cup with double cavity. It comprises radials that have overgrown small, discrete basals, thereby forming an upper (oral) cavity above the basals and a secondary, mostly closed and globular cavity below the basals (Hess in Hess and Messing 2011, p. 207). According to Rasmussen (1961, p. 362) the presence of two cavities is common to all Roveacrinidae, except perhaps Discocrinus. The cavities are separated by a thin wall. In most specimens this wall lacks distinct sutures, but it in some it comprised small basals. The smallest specimens of Poecilocrinus latealatus in the present material lack a separating wall, but it is present at later stages (Fig. 6). Two chambers connected by openings are well displayed in broken specimens of Orthogonocrinus apertus (Fig. 12d, e) and Roveacrinus peracutus (Fig. 17e). Schneider (1989, 1995) proposed that the lower, aboral cavity was able to store gas or low-density fluids and functioned as a hydrostatic organ. He assumed this not only for Roveacrinus geinitzi Schneider 1989 and Orthogonocrinus janeti (Valette 1917), but for all roveacrinids with two body chambers. Accordingly, the hydrostatic organ supported vertical movements without much muscle activity. Jagt (2005) suggested that voids in the cup of the late Maastrichtian Birgenelocrinus degraafi Jagt 1999 were probably used for gaseous exchange. The idea of a hydrostatic organ to facilitate movement in the water column seems conjectural. Accumulation of gas in the lower chamber might even turn the cup over so that the aboral apex is up and the arms are hanging down. Morphology of Poecilocrinus species does not support such a position. After Peck (1943), the position of roveacrinids in the water column was not discussed by Schneider (1989, 1995) nor by any other author (see above). For roveacrinids with a conical cup, such as Orthogonocrinus apertus or Roveacrinus peracutus, the notion of an adoral-down position might appear attractive at first sight. Upward movement would be facilitated by the streamlined shape of the cup and possible lightening from gas in the aboral chamber. The animal would feed while sinking in upstream fashion. However, the presence of brachials with lateral wings in Roveacrinus communis favours a mouth-up lifestyle (see next section). The cups of Orthogonocrinus apertus are characterised by particularly large, vertical aboral ligament pits. Muscle contraction would have loaded the aboral collagen spring; and upon muscle relaxation the arms were rapidly pushed downwards to sift the plankton from the water downstream and convey it to the oral cavity. Such food collection is linked to a mouth-up position. Roveacrinus pyramidalis has a rather small and indistinct aboral cavity, and the peculiar brachials also suggest a mouth-up position during floating. If development of an aboral cavity during ontogeny was unconnected with movement in the water column it must have served some other purpose, perhaps the temporary storage of nutrients, or even room for the reproductive system? The rather wide openings between the cavities in species such as O. apertus, R. acutus and R. alatus favour of such an assumption.
Cups: horizontal wings or vertical ridges and flanges The Duck Creek material contains three types of roveacrinid cups that are distinct at the adult stage. Poecilocrinus latealatus has cups with horizontal wings below the articular facets; the wings were supported by vertical flanges leading to the aboral apex. In combination with the winged proximal brachials this form was pelagic. The cup of Roveacrinus pyramidalis has irregular vertical flanges or processes below the facets. In combination with rod-shaped, very tall second primibrachials a benthic lifestyle may seem possible, as discussed in the next section. The cups of Orthogonocrinus apertus and Roveacrinus peracutus are conical and have moderate vertical flanges. Morphology of the sharply curved articular facet of Orthogonocrinus suggests that rapid vertical arm movements were possible by alternating muscle contraction and relaxation as explained above. The conical shape would have facilitated vertical movements of the cup. Altogether, the brachials are poorly known, although Rasmussen (1961, pl. 56, fig. 3) figured a cup of Roveacrinus peracutus with tall, keeled second primibrachials and moderately high first secundibrachials still attached. Specimens of Roveacrinus communis, described by Sieverts (1932) as Drepanocrinus sessilis, have brachials with lateral flanges and adoral food grooves (see also Rasmussen 1961, pl. 53, figs. 4–6) that suggest a pelagic, mouth-up lifestyle. The Del Rio material includes Roveacrinus alatus and R. spinosus whose cups are provided with lateral flanges and spines suitable for floating. In combination with spiny brachials a pelagic lifestyle may be assumed for these forms.
Morphology of brachials The Duck Creek material includes numerous brachials of two basic types, assignable to Poecilocrinus latealatus and Roveacrinus pyramidalis. Poecilocrinus has first primibrachials with a bowl of variable size on the free, aboral surface. The second primibrachials have a large dish-like and aborally concave wing. Such a wing is also developed in the following secundibrachials whose adoral side contains the body of the ossicle with the articular facets. In more distal secundibrachials the wings are narrower but taller, and they eventually become spine-like (Fig. 18). The brachials have a distinct food groove, and muscle fossae commonly are corrugated. Articulations are mostly muscular, but brachials, both with and without wings, may also have synostosial articulations. The partly articulated specimen in Fig. 6a and intact specimens of the similar Poecilocrinus dispandus Peck figured by Scott et al. (1977) give the impression of a compact, inflexible crown in the lower part. In contrast, the middle, but especially the distal parts of the arms were movable and suitable for swimming. Dish-like lateral wings that are concave aborally are also developed in proximal brachials of the Jurassic Saccocoma tenella (Hess and Etter 2011; see also Fig. 20 here). However, in S. tenella more distal brachials carry paired vertical processes flanking the food grooves. The arms are heavily branched. Such sophisticated morphology, not seen in other Rovacrinida, led Hess and Etter (2011) to propose a pulsating funnel for feeding, in upstream fashion and mouth-up position. The similar bowl-shaped cups and proximal arms with aborally concave wings suggest that Poecilocrinus also lived mouth-up and was able to swim, using the mobile part of the arms. The wings served to slow the sinking process during which the animals could feed in the downstream mode, common to crinoids. In contrast to Saccocoma tenella, the cups of Poecilocrinus latealatus are highly variable, especially during development (Fig. 4), which is quite unusual for a pelagic animal. It cannot be excluded that the youngest stages of Poecilocrinus were benthic. Saccocoma tenella with its essentially unchanged cup morphology during growth certainly was pelagic.
Brachials of Roveacrinus pyramidalis are very different. As reconstructed (Figs. 10r+q+d; 19), the first primibrachials are directed outwards and upwards at an angle of 20°–30° from the vertical axis. Size and height of the second primibrachials are out of proportion to cups and first primibrachials. The primibrachials are linked by an immovable cryptosynarthrial articulation, seemingly cumbersome morphology for a mobile, pelagic animal. Up to the first branching, arm mobility was limited to the facet between cup and first primibrachial; and the lower part of the crown must have resembled a tall cage, although with partly movable bars (Fig. 19). The downward prolongation of certain second primibrachials (Fig. 10d) may have helped to anchor the animal in the sediment. The primibrachials and secundibrachials (Fig. 11) lack proper food grooves; these are partly replaced by indistinct impressions on the adoral side of the ossicles. Except in the proximal part of the first primibrachials (Fig. 10k, l, q), the stereom of the brachials is dense and the surface smooth. Such morphology is not in favour of a floating lifestyle. Secundibrachials are reinforced aborally by a ridge or keel, as if to avoid easy breaking (Fig. 11e, j, k, n–o). If it were not for the similar brachials of the Triassic Osteocrinus rectus (Frizzell and Ex1ine 1956), as figured by Kristan-Tollmann (1970, fig. 5; see also Hess, in Hess and Messing 2011, fig. 109, 3b), and of other species of Osteocrinus (Wang et al. 2007, 2008), a benthic mode of life might be an acceptable option. The Triassic forms and the Cretaceous Roveacrinus pyramidalis share tall, rod-shaped brachials connected by muscular and ligamentary articulations; and even distal secundibrachials are similar (Kristan-Tollmann in Kristan-Tollmann and Krystyn 1975, pl. 6, figs. 2, 3). The Triassic species also lack proper food grooves so that food presumably was acquired as in R. pyramidalis. A pelagic lifestyle, as already proposed by Kristan-Tollmann (1970), is substantiated by the mass occurrence of species of Osteocrinus in anoxic black shales of the Xiaowa Formation (lower Carnian) of southern China (Wang et al. 2007; Wang et al. 2008). The Chinese Osteocrinus may belong to two different species, one with a short, funnel-shaped aboral process on the cup (referred by Wang et al. 2007 to O. cf. virgatus Kristan-Tollmann 1970), and one with a long, spine-like aboral process (referred to O. spinosus Kristan-Tollmann 1970); these finds await proper description. According to Hans Hagdorn (pers. comm., 2015) taphonomic data indicate mass death of the animals which sank to the bottom in various states of disarticulation. Longer (=taller) brachials commonly are current-oriented (Wang et al. 2008, fig. 12A). An arm fragment in initial state of disarticulation is composed of quite a number of tall brachials so that the arms were unusually long and dominant in relation to the cups (Wang et al. 2008, fig. 12B). Wang et al. (2008, text-fig. 12C) figured a low cone-shaped cup associated with oversized-looking primibrachials, an almost irrational morphology astonishingly similar to that of R. pyramidalis (Fig. 10r+q+d). Aboral processes (“centrodorsals”) are coarse-meshed and may be extremely long (Wang et al. 2007, pl. 2, fig. 3), suggesting a stabilising function during floating. The stereom of the high brachials is wide-meshed near the facets; and the shafts of the larger brachials are thin-walled and apparently hollow tubes. They were easily broken after burial and compaction in the sediment. Hollow brachials are most unusual in crinoids and seem to defy the architectural constraints of the group. Possibly the stereom was wide-meshed or even mostly absent within the brachials for easier floating. High brachials are not hollow in the specimens from the Eastern Alps described by Kristan-Tollmann (1970), although this could have been the result of lithification and recrystallisation, not present in the Chinese material which is preserved in bituminous shales (Wang et al. 2008).
Lifestyle: benthic versus nektonic or pelagic The diminutive stalkless crinoids of the Order Roveacrinida have adaptations consistent with a pelagic lifestyle, such as a delicate skeleton, and cups ornamented by spines, flanges or wings to increase frictional drag. Nevertheless, their lifestyle has been a matter of controversy. As passive suspension feeders they may collect food by exposing their arms into the current (benthic lifestyle), or by moving actively through the water (nektonic lifestyle). Drifting passively with the plankton may occur early in development, but capture of food essentially depends on movement of particles past the arms.
All Roveacrinida are linked by the synapomorphy of strongly sloping cryptosynarthries in the proximal part of the arms (Hess in Hess and Messing 2011). The most primitive member is the upper Triassic Axicrinus alexandri Kristan-Tollmann (1977) with distinct basals and compact radials and primibrachials devoid of flanges or spines. It has been considered by Kristan-Tollmann (1971) to have been pelagic (“Schwebcrinoiden”); Hess (2010) suggested that it was essentially benthic, but may at times have been an active swimmer. The other Triassic Roveacrinida, grouped in the Somphocrinidae, have cups with spine-like aboral processes and long arms; lateral wings and flanges are absent. They were generally considered to have been pelagic (Peck 1948; Kristan-Tollmann 1977), not the least because of their mass occurrence in the Tethys realm (Kristan-Tollmann 1988). This assumption is confirmed by the occurrence in Ladinian black shales of China, discussed above.
The upper Jurassic Saccocoma tenella has a lightened skeleton, dish-like wings on proximal brachials and oral processes on distal brachials. In contrast to Milsom (1994) and Manni et al. (1997) Hess and Etter (2011) proposed that S. tenella was nektonic, feeding actively by a pulsating funnel. The presence of wings and oral processes in even the smallest specimens (cup diameter 2 mm) is also in favour of a permanently pelagic lifestyle. This is substantiated by ecologic and taphonomic evidence. Other Saccocoma species, such as S. quenstedti Sieverts-Doreck & Hess (Hess 2002) and S. longipinna Hess (2002), lack oral processes on the distal brachials. Thus, they could not feed by a pulsating funnel but are considered pelagic by virtue of their morphology.
The Cretaceous roveacrinids have generally been described as pelagic, although Scott et al. (1977) and Milsom (1999) left this question open. In a short note Milsom (1999) suggested that roveacrinids exploited two different lifestyles; (1) benthic with the ability to swim, and (2) nektonic. Unfortunately, these assumptions were not substantiated by corresponding evidence, nor were ontogenetic stages considered. Perhaps induced by her paper on Saccocoma, Milsom (1994) characterised the benthic roveacrinids by the presence of flanges and spines on calyx (=cup) and proximal brachials, and by limited arm mobility. Nektonic roveacrinids are characterised by an enlarged dorsal (aboral) cavity, extreme arm mobility, and are virtually devoid of ornamentation, without evidence for skeletal lightening. As shown by the present material some characters are present in Poecilocrinus, but others are not. Adult individuals of Poecilocrinus have characters indicating a pelagic mode of life. Forms with rod-shaped brachials, such as the pelagic Osteocrinus and Roveacrinus pyramidalis, were not discussed by Milsom. A pelagic mode of life of adult roveacrinids with downstream feeding during sinking is accepted.
Lifestyle: open questions The mode of life of two of the species discussed here, Poecilocrinus latealatus and Roveacrinus pyramidalis, now appears resolved. However, some open questions remain. Juvenile cups of the two forms are similar. In Poecilocrinus latealatus lateral dish-like wings on the cups are developed at a later stage, making a benthic lifestyle at the postlarval stage seem possible. On the other hand, appearance of lateral spines on juvenile cups of Roveacrinus pyramidalis (Fig. 7b–d) suggests an early step to floating. Variability of interfacet processes is present in P. latealatus, R. pyramidalis and Orthogonocrinus apertus; and the relevance of this is unclear. In O. apertus muscle fossae are restricted to the lower part of the processes and are quite uniform, regardless of the height of the processes (Fig. 12). Unfortunately, secundibrachials have not been identified in this species to confirm the supposed pelagic lifestyle. In contrast, secundibrachials with lateral wings are known from species of Roveacrinus with conical, high cups, indicating a floating mode of life. This is also assumed for Roveacrinus spinosus with processes on cups and second primibrachials as well as spines on secundibrachials. Incidentally, this species also has pinnules on some of the brachials. In R. spinosus lateral processes may occur on one side only in axillary secundibrachials (Fig. 15i, j) and in cups (Fig. 14). This is also true of Roveacrinus alatus (see Peck 1943, pl. 73, fig. 4, 7). Weird-looking cups are not only present in R. spinosus and R. alatus (Peck 1943; Schneider 1988) but also in the Late Campanian Veugelersia (see Jagt 1999), which resemble ragged ghosts. Is there a possibility that unilateral processes or spines on cups and brachials were deterrents to predators, and if so, which ones? Pronounced asymmetry is unexpected in pelagic crinoids, not the least because symmetric cups of Orthogonocrinus apertus (Fig. 12) and Roveacrinus peracutus (Fig. 17) occur alongside asymmetric cups of Roveacrinus pyramidalis (Fig. 9) and Roveacrinus spinosus (Fig. 14). A convincing explanation for such differences has not been offered yet.
A real puzzle is how Roveacrinus pyramidalis and the Triassic Osteocrinus were able to feed in the absence of food grooves on the rod-shaped brachials. The usual down-current capture of phyto- or zooplankton by tube feet and transport along the food grooves to the mouth is not possible. A Recent analogue is not available so that feeding remains a matter of conjecture. If the arms were not used for the collection of food they merely served to transport the cup to places where perhaps tiny food particles were captured and transported by ciliated cells to the mouth, or dissolved nutrients were absorbed directly from the water. But why were the arms so prominent and, at least in the Chinese Osteocrinus, so long? That such peculiar arm morphology was “invented” at least twice during earth history, with a gap of 125 myr, speaks of its success.
Conclusions
The Cretaceous Roveacrinidae includes the largest diversity of microcrinoids known in Earth history (Fig. 20), the first well-identified species being Discocrinus integer from the lower Albian. A major radiation took place in the mid-Cretaceous in Texas, producing the forms described in the present paper. Some species may be derived from Saccocomidae of the early Late Jurassic radiation. The distinctive genus Poecilocrinus can be derived from Saccocoma, whereas Crassicoma was probably ancestor to Discocrinus and Plotocrinus. Ancestry of Roveacrinus and Orthogonocrinus with conical cups is unclear. These resemble the Triassic Somphocrinidae, but this may be due to convergent evolution resulting from similar lifestyles. High variability of cups and absence of brachials that can be assigned to any given species hamper a phylogenetic approach to the classification of some of the species. Poecilocrinus and Roveacrinus pyramidalis have similar cups as juveniles, but their adult cups and especially their brachials are markedly different. This might be explained by different lifestyles, pelagic in P. latealatus and benthic in R. pyramidalis. However, rod-shaped brachials, such as those of R. pyramidalis also occur in Triassic Somphocrinidae of the genus Osteocrinus (upper Ladinian-lower Carnian). Based on their mass occurrence and wide distribution in different lithofacies of the Tethys Ocean (“Osteocrinus facies”) Kristan-Tollmann (1970) assumed a pelagic lifestyle, comparable to the saccocomids of the Upper Jurassic (“Lombardia facies”, Hess 2002). Such a mode of life is confirmed by the occurrence of Osteocrinus in Upper Triassic black shales of southern China (Wang et al. 2007; Wang et al. 2008). Roveacrinus pyramidalis and Poecilocrinus latealatus developed in parallel from similar juvenile cups into widely different adults. The latter presumably fed on coccolithophores and foraminifera that became the dominant plankton at mid-Cretaceous times. The feeding mode of R. pyramidalis is an enigma, as is that of the similar Osteocrinus. Other unanswered questions relate to the asymmetry of the interfacet processes in cups of R. pyramidalis and Orthogonocrinus apertus, and the variable lateral flanges and processes in the cups of Roveacrinus alatus and R. spinosus. Asymmetry and variability of cups is not present in the truly pelagic Saccocoma tenella which probably lived on coccolithophores, as demonstrated by their remains as a component of the Solnhofen limestone.
References
Brodacki, M. (2006). Functional anatomy and mode of life of the latest Jurassic crinoid Saccocoma. Acta Palaeontologica Polonica, 51, 261–270.
Destombes, P. (1985). Roveacrinidae nouveaux de l’Albien du Bassin de Paris. Bulletin trimestriel de la Société géologique de Normandie et Amis du Muséum du Havre, 71, 9–16.
Destombes, P., & Breton, G. (2001). Platelicrinus campaniensis nov. gen. nov. sp. Roveacrinidae (Echinodermata, Crinoidea) du Campanien des Charentes, France. Bulletin trimestriel de la Société géologique de Normandie et Amis du Muséum du Havre, 87, 37–40.
Douglas, J. A. (1908). A note on same new Chalk crinoids. Geological Magazine, ser., 5(5), 357–359.
Farinacci, A., & Manni, R. (2003). Roveacrinids from the Northern Arabian Plate in SE Turkey. Turkish Journal of Earth Sciences, 12, 209–214.
Ferré, B., & Granier, B. (2000). Roveacrinus berthoui, nov. sp., early Hauterivian representative of Roveacrinidae (Roveacrinida, Crinoidea) of Busot (Alicante, Spain). Geologica Carpathica, 51, 101–107.
Ferré, B., & Granier, B. (2001). Albian roveacrinids from the southern Congo Basin off Angola. Journal of South American Earth Sciences, 14, 219–235.
Frizzell, D. L. & Ex1ine, H. (1956). Monograph of fossil holothurian sclerites. Bulletin of the Missouri University School of Mines and Metallurgy, Technical Series, 89 (1955), 204 pp.
Gorzelak, P., Salamon, M. A. & Ferré, B. (2011). Pelagic crinoids (Roveacrinida, Crinoidea) discovered in the Neogene of Poland. Naturwissenschaften, 98, 903–908.
Hess, H. (2002). Remains of Saccocomids (Crinoidea: Echinodermata) from the Upper Jurassic of southern Germany. Stuttgarter Beiträge zur Naturkunde, Ser. B (Geologie und Paläontologie), 329, 57 pp.
Hess, H. (2010). Paleoecology of pelagic crinoids. Treatise Online, 16, 33 pp. Treatise on Invertebrate Paleontology, Part T, Revised, Volume 1, Chapter 19. The University of Kansas, Paleontological Institute.
Hess, H. (2014). The crinoid Eugeniacrinites cariophilites from the Late Jurassic of southern Germany: babies, cripples and enigmatic wing plates. Swiss Journal of Palaeontology, 133, 121–140.
Hess, H., & Etter, W. (2011). Life and death of Saccocoma tenella (Goldfuss). Swiss Journal of Geosciences, 104(Supplement 1), S99–S106.
Hess, H., & Gale, A. S. (2010). Crinoids from the Shenley Limestone (Albian) of Leighton Buzzard (Bedfordshire). Journal of Systematic Palaeontology, 8, 427–447.
Hess, H. & Messing, C. G. (2011). Treatise on Invertebrate Paleontology, Part T, Echinodermata 2, Revised, Crinoidea Volume 3 (W. I. Ausich Ed.), 261 pp. The University of Kansas Paleontological Institute: Lawrence, Kansas.
Hess, H. & Thiel, H.-V. (2015). Schwebcrinoiden aus dem untersten Cenomanium vom Kassenberg in Mülheim-Broich. Fossilien. (in press).
Jaekel, O. (1892). Über Plicatocriniden, Hyocrinus und Saccocoma. Zeitschrift der deutschen geologischen Gesellschaft, 44, 619–696.
Jagt, J. W. M. (1999). Late Cretaceous - Early Palaeogene echinoderms and the K/T boundary in the southeastern Netherlands and northeastern Belgium. Pt. 1, Introduction and Stratigraphy; Pt. 2, Crinoids. Scripta Geologica, 116, 255 pp.
Jagt, J. W. M. (2005). The youngest pelagic crinoids (latest Maastrichtian, the Netherlands). Bulletin of the Geological Society of Denmark, 52, 133–139.
Kristan-Tollmann, E. (1970). Die Osteocrinusfazies, ein Leithorizont von Schwebcrinoiden im Oberladin-Unterkarn der Tethys. Erdöl und Kohle, Erdgas, Petrochemie, 23, 781–789.
Kristan-Tollmann, E. (1977). Zur Gattungsunterscheidung und Rekonstruktion der triadischen Schwebcrinoiden. Paläontogische Zeitschrift, 51, 185–198.
Kristan-Tollmann, E. (1988). Unexpected communities among the crinoids within the Triassic Tethys and Pantalassa. In R. D. Burke, P. V. Mladenov, P. Lambert, & R. D. Parsley (Eds.), Echinoderm Biology (pp. 133–142). Rotterdam: Balkema.
Kristan-Tollmann, E., & Krystyn, L. (1975). Die Mikrofauna der ladinisch-karnischen Hallstätter Kalke von Saklibeli (Taurus-Gebirge, Türkei) I. Sitzungsberichte der Österreichischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse Abt. I, 184, 259–340.
Manni, R., Nicosia, U., & Tagliacozzo, L. (1997). Saccocoma, a normal benthonic stemlesss crinoid: an opportunistic reply within mud-dominated facies. Palaeopelagos, 7, 121–132.
Milsom, C. V. (1994). Saccocoma: a benthic crinoid from the Jurassic Solnhofen Limestone, Germany. Palaeontology, 37, 121–129.
Milsom, C. V. (1999). Rovings of the roveacrinids. In M. D. Candia Carnevali, F. Bonasoro (Eds.), Echinoderm Research 1998, 339. Balkema: Rotterdam.
Nekvasilová, O., & Prokop, R. (1963). Roveacrinidae (Crinoidea) from the Upper Cretaceous of Bohemia. Věstnik Ŭstredniho Ustavu Geologickěko, 38, 49–52.
Peck, R. E. (1943). Lower Cretaceous crinoids from Texas. Journal of Paleontology, 17, 451–475.
Peck, R. E. (1948). A Triassic crinoid from Mexico. Journal of Paleontology, 22, 81–84.
Peck, R. E. (1955). Cretaceous microcrinoids from England. Journal of Paleontology, 29, 1019–1029.
Pisera, A. (1983). Comatulid and roveacrinid crinoids from the Cretaceous of central Poland. Acta Palaeontologica Polonica, 28, 385–391.
Rasmussen, H. Wienberg. (1971). Cretaceous Grinoidea (Comatulida and Roveacrinida) from England and France. Bulletin of the Geological Society of Denmark, 20, 285–294.
Rasmussen, H. W. (1978). Articulata. In R. C. Moore & C. Teichert, Eds., Treatise on Invertebrate Paleontology. Pt. T, Echinodermata 2, vol. 3, T813-T928. Geological Society of America, Boulder; and University of Kansas Press, Lawrence.
Rasmussen, H. Wienberg (1961). A Monograph on the Cretaceous Crinoidea. Kongelige Danske Videnskaberne Selskab, Biologiske Skrifter 12 (1), 428 pp.
Salamon, M. A., Gorzelak, P., Ferré, B., & Lach, R. (2010). Roveacrinids (Crinoidea, Echinodermata) survived the Cretaceous-Paleogene (K-Pg) extinction event. Geology, 38, 883–885.
Schmid, F. (1971). Mesofaunen aus dem Alb von Hannover. Bericht der Naturhistorischen Gesellschaft Hannover, Beiheft, 7, 69–77.
Schneider, H. L. (1987). Zur Kelchmorphologie und Systematik der Roveacrinidae Peck, 1943 (Crinoidea, Oberkreide). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 175, 181–206.
Schneider, H. L. (1988). Roveacrinus alatus Douglas 1908, ein Crinoid aus der Oberkreide von Nordrhein-Westfalen. Der Aufschluss, 39, 277–281. Heidelberg.
Schneider, H. L. (1989). Zur Morphologie und Ontogenese von Roveacrinus geinitzi n. sp. (Crinoidea, Oberkreide). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 178, 167–181.
Schneider, H. L. (1995). Crinoidea (Roveacrinida) aus dem Unter-Turon in Wüllen (Münsterländer Kreidebecken/Nordrhein-Westfalen). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 198, 35–46.
Scott, R. W., Root, S. A., Tenery, J. H., & Nestell, M. (1977). Morphology of the Cretaceous microcrinoid Poecilocrinus (Roveacrinidae). Journal of Paleontology, 51, 343–349.
Sieverts, H. (1932). Über die Crinoidengattung Drepanocrinus Jaekel. Jahrbuch der Preussischen Geologischen Landesanstalt, 53, 559–610.
Valette, A. (1917). Note sur les Crinoïdes de la craie blanche. Bulletin de la Société des sciences historiques et naturelles de l’Yonne, 1916, 79–178.
Wang, X., Bachmann, G. H., Hagdorn, H., Sander, P. M., Cuny, G., Chen, X., et al. (2008). The Late Triassic black shales of the Guanling Area (Guizhou Province, Southwestern China)—A Unique Marine Reptile and Pelagic Crinoid Fossil Lagerstätte. Palaeontology, 51, 27–61.
Wang, Ch., Wang, X., Hagdorn, H., Chen, X.-H., & Chen, L. (2007). The first discovery of Triassic roveacrinids in China and its significance. Acta Palaeontologica Sinica, 46, 334–339. (In Chinese with English Abstract).
Acknowledgments
My warm thanks to Andy Gale for keeping my old brain on the go with new material that he tirelessly collects and processes. The thorough reviews and constructive comments by John W. M. Jagt and an anonymous reviewer are gratefully acknowledged. I am very grateful to Evi Bieler, Zentrum für Mikroskopie at the University of Basel for her patience with the SEM micrographs. Hans-Volker Thiel provided roveacrinid material from the lower Cenomanian of Germany for comparison; this material will be described in a separate paper. I like to thank Walter Etter for the line drawings. Hans Hagdorn provided data on Triassic somphocrinids and reviewed an earlier version of the typescript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hess, H. Roveacrinids (Crinoidea) from the mid-Cretaceous of Texas: ontogeny, phylogeny, functional morphology and lifestyle. Swiss J Palaeontol 134, 77–107 (2015). https://doi.org/10.1007/s13358-015-0076-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-015-0076-z